
Minigene as a Novel Regulatory Element in Toxin-Antitoxin Systems


Abstract
1. Introduction
2. Results
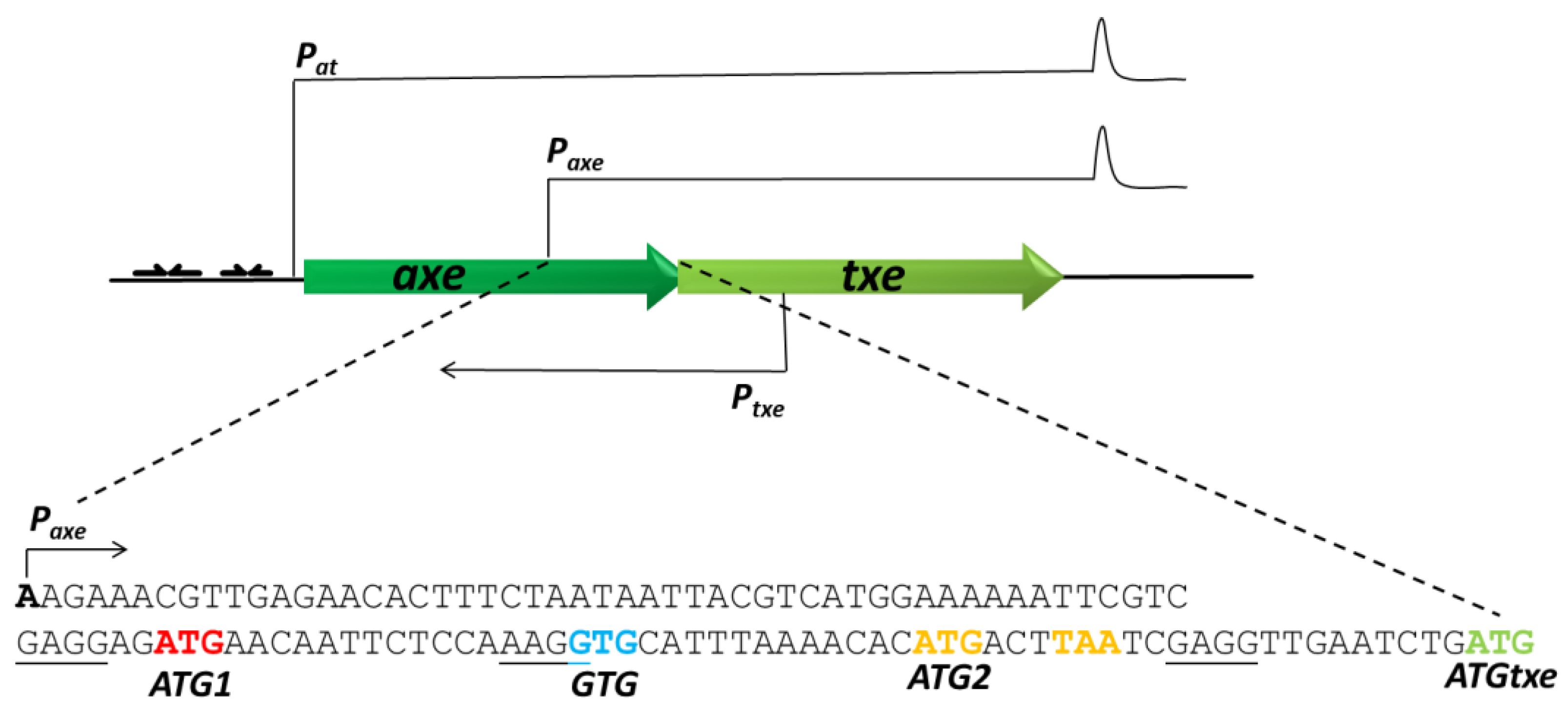
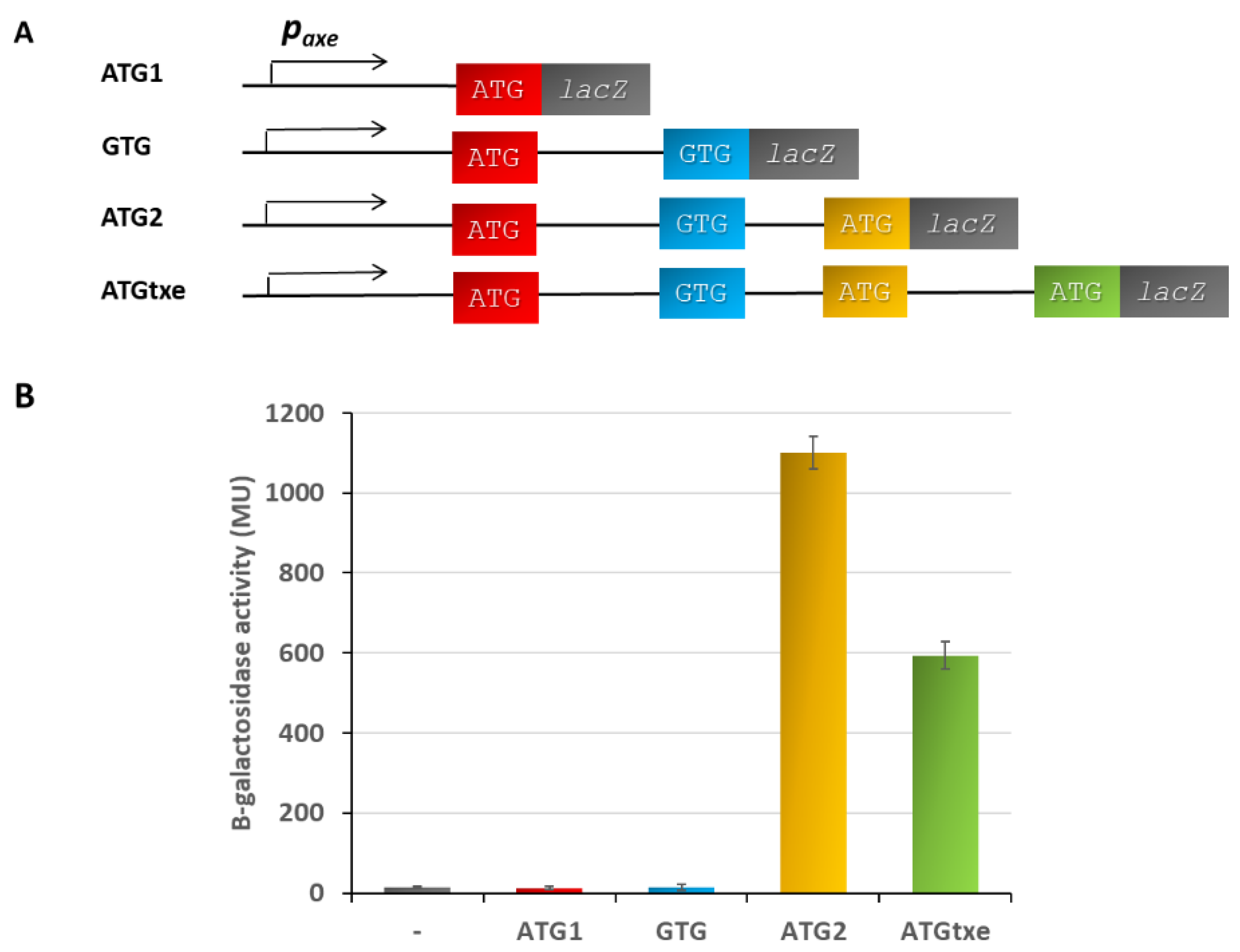
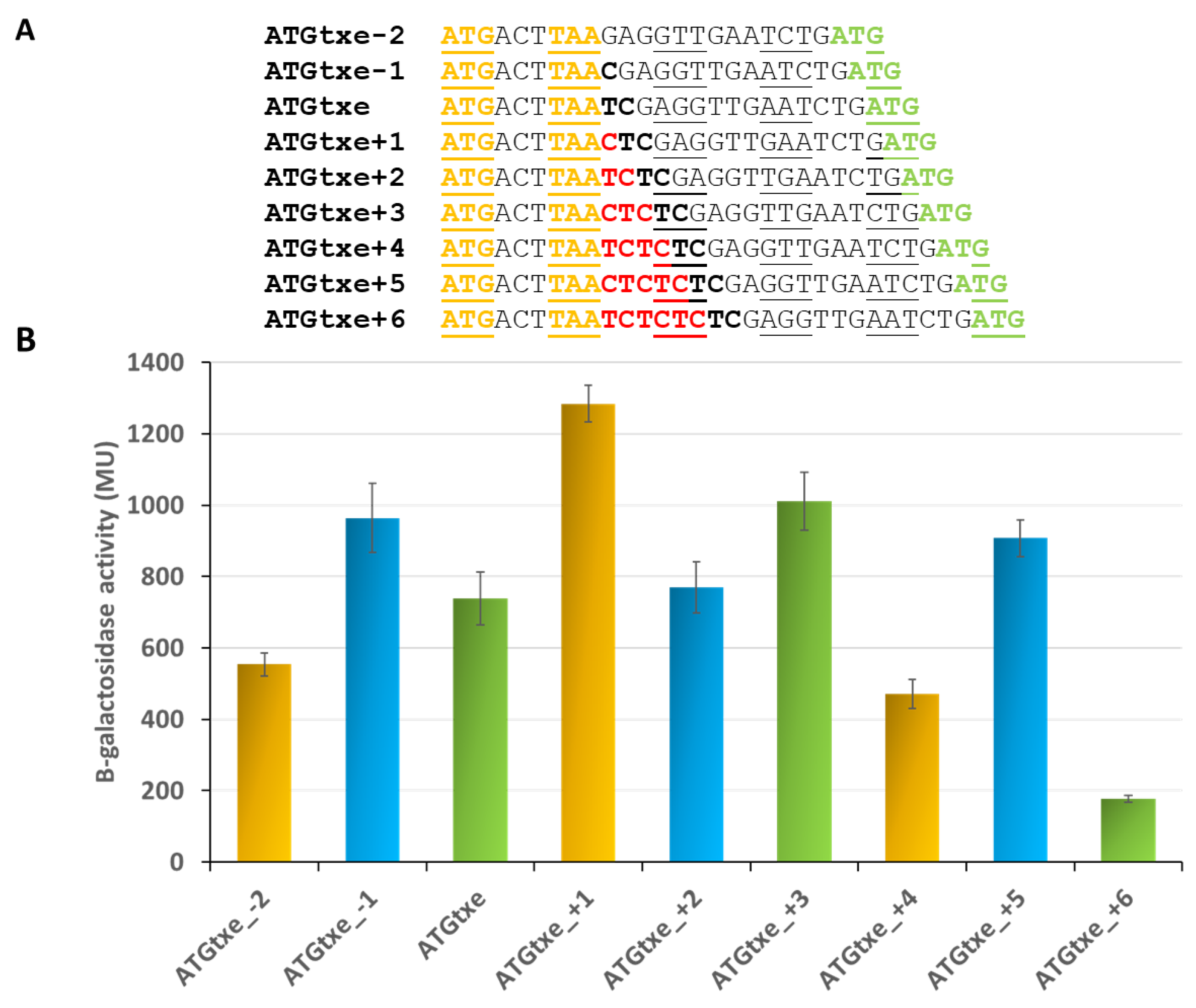
2.1. A Two-Codon Minigene Located within the Leader of paxe Transcript Has a Positive Effect on the Txe Gene Translation
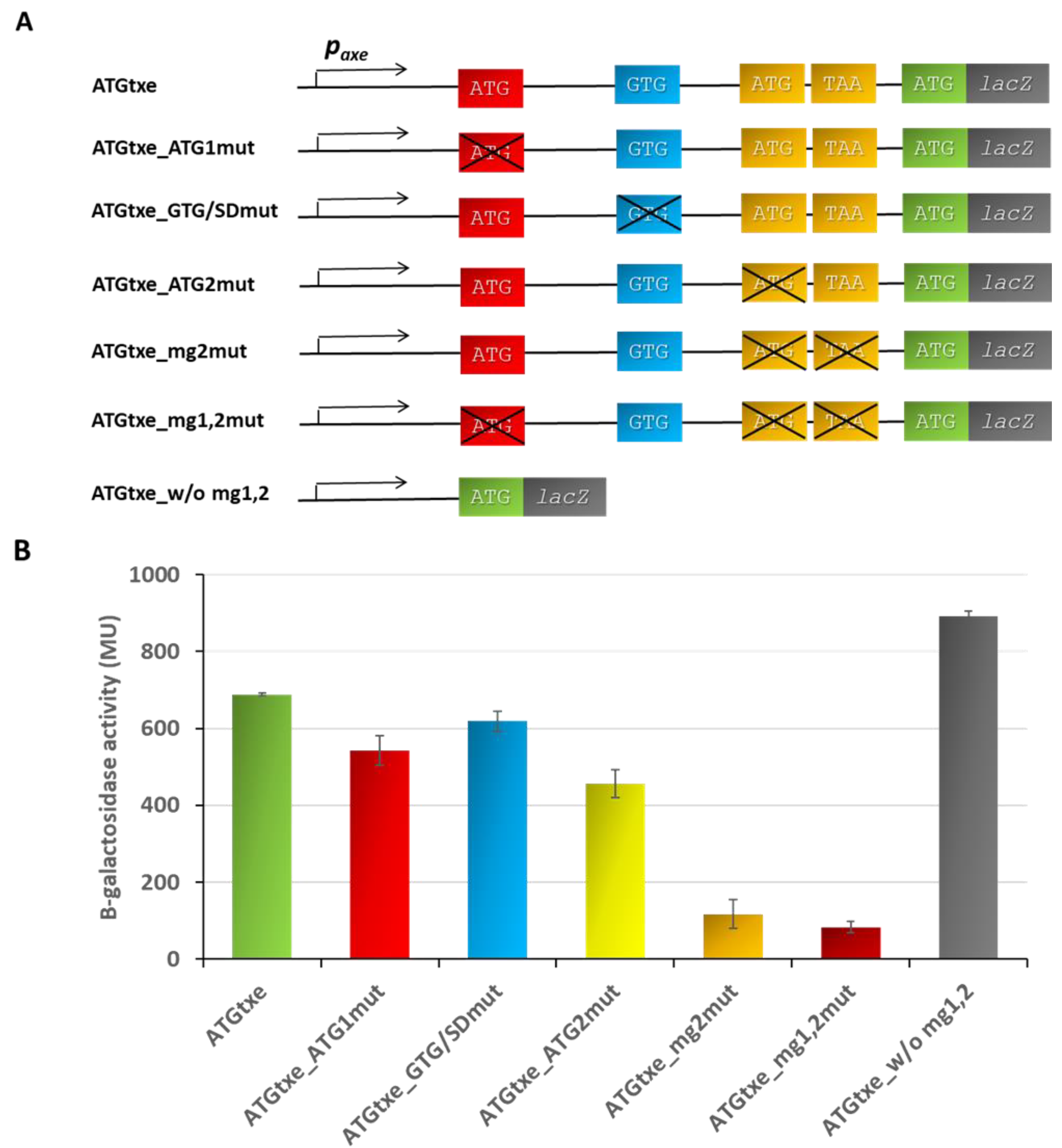
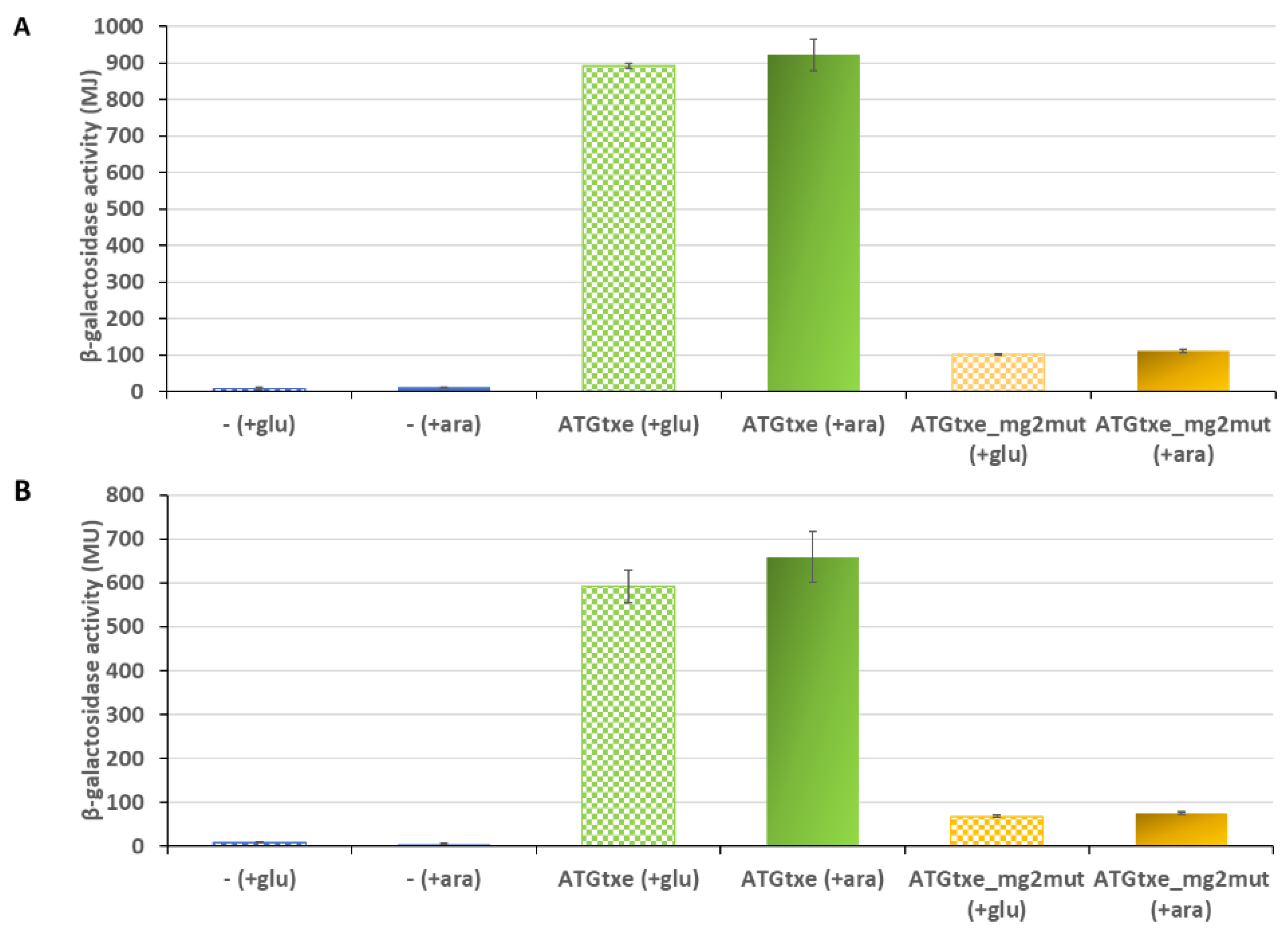
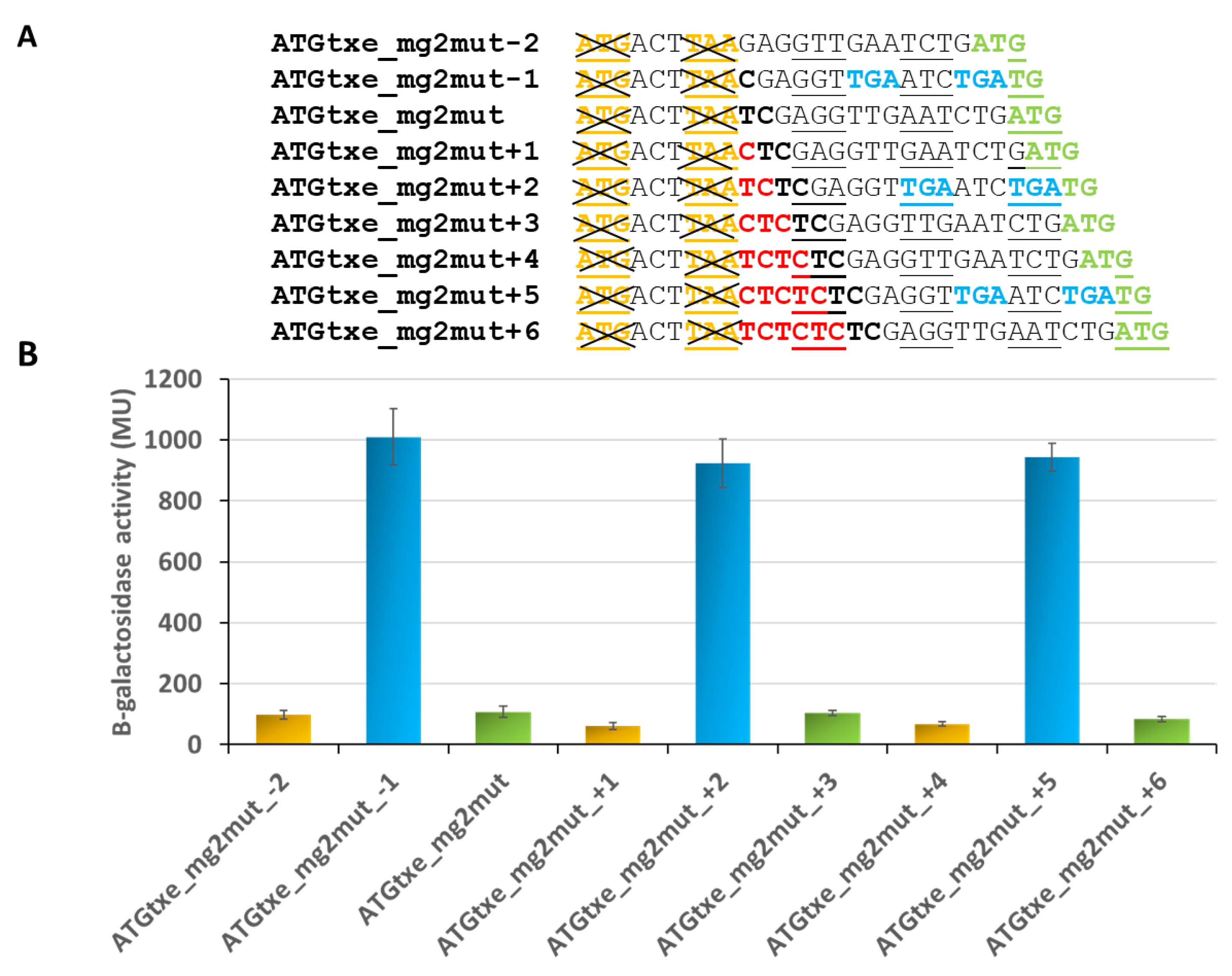
2.2. The mg2 Minigene Acts in Cis to Regulate Translation of Txe and Does Not Display Toxicity towards E. coli Cells
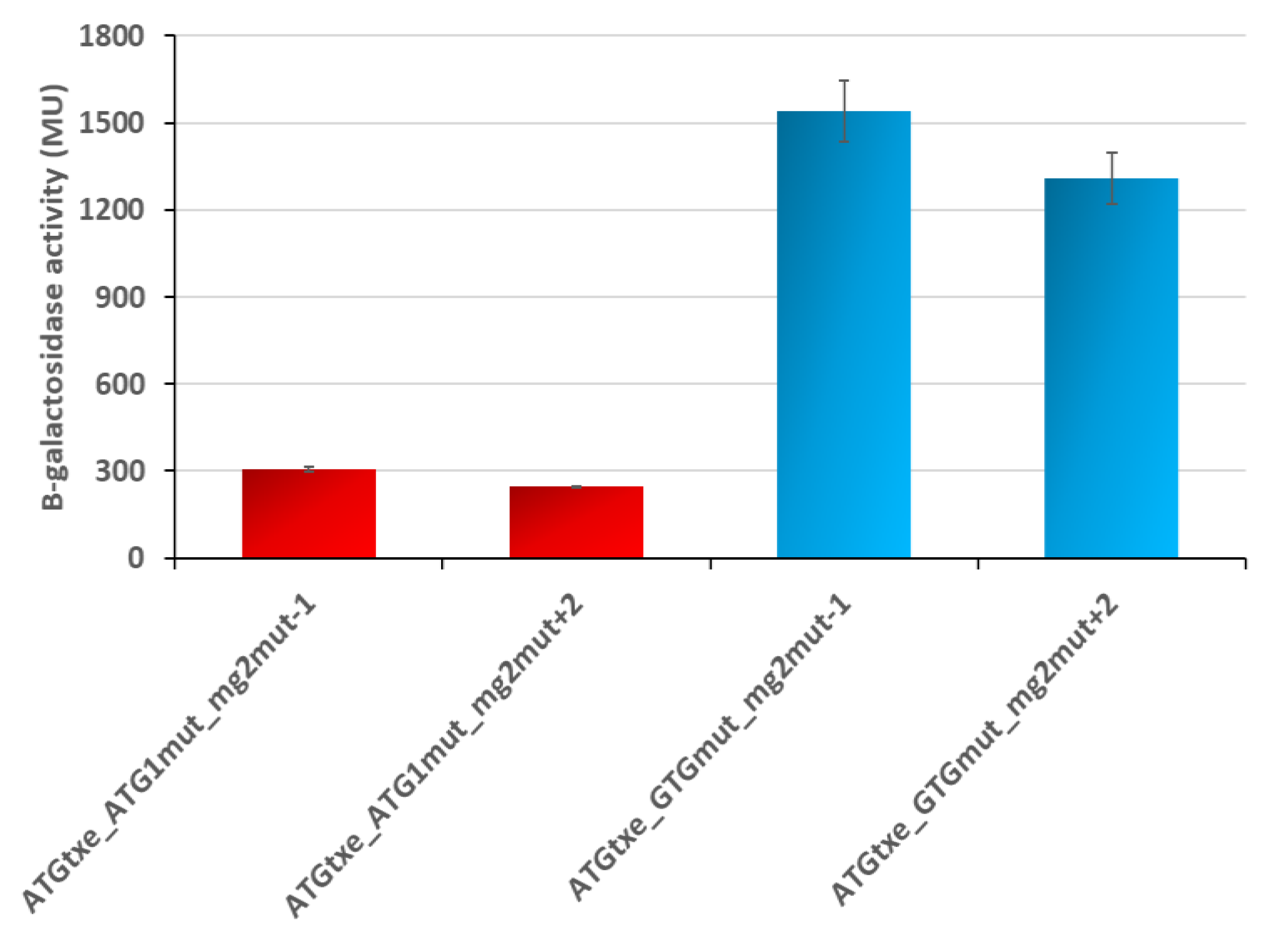
2.3. The Distance between mg2 and Txe, as Well as the Reading Frame of Both Genes, Are Important Features Modulating Expression of the Latter
2.4. Two-Codon Mini-ORFs Are Present within the Antitoxin Gene of the relBE and mazEF TA Families in Diverse Bacterial Species
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids and Oligonucleotides
4.2. Cloning and Mutagenesis Procedures
4.3. Promoter Fusion Studies and β-Galactosidase Assays
4.4. Bacterial Growth Rates
4.5. Bioinformatic Analyses
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yamaguchi, Y.; Park, J.-H.; Inouye, M. Toxin-antitoxin systems in bacteria and archaea. Annu. Rev. Genet. 2011, 45, 61–79. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Comprehensive comparative-genomic analysis of Type 2 toxin-antitoxin systems and related mobile stress response systems in prokaryotes. Biol. Direct 2009, 4, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Leplae, R.; Geeraerts, D.; Hallez, R.; Guglielmini, J.; Drze, P.; Van Melderen, L. Diversity of bacterial type II toxin-antitoxin systems: A comprehensive search and functional analysis of novel families. Nucleic Acids Res. 2011, 39, 5513–5525. [Google Scholar] [CrossRef] [PubMed]
