
Activin A Promotes Osteoblastic Differentiation of Human Preosteoblasts through the ALK1-Smad1/5/9 Pathway

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Expression of Activin A in Rat PDL Tissue and Saos2 Cells
2.2. Effects of Activin A on Osteoblastic Differentiation of Saos2 Cells
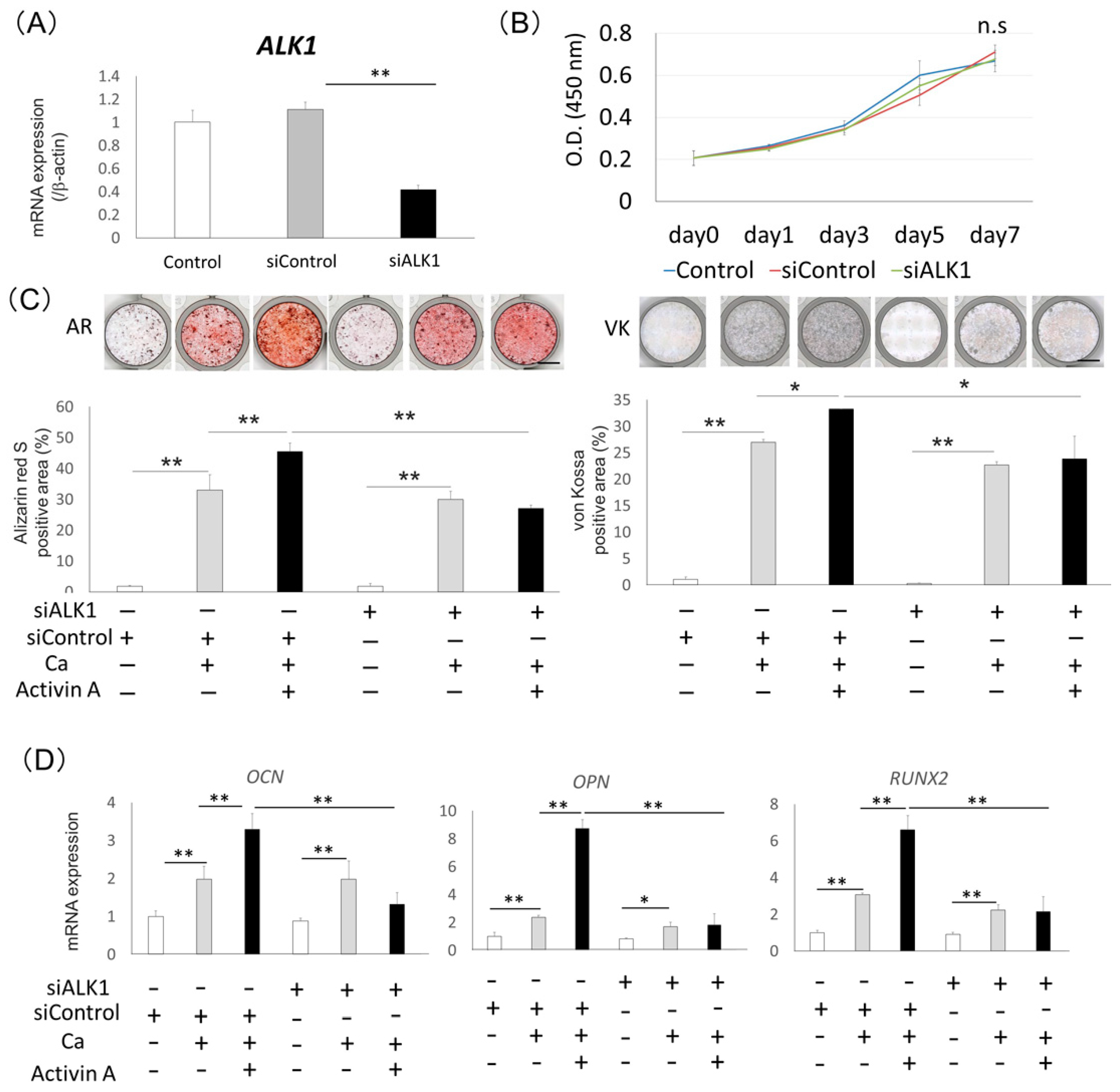
2.3. Effects of Activin Type I Receptors on Activin A-Induced Osteoblastic Differentiation in Saos2 Cells
2.4. Expression of ALK1 in Rat PDL Tissue
2.5. Expression of ALK1 in Surgically Damaged PDL Tissue
2.6. Effects of ALK1 Knockdown on Phosphorylation of Activin A-Related Intracellular Signaling Molecules in Saos2 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Animal Model
4.3. Western Blotting Analysis
4.4. Immunofluorescence and Immunochemical Analyses
4.5. Treatment of Saos2 Cells with IL-1β or TNF-α
4.6. Quantitative Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.7. Osteoblastic Differentiation of Saos2 Cells
4.8. Small Interfering RNA Transfection
4.9. Proliferation Assay
4.10. ALP Activity Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Beertsen, W.; McCulloch, C.A.; Sodek, J. The periodontal ligament: A unique, multifunctional connective tissue. Periodontology 2000 1997, 13, 20–40. [Google Scholar] [CrossRef]
- Moxham, B.J.; Berkovitz, B.K. Continuous monitoring of the movements of erupting and newly erupted teeth of limited growth (ferret mandibular canines) and their responses to hexamethonium. Arch. Oral. Biol. 1988, 33, 919–923. [Google Scholar] [CrossRef]
- Bartold, P.M.; McCulloch, C.A.; Narayanan, A.S.; Pitaru, S. Tissue engineering: A new paradigm for periodontal regeneration based on molecular and cell biology. Periodontology 2000 2000, 24, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Shimono, M.; Ishikawa, T.; Ishikawa, H.; Matsuzaki, H.; Hashimoto, S.; Muramatsu, T.; Shima, K.; Matsuzaka, K.; Inoue, T. Regulatory mechanisms of periodontal regeneration. Microsc. Res. Tech. 2003, 60, 491–502. [Google Scholar] [CrossRef]
- Nandini, S.G.; Lipner, H.; Moudgal, N.R. A model system for studying inhibin. Endocrinology 1976, 98, 1460–1465. [Google Scholar] [CrossRef]
- Bernard, D.J.; Tran, S. Mechanisms of activin-stimulated FSH synthesis: The story of a pig and a FOX. Biol. Reprod. 2013, 88, 78. [Google Scholar] [CrossRef]
- Bloise, E.; Ciarmela, P.; Dela Cruz, C.; Luisi, S.; Petraglia, F.; Reis, F.M. Activin A in Mammalian Physiology. Physiol. Rev. 2019, 99, 739–780. [Google Scholar] [CrossRef]
