
X-ray Characterization of Conformational Changes of Human Apo- and Holo-Transferrin

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
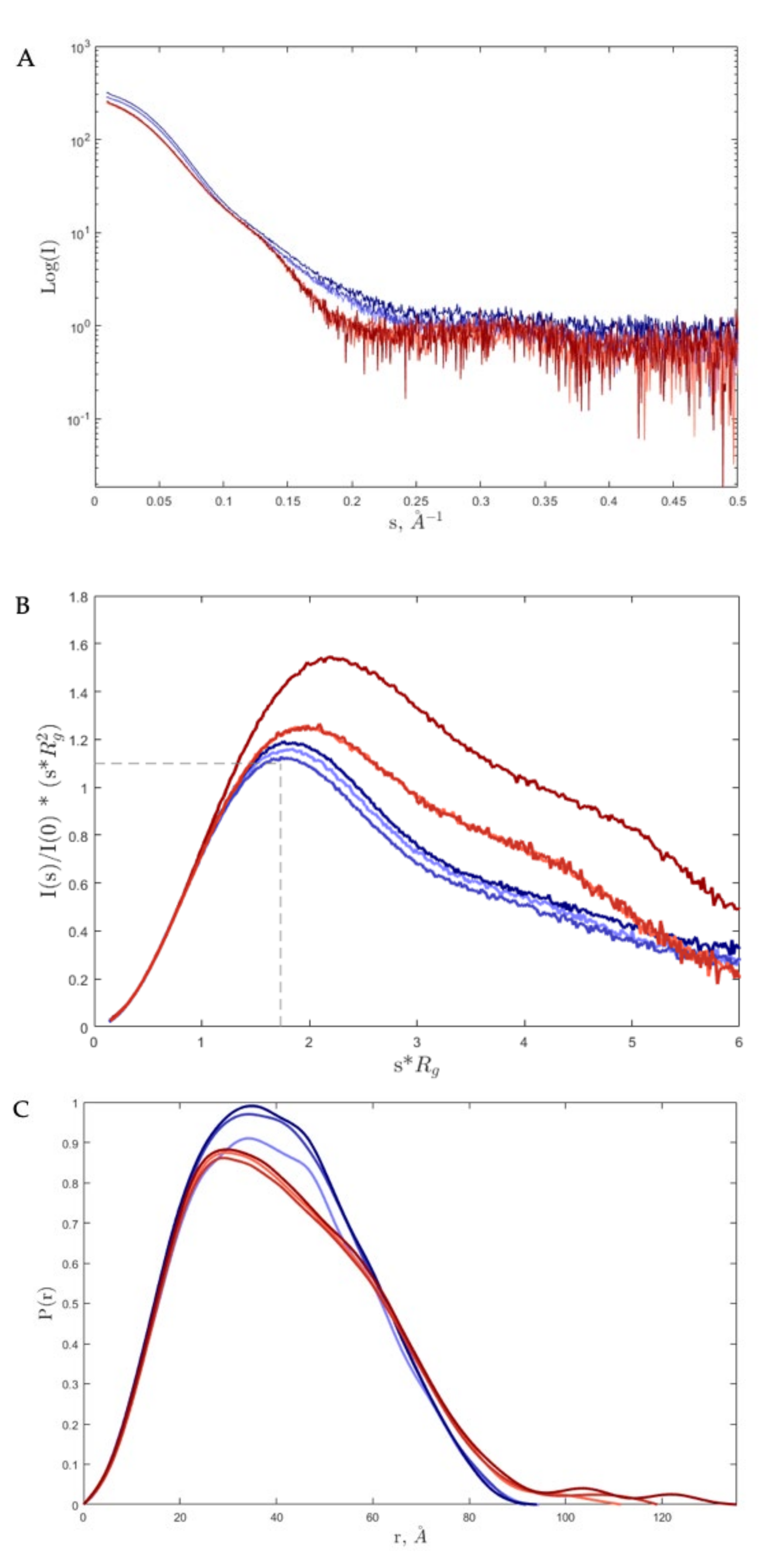
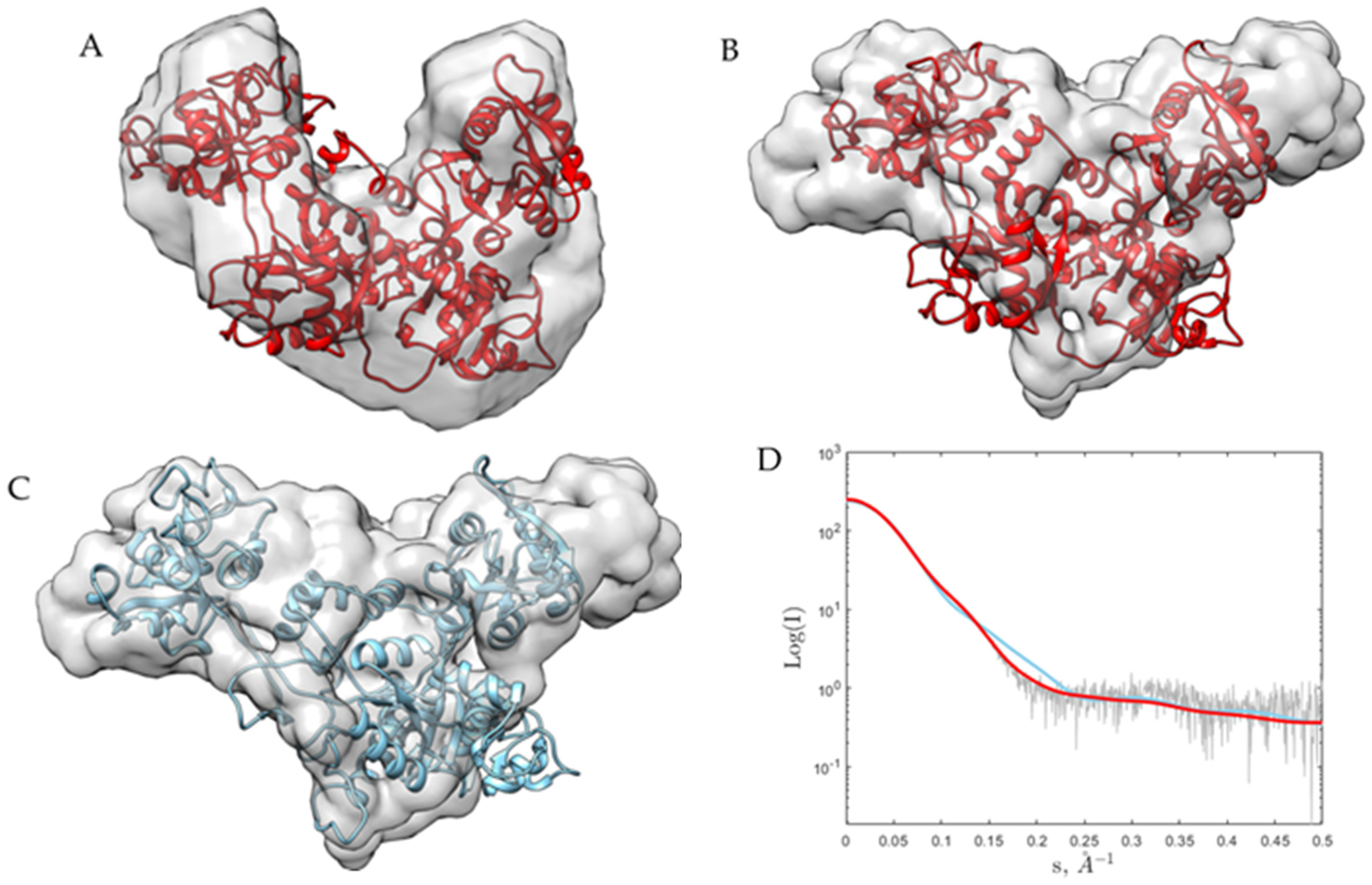
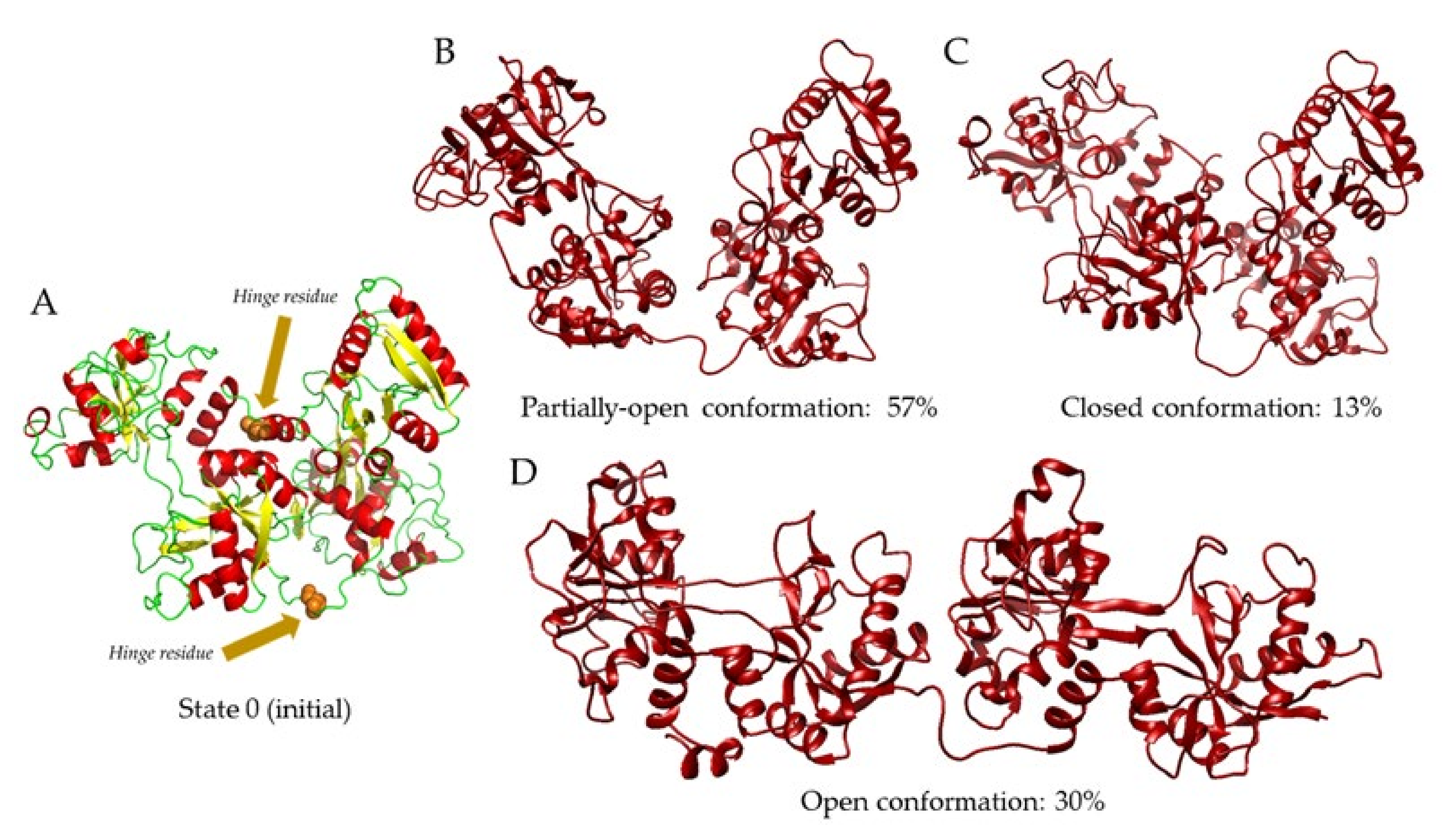
2. Results and Discussion
3. Materials and Methods
3.1. Purification of Transferrin
3.2. Crystallization and Macromolecular Crystallography
3.3. X-ray Diffraction
3.4. Small-Angle X-ray Scattering (SAXS)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, M.; Lai, T.P.; Wang, L.; Zhang, H.; Yang, N.; Sadler, P.J.; Sun, H. “Anion clamp” allows flexible protein to impose coordination geometry on metal ions. Chem. Commun. 2015, 51, 7867–7870. [Google Scholar] [CrossRef]
- Mizutani, K.; Toyoda, M.; Mikami, B. X-ray structures of transferrins and related proteins. Biochim. Biophys. Acta-Gen. Subj. 2012, 1820, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, H. Transferrin and transferrin receptors update. Free Radic. Biol. Med. 2019, 133, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, P.; Alata, W.; Tremblay, C.; Paris-Robidas, S.; Calon, F. Transferrin Receptor-Mediated Uptake at the Blood–Brain Barrier Is Not Impaired by Alzheimer’s Disease Neuropathology. Mol. Pharm. 2019, 16, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Campos-Escamilla, C. The role of transferrins and iron-related proteins in brain iron transport: Applications to neurological diseases. Adv. Protein Chem. Struct. Biol. 2021, 123, 133–162. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.A.; Hanck-Silva, G.; Baveloni, F.G.; Junior, J.A.O.; De Lima, F.T.; Eloy, J.O.; Chorilli, M. A Review of Properties, Delivery Systems and Analytical Methods for the Characterization of Monomeric Glycoprotein Transferrin. Crit. Rev. Anal. Chem. 2020, 51, 399–410. [Google Scholar] [CrossRef]
- Śliwińska-Hill, U. Interaction of imatinib mesylate with human serum transferrin: The comparative spectroscopic studies. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2017, 173, 468–475. [Google Scholar] [CrossRef]