- Kędzierska, B.; Hayes, F. Transcriptional Control of Toxin-Antitoxin Expression: Keeping Toxins Under Wraps Until the Time is Right. In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; Wiley: Hoboken, NJ, USA, 2016; Volume 1, pp. 463–472. [Google Scholar]
- Hayes, F.; Kedzierska, B. Regulating toxin-antitoxin expression: Controlled detonation of intracellular molecular timebombs. Toxins 2014, 6, 337–358. [Google Scholar] [CrossRef]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, targets, and triggers: An overview of toxin-antitoxin biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef]
- Wang, X.; Yao, J.; Sun, Y.C.; Wood, T.K. Type VII toxin/antitoxin classification system for antitoxins that enzymatically neutralize toxins. Trends Microbiol. 2021, 29, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-Y.; Lee, B.-J. Structure, biology, and therapeutic application of toxin–antitoxin systems in pathogenic bacteria. Toxins 2016, 8, 305. [Google Scholar] [CrossRef] [PubMed]
- Fraikin, N.; Goormaghtigh, F.; Van Melderen, L. Type II toxin-antitoxin systems: Evolution and revolutions. J. Bacteriol. 2020, 202, 1–14. [Google Scholar] [CrossRef]
- Grady, R.; Hayes, F. Axe-Txe, a broad-spectrum proteic toxin-antitoxin system specified by a multidrug-resistant, clinical isolate of Enterococcus faecium. Mol. Microbiol. 2003, 47, 1419–1432. [Google Scholar] [CrossRef]
- Boss, L.; Labudda, Ł.; Wegrzyn, G.; Hayes, F.; Kedzierska, B. The Axe-Txe complex of enterococcus faecium presents a multilayered mode of toxin-antitoxin gene expression regulation. PLoS ONE 2013, 8, e73569. [Google Scholar] [CrossRef]
- Kędzierska, B.; Potrykus, K.; Szalewska-Pałasz, A.; Wodzikowska, B. Insights into transcriptional repression of the homologous toxin-antitoxin cassettes YeFm-YoeB and Axe-Txe. Int. J. Mol. Sci. 2020, 21, 9062. [Google Scholar] [CrossRef] [PubMed]