- Wu, H.; Chen, Y.; Winnall, W.R.; Phillips, D.J.; Hedger, M.P. Acute regulation of activin A and its binding protein, follistatin, in serum and tissues following lipopolysaccharide treatment of adult male mice. Am. J. Physiol Regul Integr Comp. Physiol. 2012, 303, R665–R675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, O.; Nau, R.; Michel, U. Comparative analysis of follistatin-, activin beta A- and activin beta B-mRNA steady-state levels in diverse porcine tissues by multiplex S1 nuclease analysis. Eur. J. Endocrinol. 2000, 142, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Meunier, H.; Rivier, C.; Evans, R.M.; Vale, W. Gonadal and extragonadal expression of inhibin alpha, beta A, and beta B subunits in various tissues predicts diverse functions. Proc. Natl. Acad. Sci. USA 1988, 85, 247–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicks, K.M.; Perrien, D.S.; Akel, N.S.; Suva, L.J.; Gaddy, D. Regulation of osteoblastogenesis and osteoclastogenesis by the other reproductive hormones, Activin and Inhibin. Mol. Cell Endocrinol. 2009, 310, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Zaragosi, L.E.; Wdziekonski, B.; Villageois, P.; Keophiphath, M.; Maumus, M.; Tchkonia, T.; Bourlier, V.; Mohsen-Kanson, T.; Ladoux, A.; Elabd, C.; et al. Activin a plays a critical role in proliferation and differentiation of human adipose progenitors. Diabetes 2010, 59, 2513–2521. [Google Scholar] [CrossRef] [Green Version]
- Gokoffski, K.K.; Wu, H.H.; Beites, C.L.; Kim, J.; Kim, E.J.; Matzuk, M.M.; Johnson, J.E.; Lander, A.D.; Calof, A.L. Activin and GDF11 collaborate in feedback control of neuroepithelial stem cell proliferation and fate. Development 2011, 138, 4131–4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholls, P.K.; Stanton, P.G.; Chen, J.L.; Olcorn, J.S.; Haverfield, J.T.; Qian, H.; Walton, K.L.; Gregorevic, P.; Harrison, C.A. Activin signaling regulates Sertoli cell differentiation and function. Endocrinology 2012, 153, 6065–6077. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Furue, M.; Nagamine, K.; Abe, T.; Fukui, Y.; Myoishi, Y.; Fujii, T.; Okamoto, T.; Taketani, Y.; Asashima, M. Modulation of activin A-induced differentiation in vitro by vascular endothelial growth factor in Xenopus presumptive ectodermal cells. Vitr. Cell Dev. Biol. Anim. 2005, 41, 104–110. [Google Scholar] [CrossRef]
- Salogni, L.; Musso, T.; Bosisio, D.; Mirolo, M.; Jala, V.R.; Haribabu, B.; Locati, M.; Sozzani, S. Activin A induces dendritic cell migration through the polarized release of CXC chemokine ligands 12 and 14. Blood 2009, 113, 5848–5856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neel, J.C.; Lebrun, J.J. Activin and TGFbeta regulate expression of the microRNA-181 family to promote cell migration and invasion in breast cancer cells. Cell Signal. 2013, 25, 1556–1566. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.G.; Lui, H.M.; Lin, S.L.; Lee, J.M.; Ying, S.Y. Regulation of cell proliferation, apoptosis, and carcinogenesis by activin. Exp. Biol. Med. 2002, 227, 75–87. [Google Scholar] [CrossRef]