- Śliwińska-Hill, U. Spectroscopic studies of simultaneous binding of cyclophosphamide and imatinib mesylate to human holo-transferrin. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 252, 119538. [Google Scholar] [CrossRef]
- Shamsi, A.; Mohammad, T.; Khan, M.S.; Shahwan, M.; Husain, F.M.; Rehman, T.; Hassan, I.; Ahmad, F.; Islam, A. Unraveling Binding Mechanism of Alzheimer’s Drug Rivastigmine Tartrate with Human Transferrin: Molecular Docking and Multi-Spectroscopic Approach towards Neurodegenerative Diseases. Biomolecules 2019, 9, 495. [Google Scholar] [CrossRef] [Green Version]
- Shamsi, A.; Shahwan, M.; Ahamad, S.; Hassan, I.; Ahmad, F.; Islam, A. Spectroscopic, calorimetric and molecular docking insight into the interaction of Alzheimer’s drug donepezil with human transferrin: Implications of Alzheimer’s drug. J. Biomol. Struct. Dyn. 2019, 38, 1094–1102. [Google Scholar] [CrossRef]
- Amroabadi, M.K.; Taheri-Kafrani, A.; Saremi, L.H.; Rastegari, A.A. Spectroscopic studies of the interaction between alprazolam and apo-human serum transferrin as a drug carrier protein. Int. J. Biol. Macromol. 2018, 108, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Zhang, H.; Wang, M.; Hao, Q.; Sun, H. Iron and bismuth bound human serum transferrin reveals a partially-opened conformation in the N-lobe. Sci. Rep. 2012, 2, 999. [Google Scholar] [CrossRef] [Green Version]
- Wally, J.; Halbrooks, P.J.; Vonrhein, C.; Rould, M.A.; Everse, S.J.; Mason, A.B.; Buchanan, S.K. The Crystal Structure of Iron-free Human Serum Transferrin Provides Insight into Inter-lobe Communication and Receptor Binding. J. Biol. Chem. 2006, 281, 24934–24944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtin, J.P.; Wang, M.; Cheng, T.; Jin, L.; Sun, H. The role of citrate, lactate and transferrin in determining titanium release from surgical devices into human serum. J. Biol. Inorg. Chem. 2018, 23, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.C.; Kim, H.; Ferguson, R.; Lockart, M.; Vincent, J.B. Significance of conformation changes during the binding and release of chromium(III) from human serum transferrin. J. Inorg. Biochem. 2020, 206, 111040. [Google Scholar] [CrossRef] [PubMed]