- Miravet-Verde, S.; Ferrar, T.; Espadas-García, G.; Mazzolini, R.; Gharrab, A.; Sabido, E.; Serrano, L.; Lluch-Senar, M. Unraveling the hidden universe of small proteins in bacterial genomes. Mol. Syst. Biol. 2019, 15. [Google Scholar] [CrossRef]
- Orr, M.W.; Mao, Y.; Storz, G.; Qian, S.B. Alternative ORFs and small ORFs: Shedding light on the dark proteome. Nucleic Acids Res. 2021, 48, 1029–1042. [Google Scholar] [CrossRef]
- Weaver, J.; Mohammad, F.; Buskirk, A.R.; Storz, G. Identifying small proteins by ribosome profiling with stalled initiation complexes. MBio 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Beck, H.J.; Fleming, I.M.C.; Janssen, G.R. 5′-terminal AUGs in escherichia coli mRNAs with Shine-Dalgarno sequences: Identification and analysis of their roles in non-canonical translation initiation. PLoS ONE 2016, 11, e0160144. [Google Scholar] [CrossRef]
- Adams, P.P.; Baniulyte, G.; Esnault, C.; Chegireddy, K.; Singh, N.; Monge, M.; Dale, R.K.; Storz, G.; Wade, J.T. Regulatory roles of escherichia coli 5′ utr and orf-internal rnas detected by 3′ end mapping. Elife 2021, 10, 1–33. [Google Scholar] [CrossRef]
- Gollnick, P.; Babitzke, P. Transcription attenuation. Biochim. Biophys. Acta 2002, 1577, 240–250. [Google Scholar] [CrossRef]
- Levin, H.L.; Schachman, H.K. Regulation of aspartate transcarbamoylase synthesis in Escherichia coli: Analysis of deletion mutations in the promoter region of the pyrBI operon (attenuation/galK expression/pKOl/termination of transcription). Proc. Nadl. Acad. Sci. USA 1985, 82, 4643–4647. [Google Scholar] [CrossRef]
