
The Small-Molecule Wnt Inhibitor ICG-001 Efficiently Inhibits Colorectal Cancer Stemness and Metastasis by Suppressing MEIS1 Expression

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Increased Stemness and Wnt Activation Are Associated with CRC Metastasis
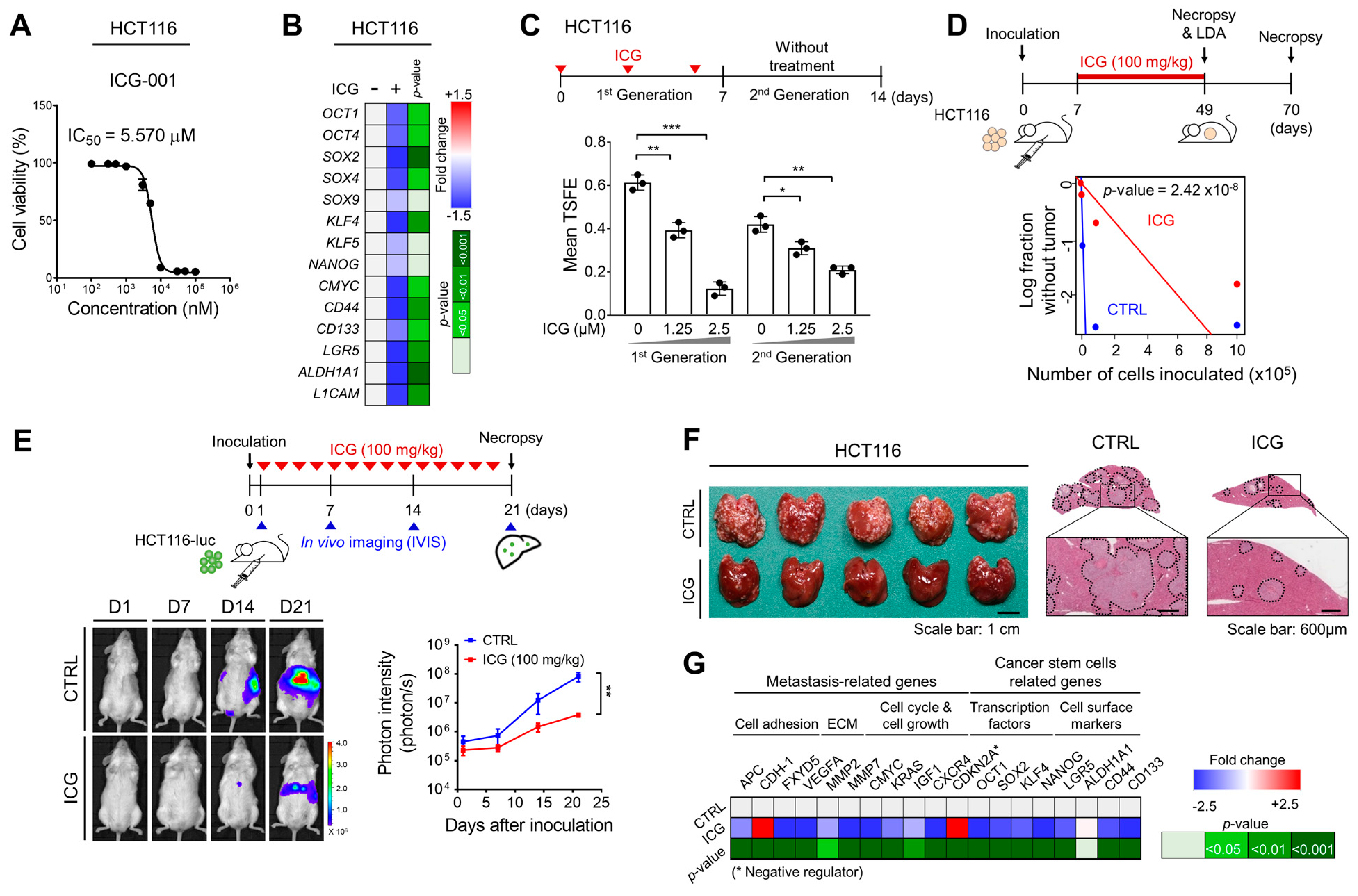
2.2. Targeting Wnt Signaling with ICG-001 Efficiently Attenuates CRC Stemness and Metastasis
2.3. MEIS1 Is a Potential Target Gene of ICG-001, Which Is Associated with CRC Stemness and Clinical Malignancy
2.4. MEIS1 Overexpression Enhances the Self-Renewal Capacity of CSCs and Metastasis of CRC
3. Discussion
4. Materials and Methods
4.1. Bioinformatics Analysis
4.2. Cell Culture and Reagents
4.3. Sphere Formation Assay
4.4. Reverse Transcription–Quantitative Polymerase Chain Reaction (RT-qPCR)
4.5. Western Blot Analysis
4.6. Immunofluorescence Assay
4.7. Animal Study
4.8. Small Interfering RNA (siRNA)-Mediated Knockdown
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.B.; Pastushenko, I.; Skibinski, A.; Blanpain, C.; Kuperwasser, C. Phenotypic plasticity: Driver of cancer initiation, progression, and therapy resistance. Cell Stem Cell 2019, 24, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef]
- Jang, G.-B.; Hong, I.-S.; Kim, R.-J.; Lee, S.-Y.; Park, S.-J.; Lee, E.-S.; Park, J.H.; Yun, C.-H.; Chung, J.-U.; Lee, K.-J. Wnt/β-Catenin small-molecule inhibitor CWP232228 preferentially inhibits the growth of breast cancer stem-like cells. Cancer Res. 2015, 75, 1691–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, G.-B.; Kim, J.-Y.; Cho, S.-D.; Park, K.-S.; Jung, J.-Y.; Lee, H.-Y.; Hong, I.-S.; Nam, J.-S. Blockade of Wnt/β-catenin signaling suppresses breast cancer metastasis by inhibiting CSC-like phenotype. Sci. Rep. 2015, 5, 12465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-Y.; Lee, H.-Y.; Park, K.-K.; Choi, Y.-K.; Nam, J.-S.; Hong, I.-S. CWP232228 targets liver cancer stem cells through Wnt/β-catenin signaling: A novel therapeutic approach for liver cancer treatment. Oncotarget 2016, 7, 20395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-Y.; Kim, J.-Y.; Choi, J.-H.; Kim, J.-H.; Lee, C.-J.; Singh, P.; Sarkar, S.; Baek, J.-H.; Nam, J.-S. Inhibition of LEF1-mediated DCLK1 by niclosamide attenuates colorectal cancer stemness. Clin. Cancer Res. 2019, 25, 1415–1429. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.C.C.; Lo, E.S.F.; Lee, K.C.; Chan, J.K.; Hsiao, W.W. Prognostic and diagnostic significance of β-catenin nuclear immunostaining in colorectal cancer. Clin. Cancer Res. 2004, 10, 1401–1408. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, L.; Felipe De Sousa, E.M.; Van Der Heijden, M.; Cameron, K.; De Jong, J.H.; Borovski, T.; Tuynman, J.B.; Todaro, M.; Merz, C.; Rodermond, H. Wnt activity defines colon cancer stem cells and is regulated by the microenvironment. Nat. Cell Biol. 2010, 12, 468–476. [Google Scholar] [CrossRef]
- Emami, K.H.; Nguyen, C.; Ma, H.; Kim, D.H.; Jeong, K.W.; Eguchi, M.; Moon, R.T.; Teo, J.-L.; Oh, S.W.; Kim, H.Y. A small molecule inhibitor of β-catenin/cyclic AMP response element-binding protein transcription. Proc. Natl. Acad. Sci. USA 2004, 101, 12682–12687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulter, L.; Guest, R.V.; Kendall, T.J.; Wilson, D.H.; Wojtacha, D.; Robson, A.J.; Ridgway, R.A.; Samuel, K.; Van Rooijen, N.; Barry, S.T. WNT signaling drives cholangiocarcinoma growth and can be pharmacologically inhibited. J. Clin. Investig. 2015, 125, 1269–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Chen, H.; Zheng, P.; Zheng, Y.; Luo, Q.; Xie, G.; Ma, Y.; Shen, L. ICG-001 suppresses growth of gastric cancer cells and reduces chemoresistance of cancer stem cell-like population. J. Exp. Clin. Cancer Res. 2017, 36, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arensman, M.D.; Telesca, D.; Lay, A.R.; Kershaw, K.M.; Wu, N.; Donahue, T.R.; Dawson, D.W. The CREB-binding protein inhibitor ICG-001 suppresses pancreatic cancer growth. Mol. Cancer Ther. 2014, 13, 2303–2314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morata-Tarifa, C.; Jiménez, G.; García, M.A.; Entrena, J.M.; Griñán-Lisón, C.; Aguilera, M.; Picon-Ruiz, M.; Marchal, J.A. Low adherent cancer cell subpopulations are enriched in tumorigenic and metastatic epithelial-to-mesenchymal transition-induced cancer stem-like cells. Sci. Rep. 2016, 6, 18772. [Google Scholar] [CrossRef] [Green Version]
- Cave, D.D.; Hernando-Momblona, X.; Sevillano, M.; Minchiotti, G.; Lonardo, E. Nodal-Induced L1CAM/CXCR4 subpopulation sustains tumor growth and metastasis in colorectal cancer derived organoids. Theranostics 2021, 11, 5686–5699. [Google Scholar] [CrossRef]