- Kulakova, A.; Indrakumar, S.; Sønderby, P.; Gentiluomo, L.; Streicher, W.; Roessner, D.; Frieß, W.; Peters, G.H.J.; Harris, P. Structural-stability studies on recombinant human transferrin. Biorxiv 2019, 742957, preprint. [Google Scholar] [CrossRef]
- Otálora, F.; Gavira, J.A.; Ng, J.D.; García-Ruiz, J.M. Counterdiffusion methods applied to protein crystallization. Prog. Biophys. Mol. Biol. 2009, 101, 26–37. [Google Scholar] [CrossRef]
- Gavira, J.A. Current trends in protein crystallization. Arch. Biochem. Biophys. 2016, 602, 3–11. [Google Scholar] [CrossRef]
- Gavira, J.A.; Otálora, F.; González-Ramírez, L.A.; Melero, E.; Van Driessche, A.E.; García-Ruíz, J.M. On the Quality of Protein Crystals Grown under Diffusion Mass-transport Controlled Regime (I). Crystals 2020, 10, 68. [Google Scholar] [CrossRef] [Green Version]
- Ng, J.D.; Gavira, J.A.; García-Ruiz, J.M. Protein crystallization by capillary counterdiffusion for applied crystallographic structure determination. J. Struct. Biol. 2003, 142, 218–231. [Google Scholar] [CrossRef]
- Mesters, J.R.; Hilgenfeld, R. Protein Glycosylation, Sweet to Crystal Growth? Cryst. Growth Des. 2007, 7, 2251–2253. [Google Scholar] [CrossRef]
- Gräwert, T.W.; Svergun, D.I. Structural Modeling Using Solution Small-Angle X-ray Scattering (SAXS). J. Mol. Biol. 2020, 432, 3078–3092. [Google Scholar] [CrossRef] [PubMed]
- Guinier, A. La diffraction des rayons X aux très petits angles: Application à l’étude de phénomènes ultramicroscopiques. Ann. Phys. 1939, 11, 161–237. [Google Scholar] [CrossRef]
- Glatter, O. A new method for the evaluation of small-angle scattering data. J. Appl. Crystallogr. 1977, 10, 415–421. [Google Scholar] [CrossRef]
- Svergun, D.I. Restoring low resolution structure of biological macromolecules from solution scattering using simulated annealing. Biophys. J. 1999, 76, 2879–2886. [Google Scholar] [CrossRef] [Green Version]