- Park, S.-Y.; Cromie, M.J.; Lee, E.; Groisman, E.A. A bacterial mRNA leader that employs different mechanisms to sense disparate intracellular signals. Cell 2010, 142, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Ramu, H.; Mankin, A.; Vazquez-Laslop, N. Programmed drug-dependent ribosome stalling: MicroReview. Mol. Microbiol. 2009, 71, 811–824. [Google Scholar] [CrossRef]
- Yakhnin, H.; Yakhnin, A.V.; Babitzke, P. The trp RNA-binding attenuation protein (TRAP) of Bacillus subtilis regulates translation initiation of ycbK, a gene encoding a putative efflux protein, by blocking ribosome binding. Mol. Microbiol. 2006, 61, 1252–1266. [Google Scholar] [CrossRef]
- Lodato, P.B.; Hsieh, P.-K.; Belasco, J.G.; Kaper, J.B. The ribosome binding site of a mini-ORF protects a T3SS mRNA from degradation by RNase, E. Mol. Microbiol. 2012, 86, 1167. [Google Scholar] [CrossRef]
- Islam, M.S.; Shaw, R.K.; Frankel, G.; Pallen, M.J.; Busby, S.J.W. Translation of a minigene in the 5′ leader sequence of the enterohaemorrhagic Escherichia coli LEE1 transcription unit affects expression of the neighbouring downstream gene. Biochem. J. 2012, 441, 247–253. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Christensen, S.K.; Maenhaut-Michel, G.; Mine, N.; Gottesman, S.; Gerdes, K.; Van Melderen, L. Overproduction of the Lon protease triggers inhibition of translation in Escherichia coli: Involvement of the yefM-yoeB toxin-antitoxin system. Mol. Microbiol. 2004, 51, 1705–1717. [Google Scholar] [CrossRef]
- Miller, J. Experiments in molecular genetics. Astrophys. Space Sci. 1972, 203–209. [Google Scholar]
- Chen, H.; Bjerknes, M.; Kumar, R.; Jay, E. Determination of the optimal aligned spacing between the Shine-Dalgarno sequence and the translation initiation codon of escherichia coli m RNAs. Nucleic Acids Res. 1994, 22, 4953–4957. [Google Scholar] [CrossRef]