- Managò, S.; Tramontano, C.; Cave, D.D.; Chianese, G.; Zito, G.; De Stefano, L.; Terracciano, M.; Lonardo, E.; De Luca, A.C.; Rea, I. SERS quantification of galunisertib delivery in colorectal cancer cells by plasmonic-assisted diatomite nanoparticles. Small 2021, 17, 2101711. [Google Scholar] [CrossRef]
- Munro, M.J.; Wickremesekera, S.K.; Peng, L.; Tan, S.T.; Itinteang, T. Cancer stem cells in colorectal cancer: A review. J. Clin. Pathol. 2018, 71, 110–116. [Google Scholar] [CrossRef]
- Anastas, J.N.; Moon, R.T. WNT signalling pathways as therapeutic targets in cancer. Nat. Rev. Cancer 2013, 13, 11–26. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Chen, B. Oct4 was a novel target of Wnt signaling pathway. Mol. Cell. Biochem. 2012, 362, 233–240. [Google Scholar] [CrossRef]
- Manhas, J.; Bhattacharya, A.; Agrawal, S.; Gupta, B.; Das, P.; Deo, S.; Pal, S.; Sen, S. Characterization of cancer stem cells from different grades of human colorectal cancer. Tumor Biol. 2016, 37, 14069–14081. [Google Scholar] [CrossRef]
- O’Brien, C.A.; Kreso, A.; Jamieson, C.H. Cancer stem cells and self-renewal. Clin. Cancer Res. 2010, 16, 3113–3120. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Kang, Y. Epithelial-Mesenchymal plasticity in cancer progression and metastasis. Dev. Cell 2019, 49, 361–374. [Google Scholar] [CrossRef]
- Liu, C.; Takada, K.; Zhu, D. Targeting Wnt/β-catenin pathway for drug therapy. Med. Drug Discov. 2020, 8, 100066. [Google Scholar] [CrossRef]
- Zhang, Y.; Shao, F.; Yang, C.; Yang, D.; Du, W. ICG001 inhibits colon cancer growth by suppressing eIF3D signaling. Int. J. Clin. Exp. Med. 2019, 12, 3476–3483. [Google Scholar]
- Mann, R.S.; Affolter, M. Hox proteins meet more partners. Curr. Opin. Genet. Dev. 1998, 8, 423–429. [Google Scholar] [CrossRef]
- Mohr, S.; Doebele, C.; Comoglio, F.; Berg, T.; Beck, J.; Bohnenberger, H.; Alexe, G.; Corso, J.; Ströbel, P.; Wachter, A. Hoxa9 and Meis1 cooperatively induce addiction to Syk signaling by suppressing miR-146a in acute myeloid leukemia. Cancer Cell 2017, 31, 549–562.e11. [Google Scholar] [CrossRef] [Green Version]