- Panjkovich, A.; Svergun, D.I. Deciphering conformational transitions of proteins by small angle X-ray scattering and normal mode analysis. Phys. Chem. Chem. Phys. 2015, 18, 5707–5719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneidman-Duhovny, D.; Hammel, M.; Tainer, J.; Sali, A. FoXS, FoXSDock and MultiFoXS: Single-state and multi-state structural modeling of proteins and their complexes based on SAXS profiles. Nucleic Acids Res. 2016, 44, W424–W429. [Google Scholar] [CrossRef]
- Emekli, U.; Schneidman-Duhovny, D.; Wolfson, H.J.; Nussinov, R.; Haliloglu, T. HingeProt: Automated prediction of hinges in protein structures. Proteins Struct. Funct. Bioinform. 2007, 70, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Bogaerts, J.; Johannessen, C. On/off resonance Raman optical activity of human serum transferrin. J. Raman Spectrosc. 2019, 50, 641–646. [Google Scholar] [CrossRef]
- Flores-Ibarra, A.; Campos-Escamilla, C.; Guerra, Y.; Rudiño-Piñera, E.; Demitri, N.; Polentarutti, M.; Cuéllar-Cruz, M.; Moreno, A. Novel Devices for Transporting Protein Crystals to the Synchrotron Facilities and Thermal Protection of Protein Crystals. Crystals 2018, 8, 340. [Google Scholar] [CrossRef] [Green Version]
- Russi, S.; Song, J.; McPhillips, S.E.; Cohen, A.E. The Stanford Automated Mounter: Pushing the limits of sample exchange at the SSRL macromolecular crystallography beamlines. J. Appl. Crystallogr. 2016, 49, 622–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, A.; Moorhead, P.; McPhillips, S.E.; Song, J.; Sharp, K.; Taylor, J.R.; Adams, P.; Sauter, N.K.; Soltis, S.M. Web-Ice: Integrated data collection and analysis for macromolecular crystallography. J. Appl. Crystallogr. 2008, 41, 176–184. [Google Scholar] [CrossRef] [Green Version]
- Minor, W.; Cymborowski, M.; Otwinowski, Z.; Chruszcz, M. HKL-3000: The integration of data reduction and structure solution—From diffraction images to an initial model in minutes. Acta Crystallogr. Sect. D Biol. Crystallogr. 2006, 62, 859–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabsch, W. Research papers XDS research papers. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Evans, P.R.; Murshudov, G.N. How good are my data and what is the resolution? Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.; Krissinel, E.B.; Leslie, A.G.W.; McCoy, A.; et al. Overview of theCCP4 suite and current developments. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vagin, A.A.; Teplyakov, A. MOLREP: An Automated Program for Molecular Replacement. J. Appl. Crystallogr. 1997, 30, 1022–1025. [Google Scholar] [CrossRef]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonine, P.V.; Mustyakimov, M.; Grosse-Kunstleve, R.W.; Moriarty, N.W.; Langan, P.; Adams, P. Joint X-ray and neutron refinement with phenix.refine. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 1153–1163. [Google Scholar] [CrossRef] [Green Version]
- Chen, V.B.; Arendall, W.B.I.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Cryst. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Guranovic, V.; Dutta, S.; Feng, Z.; Berman, H.M.; Westbrook, J. Automated and accurate deposition of structures solved by X-ray diffraction to the Protein Data Bank. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 1833–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernot, P.; Theveneau, P.; Giraud, T.; Fernandes, R.N.; Nurizzo, D.; Spruce, D.; Surr, J.; McSweeney, S.; Round, A.; Felisaz, F.; et al. New beamline dedicated to solution scattering from biological macromolecules at the ESRF. J. Phys. Conf. Ser. 2010, 247, 012009. [Google Scholar] [CrossRef]
- Blanchet, C.E.; Spilotros, A.; Schwemmer, F.; Graewert, M.A.; Kikhney, A.; Jeffries, C.M.; Frank, S.; Mark, D.; Zengerle, R.; Cipriani, F.; et al. Versatile sample environments and automation for biological solution X-ray scattering experiments at the P12 beamline (PETRA III, DESY). J. Appl. Crystallogr. 2015, 48, 431–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Round, A.; Felisaz, F.; Fodinger, L.; Gobbo, A.; Huet, J.; Villard, C.; Blanchet, C.E.; Pernot, P.; McSweeney, S.; Rössle, M.; et al. BioSAXS Sample Changer: A robotic sample changer for rapid and reliable high-throughput X-ray solution scattering experiments. Acta Crystallogr. Sect. D Biol. Crystallogr. 2015, 71, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Franke, D.; Kikhney, A.G.; Svergun, D.I. Automated acquisition and analysis of small angle X-ray scattering data. Nucl. Instrum. Methods Phys. Res. Sect. A Accel. Spectrometers Detect. Assoc. Equip. 2012, 689, 52–59. [Google Scholar] [CrossRef]