- Osterman, I.A.; Evfratov, S.A.; Sergiev, P.V.; Dontsova, O.A. Comparison of mRNA features affecting translation initiation and reinitiation. Nucleic Acids Res. 2013, 41, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Komarova, E.S.; Chervontseva, Z.S.; Osterman, I.A.; Evfratov, S.A.; Rubtsova, M.P.; Zatsepin, T.S.; Semashko, T.A.; Kostryukova, E.S.; Bogdanov, A.A.; Gelfand, M.S.; et al. Influence of the spacer region between the Shine–Dalgarno box and the start codon for fine-tuning of the translation efficiency in Escherichia coli. Microb. Biotechnol. 2020, 13, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Tenson, T.; Vega Herrera, J.; Kloss, P.; Guarneros, G.; Mankin, A.S. Inhibition of translation and cell growth by minigene expression. J. Bacteriol. 1999, 181, 1617–1622. [Google Scholar] [CrossRef]
- Heurgué-Hamard, V.; Dinçbas, V.; Buckingham, R.H.; Ehrenberg, M. Origins of minigene-dependent growth inhibition in bacterial cells. EMBO J. 2000, 19, 2701–2709. [Google Scholar] [CrossRef][Green Version]
- Dinçbas, V.; Heurgué-Hamard, V.; Buckingham, R.H.; Karimi, R.; Ehrenberg, M. Shutdown in protein synthesis due to the expression of mini-genes in bacteria. J. Mol. Biol. 1999, 291, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wei, Y.; Shen, Y.; Li, X.; Zhou, H.; Tai, C.; Deng, Z.; Ou, H.Y. TADB 2.0: An updated database of bacterial type II toxin-antitoxin loci. Nucleic Acids Res. 2018, 46, D749–D753. [Google Scholar] [CrossRef] [PubMed]
- Komarova, A.V.; Tchufistova, L.S.; Supina, E.V.; Boni, I. V Protein S1 counteracts the inhibitory effect of the extended Shine-Dalgarno sequence on translation. RNA 2002, 8, 1137–1147. [Google Scholar] [CrossRef]