- Heldin, C.H.; Moustakas, A. Signaling Receptors for TGF-beta Family Members. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Svoboda, K.K.; Feng, J.Q.; Jiang, X. The biological function of type I receptors of bone morphogenetic protein in bone. Bone Res. 2016, 4, 16005. [Google Scholar] [CrossRef] [Green Version]
- Koenig, B.B.; Cook, J.S.; Wolsing, D.H.; Ting, J.; Tiesman, J.P.; Correa, P.E.; Olson, C.A.; Pecquet, A.L.; Ventura, F.; Grant, R.A.; et al. Characterization and cloning of a receptor for BMP-2 and BMP-4 from NIH 3T3 cells. Mol. Cell Biol. 1994, 14, 5961–5974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmierer, B.; Hill, C.S. TGFbeta-SMAD signal transduction: Molecular specificity and functional flexibility. Nat. Rev. Mol. Cell Biol. 2007, 8, 970–982. [Google Scholar] [CrossRef]
- Zhou, Y.; Sun, H.; Danila, D.C.; Johnson, S.R.; Sigai, D.P.; Zhang, X.; Klibanski, A. Truncated activin type I receptor Alk4 isoforms are dominant negative receptors inhibiting activin signaling. Mol. Endocrinol. 2000, 14, 2066–2075. [Google Scholar] [CrossRef]
- Suzuki, K.; Kobayashi, T.; Funatsu, O.; Morita, A.; Ikekita, M. Activin A induces neuronal differentiation and survival via ALK4 in a SMAD-independent manner in a subpopulation of human neuroblastomas. Biochem. Biophys. Res. Commun. 2010, 394, 639–645. [Google Scholar] [CrossRef]
- Liu, C.; Chang, H.M.; Yi, Y.; Fang, Y.; Zhao, F.; Leung, P.C.K.; Yang, X. ALK4-SMAD2/3-SMAD4 signaling mediates the activin A-induced suppression of PTX3 in human granulosa-lutein cells. Mol. Cell Endocrinol. 2019, 493, 110485. [Google Scholar] [CrossRef] [PubMed]
- Olsen, O.E.; Hella, H.; Elsaadi, S.; Jacobi, C.; Martinez-Hackert, E.; Holien, T. Activins as Dual Specificity TGF-beta Family Molecules: SMAD-Activation via Activin- and BMP-Type 1 Receptors. Biomolecules 2020, 10, 519. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Jiang, W.; Lacroix, J.J.; Luo, Y.; Hao, J. Insight into Molecular Mechanism for Activin A-Induced Bone Morphogenetic Protein Signaling. Int. J. Mol. Sci. 2020, 21, 6498. [Google Scholar] [CrossRef] [PubMed]
- ten Dijke, P.; Yamashita, H.; Ichijo, H.; Franzen, P.; Laiho, M.; Miyazono, K.; Heldin, C.H. Characterization of type I receptors for transforming growth factor-beta and activin. Science 1994, 264, 101–104. [Google Scholar] [CrossRef]