- Konarev, P.; Volkov, V.V.; Sokolova, A.; Koch, M.H.J.; Svergun, D.I. PRIMUS: A Windows PC-based system for small-angle scattering data analysis. J. Appl. Crystallogr. 2003, 36, 1277–1282. [Google Scholar] [CrossRef]
- Förster, S.; Apostol, L.; Bras, W. Scatter: Software for the analysis of nano- and mesoscale small-angle scattering. J. Appl. Crystallogr. 2010, 43, 639–646. [Google Scholar] [CrossRef]
- Svergun, D.I.; Barberato, C.; Koch, M.H.J. CRYSOL– a Program to Evaluate X-ray Solution Scattering of Biological Macromolecules from Atomic Coordinates. J. Appl. Crystallogr. 1995, 28, 768–773. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Kikhney, A.G.; Borges, C.R.; Molodenskiy, D.S.; Jeffries, C.M.; Svergun, D.I. SASBDB: Towards an automatically curated and validated repository for biological scattering data. Protein Sci. 2019, 29, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Protein | Apo-Transferrin |
|---|---|
| Ligand | NAG |
| PDB identifier | 7Q1L |
| Data collection | |
| Beamline | BL14-1 |
| Space Group | P 21 21 21 |
| Cell dimensions | |
| a, b, c (Å) | 87.63, 102.15, 199.97 |
| ASU | 2 |
| Resolution (Å) | 37.24– 3.0 (3.107– 3.0) |
| Rmerge (%) | 9.32 (116.0) |
| I/σI | 12.13 (1.16) |
| Completeness (%) | 99.81 (99.82) |
| Unique reflections | 36660 (3606) |
| Multiplicity | 7.3 (8.0) |
| CC(1/2) | 0.999 (0.559) |
| Refinement | |
| Resolution (Å) | 37.24– 3.00 |
| Rwork/Rfree (%) | 22.03 / 25.01 |
| No. atoms | 10716 |
| Protein | 10592 |
| Ligands | 235 |
| Water | 7 |
| B-factor (Å2) | 120.52 |
| R.m.s deviations | |
| Bond lengths (Å) | 0.005 |
| Bond angles (0) | 0.85 |
| Ramachandran (%) | |
| Favored | 92.95 |
| Allowed | 6.75 |
| Outliers | 0.30 |
| Data Collection Parameters | ||||||
|---|---|---|---|---|---|---|
| Beamline | BM29, ESRF | |||||
| Detector | Pilatus3 2M in vacuum | |||||
| Beam size (mm) | 0.2 × 0.2 | |||||
| Energy (keV) | 12.5 | |||||
| Sample-to-detector distance (mm) | 2.867 | |||||