- Diwa, A.A.; Jiang, X.; Schapira, M.; Belasco, J.G. Two distinct regions on the surface of an RNA-binding domain are crucial for RNase E function. Mol. Microbiol. 2002, 46, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Komarova, A.V.; Tchufistova, L.S.; Dreyfus, M.; Boni, I.V. AU-rich sequences within 5′ untranslated leaders enhance translation and stabilize mRNA in Escherichia coli. J. Bacteriol. 2005, 187, 1344–1349. [Google Scholar] [CrossRef] [PubMed]
- McDowall, K.J.; Lin-Chao, S.; Cohen, S.N. A+U content rather than a particular nucleotide order determines the specificity of RNase E cleavage. J. Biol. Chem. 1994, 269, 10790–10796. [Google Scholar] [CrossRef]
- Iost, I.; Dreyfus, M. The stability of Escherichia coli lacZ mRNA depends upon the simultaneity of its synthesis and translation. EMBO J. 1995, 14, 3252–3261. [Google Scholar] [CrossRef] [PubMed]
- Wagner’, L.A.; Gesteland, R.F.; Dayhuff, T.J.; Weiss’, R.B. An efficient Shine-Dalgarno sequence but not translation is necessary for lacZ mRNA stability in Escherichia coli. J. BAcrERIOLOGY 1994, 176, 1683–1688. [Google Scholar]
- Arnold, T.E.; Yu, J.; Belasco, J.G. mRNA stabilization by the ompA 5′ untranslated region: Two protective elements hinder distinct pathways for mRNA degradation. RNA 1998, 4, 319–330. [Google Scholar]
- Cruz-Vera, L.R.; Hernández-Ramón, E.; Pérez-Zamorano, B.; Guarneros, G. The rate of peptidyl-tRNA dissociation from the ribosome during minigene expression depends on the nature of the last decoding interaction. J. Biol. Chem. 2003, 278, 26065–26070. [Google Scholar] [CrossRef]
- Menninger, J.R. The accumulation as peptidyl-transfer RNA of isoaccepting transfer RNA families in Escherichia coli with temperature-sensitive peptidyl-transfer RNA hydrolase. J. Biol. Chem. 1978, 253, 6808–6813. [Google Scholar] [CrossRef]
- Rie Heurgué -Hamard, V.; Karimi, R.; Mora, L.; Macdougall, J.; Grentzmann, G.; Ehrenberg, M.; Buckingham, R.H. Ribosome release factor RF4 and termination factor RF3 are involved in dissociation of peptidyl-tRNA from the ribosome. EMBO J. 1998, 17, 808–816. [Google Scholar] [CrossRef]