- Zhu, D.; Mackenzie, N.C.; Shanahan, C.M.; Shroff, R.C.; Farquharson, C.; MacRae, V.E. BMP-9 regulates the osteoblastic differentiation and calcification of vascular smooth muscle cells through an ALK1 mediated pathway. J. Cell Mol. Med. 2015, 19, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Takeda, K.; Imamura, T.; Aoki, H.; Sampath, T.K.; Enomoto, S.; Kawabata, M.; Kato, M.; Ichijo, H.; Miyazono, K. Roles of bone morphogenetic protein type I receptors and Smad proteins in osteoblast and chondroblast differentiation. Mol. Biol. Cell 1999, 10, 3801–3813. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Tang, M.; Huang, J.; He, B.C.; Gao, J.L.; Chen, L.; Zuo, G.W.; Zhang, W.; Luo, Q.; Shi, Q.; et al. TGFbeta/BMP type I receptors ALK1 and ALK2 are essential for BMP9-induced osteogenic signaling in mesenchymal stem cells. J. Biol. Chem. 2010, 285, 29588–29598. [Google Scholar] [CrossRef] [Green Version]
- Song, T.; Wang, W.; Xu, J.; Zhao, D.; Dong, Q.; Li, L.; Yang, X.; Duan, X.; Liang, Y.; Xiao, Y.; et al. Fibroblast growth factor 2 inhibits bone morphogenetic protein 9-induced osteogenic differentiation of mesenchymal stem cells by repressing Smads signaling and subsequently reducing Smads dependent up-regulation of ALK1 and ALK2. Int. J. Biochem. Cell Biol. 2013, 45, 1639–1646. [Google Scholar] [CrossRef]
- Wang, X.P.; Suomalainen, M.; Jorgez, C.J.; Matzuk, M.M.; Wankell, M.; Werner, S.; Thesleff, I. Modulation of activin/bone morphogenetic protein signaling by follistatin is required for the morphogenesis of mouse molar teeth. Dev. Dyn. 2004, 231, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, C.A.; Tucker, A.S.; Christensen, L.; Lau, A.L.; Matzuk, M.M.; Sharpe, P.T. Activin is an essential early mesenchymal signal in tooth development that is required for patterning of the murine dentition. Genes Dev. 1998, 12, 2636–2649. [Google Scholar] [CrossRef] [Green Version]
- Maas, R.; Bei, M. The genetic control of early tooth development. Crit. Rev. Oral Biol. Med. 1997, 8, 4–39. [Google Scholar] [CrossRef] [PubMed]
- Sugii, H.; Maeda, H.; Tomokiyo, A.; Yamamoto, N.; Wada, N.; Koori, K.; Hasegawa, D.; Hamano, S.; Yuda, A.; Monnouchi, S.; et al. Effects of Activin A on the phenotypic properties of human periodontal ligament cells. Bone 2014, 66, 62–71. [Google Scholar] [CrossRef]
- Ogawa, Y.; Schmidt, D.K.; Nathan, R.M.; Armstrong, R.M.; Miller, K.L.; Sawamura, S.J.; Ziman, J.M.; Erickson, K.L.; de Leon, E.R.; Rosen, D.M.; et al. Bovine bone activin enhances bone morphogenetic protein-induced ectopic bone formation. J. Biol. Chem. 1992, 267, 14233–14237. [Google Scholar] [CrossRef]