| s range (A−1) | 0.0025–0.6 | |||||
| Exposure time (s) | 1 | |||||
| Temperature (K) | 293 | |||||
| Data collection mode | In batch, automated sample changer | |||||
| apo-Tf | holo-Tf | |||||
| Structural parameters | pH 8.0 | pH 7.0 | pH 5.5 | pH 8.0 | pH 7.0 | pH 5.5 |
| Concentration range (mg mL−1) | 5 | 5 | 5 | 5 | 5 | 5 |
| s Interval for Fourier inversion (Å−1) | 0.014–0.258 | 0.015–0.258 | 0.017–0.258 | 0.018–0.247 | 0.021–0.249 | 0.015–0.248 |
| Rg (from P(r)) (Å) | 31.2 ± 0.02 | 31.2 ± 0.02 | 31.2 ± 0.02 | 33.0 ± 0.05 | 33.2 ± 0.05 | 34.2 ± 0.06 |
| Rg (from Guinier approximation) (Å) | 31.1 ± 0.00 | 31.1 ± 0.00 | 31.1 ± 0.01 | 32.4 ± 0.01 | 33.4 ± 0.01 | 33.7 ± 0.01 |
| sRg limits (from Guinier approximation) | 0.44–1.29 | 0.44–1.28 | 0.21–1.30 | 0.56–1.28 | 0.66–1.30 | 0.48–1.30 |
| Dmax (Å) | 94.2 | 96.4 | 98.4 | 131.0 | 135 | 146 |
| Porod coefficient | 3.7 | 3.7 | 3.7 | 3.5 | 3.0 | 3.0 |
| Porod volume estimate (nm3) | 104 | 106 | 106 | 106 | 103 | 105 |
| Model excluded volume (nm3) | 119 | |||||
| Molecular Mass (kDa) from: | ||||||
| Porod volume | 66.0 | |||||
| Excluded volume (×0.5) | 60.0 | |||||
| From sequence | 77.0 | |||||
| Modeling Ambiguity | 2.1 (might be ambiguous) | 2.4 (might be ambiguous) | ||||
| SASBDB code | SASDMP7 | SASDMN7 | SASDMM7 | SASDMS7 | SASDMR7 | SASDMM7 |
| SAXS software employed | ||||||
| Primary data reduction | ESRF online software tools | |||||
| Data processing | ScÅtter IV/ATSAS 3.0.4 | |||||
| Computation of model intensities | CRYSOL (ATSAS) | |||||
| Modeling | DAMMIN (ATSAS), SREFLEX (ATSAS), MultiFoxs | |||||
| apo-Tf | |||||
|---|---|---|---|---|---|
| pH 8.0 | |||||
| State | Conformation | Fraction | Rg | Dmax | χ2 |
| 0 | 0 | 1 | 29.5 | 102.4 | 6.5 |
| 1 | 1 | 1 | 30.4 | 103.4 | 3.6 |
| 2 | 1 | 0.9 | 29.9 | 103.2 | 1.6 |
| 2 | 0.1 | 36.7 | 127.1 | ||
| pH 7.0 | |||||
| State | Conformation | Fraction | Rg | Dmax | χ2 |
| 0 | 0 | 1 | 29.5 | 102.4 | 5.8 |
| 1 | 1 | 1 | 30.2 | 103.2 | 3.2 |
| 2 | 1 | 0.88 | 29.9 | 103.4 | 1.2 |
| 2 | 0.12 | 33.8 | 119.6 | ||
| pH 5.5 | |||||
| State | Conformation | Fraction | Rg | Dmax | χ2 |