- Ma, J.; Campbell, A.; Karlin, S. Correlations between Shine-Dalgarno sequences and gene features such as predicted expression levels and operon structures. J. Bacteriol. 2002, 184, 5733–5745. [Google Scholar] [CrossRef]
- van de Guchte, M.; Kok, J.; Venema, G. Distance-dependent translational coupling and interference in Lactococcus lactis. MGG Mol. Gen. Genet. 1991, 227, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Studer, S.M.; Joseph, S. Unfolding of mRNA secondary structure by the bacterial translation initiation complex. Mol. Cell 2006, 22, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef] [PubMed]
- Forloni, M.; Liu, A.Y.; Wajapeyee, N. In vitro mutagenesis using Double-Stranded DNA templates: Selection of mutants with DpnI. Cold Spring Harb. Protoc. 2018, 2018, 231–238. [Google Scholar] [CrossRef]
- Reikofski, J.; Tao, B.Y. Polymerase chain reaction (PCR) techniques for site-directed mutagenesis. Biotechnol. Adv. 1992, 10, 535–547. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | TA Family | Minigene Sequence | Amino Acid $ | Number of Nucleotides # |
|---|---|---|---|---|
| Agrobacterium tumefaciens C58 Atu0810 | relBE | 5′-ATGgagTGA-3′ | Glu | 86 * |
| Archaeoglobus fulgidus DSM 4304 AF2343 | relBE | 5′-ATGagcTGA-3′ | Ser | 12 |
| Bartonella henselae str. Houston-1 BH03400 | relBE | 5′-ATGttgTGA-3′ | Leu | 147 |
| Candidatus Protochlamydia amoebophila UWE25 pc1992 | relBE | 5′-ATGcttTAA-3′ | Leu | 90 |
| Enterococcus faecalis V583 EF0512 | relBE | 5′-ATGattTAG-3′ | Ile | 174 |
| Fusobacterium nucleatum subsp. nucleatum ATCC 25586 FN1099 | relBE | 5′-ATGattTAA-3′ | Ile | 76 * |
| Nitrosomonas europaea ATCC 19718 NE1562 | relBE | 5′-ATGaatTAG-3′ | Asn | 57 |