- Gaddy-Kurten, D.; Coker, J.K.; Abe, E.; Jilka, R.L.; Manolagas, S.C. Inhibin suppresses and activin stimulates osteoblastogenesis and osteoclastogenesis in murine bone marrow cultures. Endocrinology 2002, 143, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Oue, Y.; Kanatani, H.; Kiyoki, M.; Eto, Y.; Ogata, E.; Matsumoto, T. Effect of local injection of activin A on bone formation in newborn rats. Bone 1994, 15, 361–366. [Google Scholar] [CrossRef]
- Hirotani, H.; Ohtsuka-Isoya, M.; Mori, S.; Sakai, R.; Eto, Y.; Echigo, S.; Shinoda, H. Activin A increases the bone mass of grafted bone in C3H/HeJ mice. Calcif. Tissue Int. 2002, 70, 330–338. [Google Scholar] [CrossRef]
- Camargos, B.M.; Tavares, R.L.; Del Puerto, H.L.; Andrade, L.O.; Camargos, A.F.; Reis, F.M. BMP-4 increases activin A gene expression during osteogenic differentiation of mouse embryonic stem cells. Growth Factors 2015, 33, 133–138. [Google Scholar] [CrossRef]
- Ebner, J.K.; Konig, G.M.; Kostenis, E.; Siegert, P.; Aktories, K.; Orth, J.H.C. Activation of Gq signaling by Pasteurella multocida toxin inhibits the osteoblastogenic-like actions of Activin A in C2C12 myoblasts, a cell model of fibrodysplasia ossificans progressiva. Bone 2019, 127, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Shibata, H.; Yasuda, H.; Sekine, N.; Mine, T.; Totsuka, Y.; Kojima, I. Activin A increases intracellular free calcium concentrations in rat pancreatic islets. FEBS Lett. 1993, 329, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, N.; Kamiya, N.; Suzuki, N.; Matsumoto, M.; Takagi, M. Changes in extracellular activin A:follistatin ratio during differentiation of a mesenchymal progenitor cell line, ROB-C26 into osteoblasts and adipocytes. Life Sci. 2007, 81, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Ikenoue, T.; Jingushi, S.; Urabe, K.; Okazaki, K.; Iwamoto, Y. Inhibitory effects of activin-A on osteoblast differentiation during cultures of fetal rat calvarial cells. J. Cell Biochem. 1999, 75, 206–214. [Google Scholar] [CrossRef]
- Alves, R.D.; Eijken, M.; Bezstarosti, K.; Demmers, J.A.; van Leeuwen, J.P. Activin A suppresses osteoblast mineralization capacity by altering extracellular matrix (ECM) composition and impairing matrix vesicle (MV) production. Mol. Cell Proteom. 2013, 12, 2890–2900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jong, D.S.; van Zoelen, E.J.; Bauerschmidt, S.; Olijve, W.; Steegenga, W.T. Microarray analysis of bone morphogenetic protein, transforming growth factor beta, and activin early response genes during osteoblastic cell differentiation. J. Bone Min. Res. 2002, 17, 2119–2129. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, F.; Wu, Q.; Liu, X. Activin A regulates proliferation, invasion and migration in osteosarcoma cells. Mol. Med. Rep. 2015, 11, 4501–4507. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Kawaguchi, Y.; Kawamoto, M.; Ota, Y.; Tochimoto, A.; Gono, T.; Katsumata, Y.; Takagi, M.; Hara, M.; Yamanaka, H. Activation of the activin A-ALK-Smad pathway in systemic sclerosis. J. Autoimmun. 2011, 36, 181–188. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, Y.; Zhang, P.; Chen, Y.; Li, C.; Chen, J.; Wang, Y.; Li, Y. The crucial role of activin A/ALK4 pathway in the pathogenesis of Ang-II-induced atrial fibrosis and vulnerability to atrial fibrillation. Basic Res. Cardiol. 2017, 112, 47. [Google Scholar] [CrossRef] [PubMed]
- Cunha, S.I.; Pietras, K. ALK1 as an emerging target for antiangiogenic therapy of cancer. Blood 2011, 117, 6999–7006. [Google Scholar] [CrossRef] [Green Version]
- de Kroon, L.M.; Narcisi, R.; Blaney Davidson, E.N.; Cleary, M.A.; van Beuningen, H.M.; Koevoet, W.J.; van Osch, G.J.; van der Kraan, P.M. Activin Receptor-Like Kinase Receptors ALK5 and ALK1 Are Both Required for TGFbeta-Induced Chondrogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells. PLoS ONE 2015, 10, e0146124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valcourt, U.; Gouttenoire, J.; Moustakas, A.; Herbage, D.; Mallein-Gerin, F. Functions of transforming growth factor-beta family type I receptors and Smad proteins in the hypertrophic maturation and osteoblastic differentiation of chondrocytes. J. Biol. Chem. 2002, 277, 33545–33558. [Google Scholar] [CrossRef] [Green Version]
- Kantarci, A.; Van Dyke, T.E. Resolution of inflammation in periodontitis. J. Periodontol. 2005, 76, 2168–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pihlstrom, B.L.; Michalowicz, B.S.; Johnson, N.W. Periodontal diseases. Lancet 2005, 366, 1809–1820. [Google Scholar] [CrossRef] [Green Version]
- Doshi, B.M.; Perdrizet, G.A.; Hightower, L.E. Wound healing from a cellular stress response perspective. Cell Stress Chaperones 2008, 13, 393–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, S.; Doi, Y.; Kudo, H.; Furukawa, H.; Nakamura, T.; Fujimoto, S. Transient expression of activin betaA mRNA on osteoprogenitor cells in rat bone regeneration after drill-hole injury. Bone 2000, 27, 81–90. [Google Scholar] [CrossRef]
- Konig, H.G.; Kogel, D.; Rami, A.; Prehn, J.H. TGF-{beta}1 activates two distinct type I receptors in neurons: Implications for neuronal NF-{kappa}B signaling. J. Cell Biol. 2005, 168, 1077–1086. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, N.; Maeda, H.; Tomokiyo, A.; Fujii, S.; Wada, N.; Monnouchi, S.; Kono, K.; Koori, K.; Teramatsu, Y.; Akamine, A. Expression and effects of glial cell line-derived neurotrophic factor on periodontal ligament cells. J. Clin. Periodontol. 2012, 39, 556–564. [Google Scholar] [CrossRef]