- Spieker, N.; van Sluis, P.; Beitsma, M.; Boon, K.; van Schaik, B.D.; van Kampen, A.H.; Caron, H.; Versteeg, R. The MEIS1 oncogene is highly expressed in neuroblastoma and amplified in cell line IMR32. Genomics 2001, 71, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Jorissen, R.N.; Gibbs, P.; Christie, M.; Prakash, S.; Lipton, L.; Desai, J.; Kerr, D.; Aaltonen, L.A.; Arango, D.; Kruhøffer, M. Metastasis-associated gene expression changes predict poor outcomes in patients with Dukes stage B and C colorectal cancer. Clin. Cancer Res. 2009, 15, 7642–7651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-Y.; Choi, J.-H.; Nam, J.-S. Targeting cancer stem cells in triple-negative breast cancer. Cancers 2019, 11, 965. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-Y.; Kim, J.-Y.; Jang, G.-B.; Choi, J.-H.; Kim, J.-H.; Lee, C.-J.; Lee, S.; Baek, J.-H.; Park, K.-K.; Kim, J.-M. Aberrant activation of the CD45-Wnt signaling axis promotes stemness and therapy resistance in colorectal cancer cells. Theranostics 2021, 11, 8755. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.-H.; Jang, T.-Y.; Jeon, S.-E.; Kim, J.-H.; Lee, C.-J.; Yun, H.-J.; Jung, J.-Y.; Park, S.-Y.; Nam, J.-S. The Small-Molecule Wnt Inhibitor ICG-001 Efficiently Inhibits Colorectal Cancer Stemness and Metastasis by Suppressing MEIS1 Expression. Int. J. Mol. Sci. 2021, 22, 13413. https://doi.org/10.3390/ijms222413413
Choi J-H, Jang T-Y, Jeon S-E, Kim J-H, Lee C-J, Yun H-J, Jung J-Y, Park S-Y, Nam J-S. The Small-Molecule Wnt Inhibitor ICG-001 Efficiently Inhibits Colorectal Cancer Stemness and Metastasis by Suppressing MEIS1 Expression. International Journal of Molecular Sciences. 2021; 22(24):13413. https://doi.org/10.3390/ijms222413413
Chicago/Turabian StyleChoi, Jang-Hyun, Tae-Young Jang, So-El Jeon, Jee-Heun Kim, Choong-Jae Lee, Hyeon-Ji Yun, Ji-Youn Jung, So-Yeon Park, and Jeong-Seok Nam. 2021. "The Small-Molecule Wnt Inhibitor ICG-001 Efficiently Inhibits Colorectal Cancer Stemness and Metastasis by Suppressing MEIS1 Expression" International Journal of Molecular Sciences 22, no. 24: 13413. https://doi.org/10.3390/ijms222413413
APA StyleChoi, J.-H., Jang, T.-Y., Jeon, S.-E., Kim, J.-H., Lee, C.-J., Yun, H.-J., Jung, J.-Y., Park, S.-Y., & Nam, J.-S. (2021). The Small-Molecule Wnt Inhibitor ICG-001 Efficiently Inhibits Colorectal Cancer Stemness and Metastasis by Suppressing MEIS1 Expression. International Journal of Molecular Sciences, 22(24), 13413. https://doi.org/10.3390/ijms222413413