| Nostoc sp. PCC 7120 (Anabaena sp. PCC 7120) asl2101 | relBE | 5′-ATGactTAA-3′ | Thr | 108 |
| Nostoc sp. PCC 7120 (Anabaena sp. PCC 7120) asl2101 | relBE | 5′-ATGccaTAG-3′ | Pro | 78 |
| Pectobacterium atrosepticum SCRI1043 ECA2757 | relBE | 5′-ATGagaTGA-3′ | Arg | 33 |
| Salmonella enterica subsp. enterica serovar Paratyphi A str. ATCC SPA4249 | relBE | 5′-ATGcatTGA-3′ | His | 57 |
| Salmonella enterica subsp. enterica serovar Typhi str. CT18 STY4788 | relBE | 5′-ATGcacTGA-3′ | His | 57 |
| Salmonella enterica subsp. enterica serovar Typhi str. Ty2 t4483 | relBE | 5′-ATGcacTGA-3′ | His | 57 |
| Salmonella enterica subsp. enterica serovar Typhimurium str. LT2 STM4449 | relBE | 5′-ATGcatTGA-3′ | His | 57 |
| Streptococcus agalactiae 2603V/R SAG2009 | relBE | 5′-ATGgttTAA-3′ | Val | 12 |
| Nitrosomonas europaea ATCC 19718 NE1182 | mazEF | 5′-ATGgatTAG-3′ | Asp | 3 |
| Shigella flexneri 2a str. 301 SF3433 | mazEF | 5′-ATGggcTGA-3′ | Gly | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kędzierska, B.; Potrykus, K. Minigene as a Novel Regulatory Element in Toxin-Antitoxin Systems. Int. J. Mol. Sci. 2021, 22, 13389. https://doi.org/10.3390/ijms222413389
Kędzierska B, Potrykus K. Minigene as a Novel Regulatory Element in Toxin-Antitoxin Systems. International Journal of Molecular Sciences. 2021; 22(24):13389. https://doi.org/10.3390/ijms222413389
Chicago/Turabian StyleKędzierska, Barbara, and Katarzyna Potrykus. 2021. "Minigene as a Novel Regulatory Element in Toxin-Antitoxin Systems" International Journal of Molecular Sciences 22, no. 24: 13389. https://doi.org/10.3390/ijms222413389
APA StyleKędzierska, B., & Potrykus, K. (2021). Minigene as a Novel Regulatory Element in Toxin-Antitoxin Systems. International Journal of Molecular Sciences, 22(24), 13389. https://doi.org/10.3390/ijms222413389