- Yoshino, O.; Izumi, G.; Shi, J.; Osuga, Y.; Hirota, Y.; Hirata, T.; Harada, M.; Nishii, O.; Koga, K.; Taketani, Y. Activin-A is induced by interleukin-1beta and tumor necrosis factor-alpha and enhances the mRNA expression of interleukin-6 and protease-activated receptor-2 and proliferation of stromal cells from endometrioma. Fertil. Steril. 2011, 96, 118–121. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene (Abbreviation) | Forward (Top) and Reverse (Bottom) Primer Sequences | Size of Amplified Products (bp) | Annealing Temperature (°C) | Sequence ID |
|---|---|---|---|---|
| β-actin | 5’-ATTGCCGACAGGATGCAGA-3’ 5’-GAGTACTTGCGCTCAGGAGGA-3’ | 89 | 60 | NM_001101.3 |
| ALK1 | 5’-ATCCTAGGCTTCATCGCCTC-3’ 5’-TAGCCTCAGAGCCAGATGGG-3’ | 141 | 60 | NM_000020.3 |
| ALK2 | 5’-AGGCTGCTTCCAGGTTTATGAG-3’ 5’-TGGCAGCACTCCACAGCTT-3’ | 81 | 60 | NM_001105.5 |
| ALK3 | 5’-AGTGTCTCCAGTCAAGCTCTGGGTA-3’ 5’-CCATCTCTGCTGCGCTCATTTA-3’ | 97 | 60 | NM_004329.3 |
| ALK4 | 5’-TGCTGCGCCATGAAAACATC-3’ 5’-GGGACCCGTGCTCATGATAG-3’ | 105 | 60 | NM_020327.3 |
| ALK5 | 5’-CCTTCTGACCCATCAGTTGAAGA-3’ 5’-CCTAGCTGCTCCATTGGCAT-3’ | 150 | 60 | NM_004612.4 |
| ALK6 | 5’-GTTACGCCCCTCATTCCCAA-3’ 5’-GTTTTCTTAACCCGCAGGGC-3’ | 126 | 60 | NM_001256793.2 |
| Inhibinβa | 5’-TGTGCCCACCAAGCTGAGAC-3’ 5’-CTGGGCTGGGCAACTCTATGA-3’ | 124 | 60 | NM_002192.2 |
| OCN | 5’-GTGCAGAGTCCAGCAAAGGT-3’ 5’-TCAGCCAACTCGTCACAGTC-3’ | 110 | 60 | NM_199173.4 |
| OPN | 5’-ACACATATGATGGCCGAGGTGA-3’ 5’-TGTGAGGTGATGTCCTCGTCTGT-3’ | 115 | 60 | NM_000582.3 |
| RUNX2 | 5’-AACCCTTAATTTGCACTGGGTCA-3’ 5’-CAAATTCCAGCAATGTTTGTGCTAC-3’ | 145 | 60 | NM_001024630.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugii, H.; Albougha, M.S.; Adachi, O.; Tomita, H.; Tomokiyo, A.; Hamano, S.; Hasegawa, D.; Yoshida, S.; Itoyama, T.; Maeda, H. Activin A Promotes Osteoblastic Differentiation of Human Preosteoblasts through the ALK1-Smad1/5/9 Pathway. Int. J. Mol. Sci. 2021, 22, 13491. https://doi.org/10.3390/ijms222413491
Sugii H, Albougha MS, Adachi O, Tomita H, Tomokiyo A, Hamano S, Hasegawa D, Yoshida S, Itoyama T, Maeda H. Activin A Promotes Osteoblastic Differentiation of Human Preosteoblasts through the ALK1-Smad1/5/9 Pathway. International Journal of Molecular Sciences. 2021; 22(24):13491. https://doi.org/10.3390/ijms222413491
Chicago/Turabian StyleSugii, Hideki, Mhd Safwan Albougha, Orie Adachi, Hiroka Tomita, Atsushi Tomokiyo, Sayuri Hamano, Daigaku Hasegawa, Shinichiro Yoshida, Tomohiro Itoyama, and Hidefumi Maeda. 2021. "Activin A Promotes Osteoblastic Differentiation of Human Preosteoblasts through the ALK1-Smad1/5/9 Pathway" International Journal of Molecular Sciences 22, no. 24: 13491. https://doi.org/10.3390/ijms222413491