| 0 | 0 | 1 | 29.5 | 102.4 | 8.5 |
| 1 | 1 | 1 | 30.1 | 103.1 | 6.3 |
| 2 | 1 | 0.91 | 30.1 | 103.1 | 1.4 |
| 2 | 0.09 | 33.8 | 119.9 | ||
| holo-Tf | |||||
| pH 8.0 | |||||
| State | Conformation | Fraction | Rg | Dmax | χ2 |
| 0 | 0 | 1 | 29.5 | 102.4 | 28.6 |
| 1 | 1 | 1 | 31.7 | 106.4 | 7.3 |
| 3 | 1 | 0.5 | 31.6 | 103.0 | 4.1 |
| 2 | 0.26 | 29.8 | 102.5 | ||
| 3 | 0.24 | 37.31 | 127.7 | ||
| pH 7.0 | |||||
| State | Conformation | Fraction | Rg | Dmax | χ2 |
| 0 | 0 | 1 | 29.5 | 102.4 | 45.6 |
| 1 | 1 | 1 | 31.9 | 109.6 | 13.2 |
| 3 | 1 | 0.44 | 31.9 | 104.0 | 6.43 |
| 2 | 0.35 | 30.4 | 103.9 | ||
| 3 | 0.21 | 37.5 | 132.9 | ||
| pH 5.5 | |||||
| State | Conformation | Fraction | Rg | Dmax | χ2 |
| 0 | 0 | 1 | 29.5 | 102.4 | 33.6 |
| 1 | 1 | 1 | 31.9 | 109.9 | 4.59 |
| 3 | 1 | 0.57 | 31.4 | 102.4 | 2.4 |
| 2 | 0.13 | 29.5 | 101.9 | ||
| 3 | 0.30 | 37.4 | 132.6 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos-Escamilla, C.; Siliqi, D.; Gonzalez-Ramirez, L.A.; Lopez-Sanchez, C.; Gavira, J.A.; Moreno, A. X-ray Characterization of Conformational Changes of Human Apo- and Holo-Transferrin. Int. J. Mol. Sci. 2021, 22, 13392. https://doi.org/10.3390/ijms222413392
Campos-Escamilla C, Siliqi D, Gonzalez-Ramirez LA, Lopez-Sanchez C, Gavira JA, Moreno A. X-ray Characterization of Conformational Changes of Human Apo- and Holo-Transferrin. International Journal of Molecular Sciences. 2021; 22(24):13392. https://doi.org/10.3390/ijms222413392
Chicago/Turabian StyleCampos-Escamilla, Camila, Dritan Siliqi, Luis A. Gonzalez-Ramirez, Carmen Lopez-Sanchez, Jose Antonio Gavira, and Abel Moreno. 2021. "X-ray Characterization of Conformational Changes of Human Apo- and Holo-Transferrin" International Journal of Molecular Sciences 22, no. 24: 13392. https://doi.org/10.3390/ijms222413392
APA StyleCampos-Escamilla, C., Siliqi, D., Gonzalez-Ramirez, L. A., Lopez-Sanchez, C., Gavira, J. A., & Moreno, A. (2021). X-ray Characterization of Conformational Changes of Human Apo- and Holo-Transferrin. International Journal of Molecular Sciences, 22(24), 13392. https://doi.org/10.3390/ijms222413392