
Brassinazole Resistant 1 Activity Is Organ-Specific and Genotype-Dependent in Barley Seedlings


Abstract
1. Introduction
2. Results
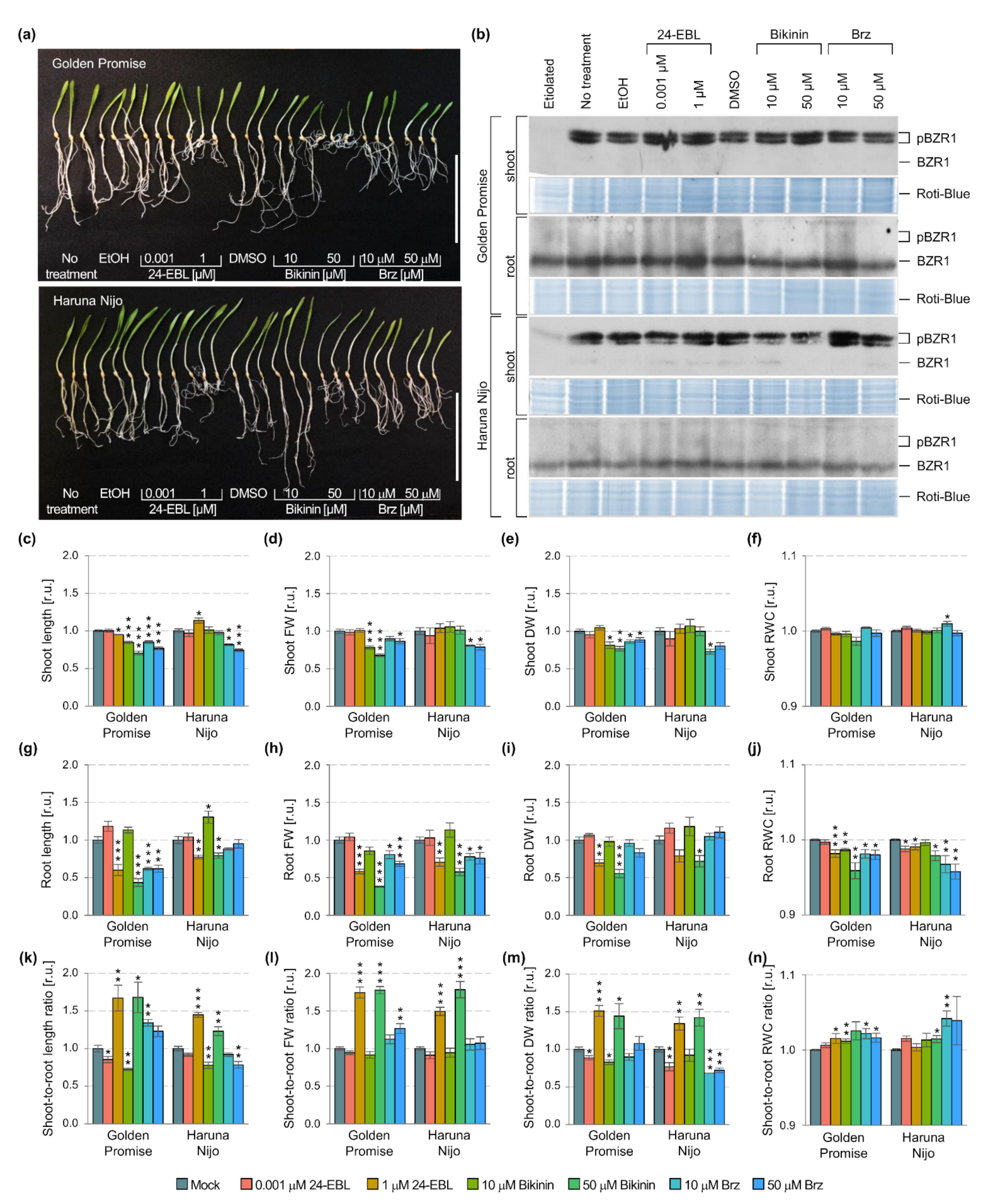
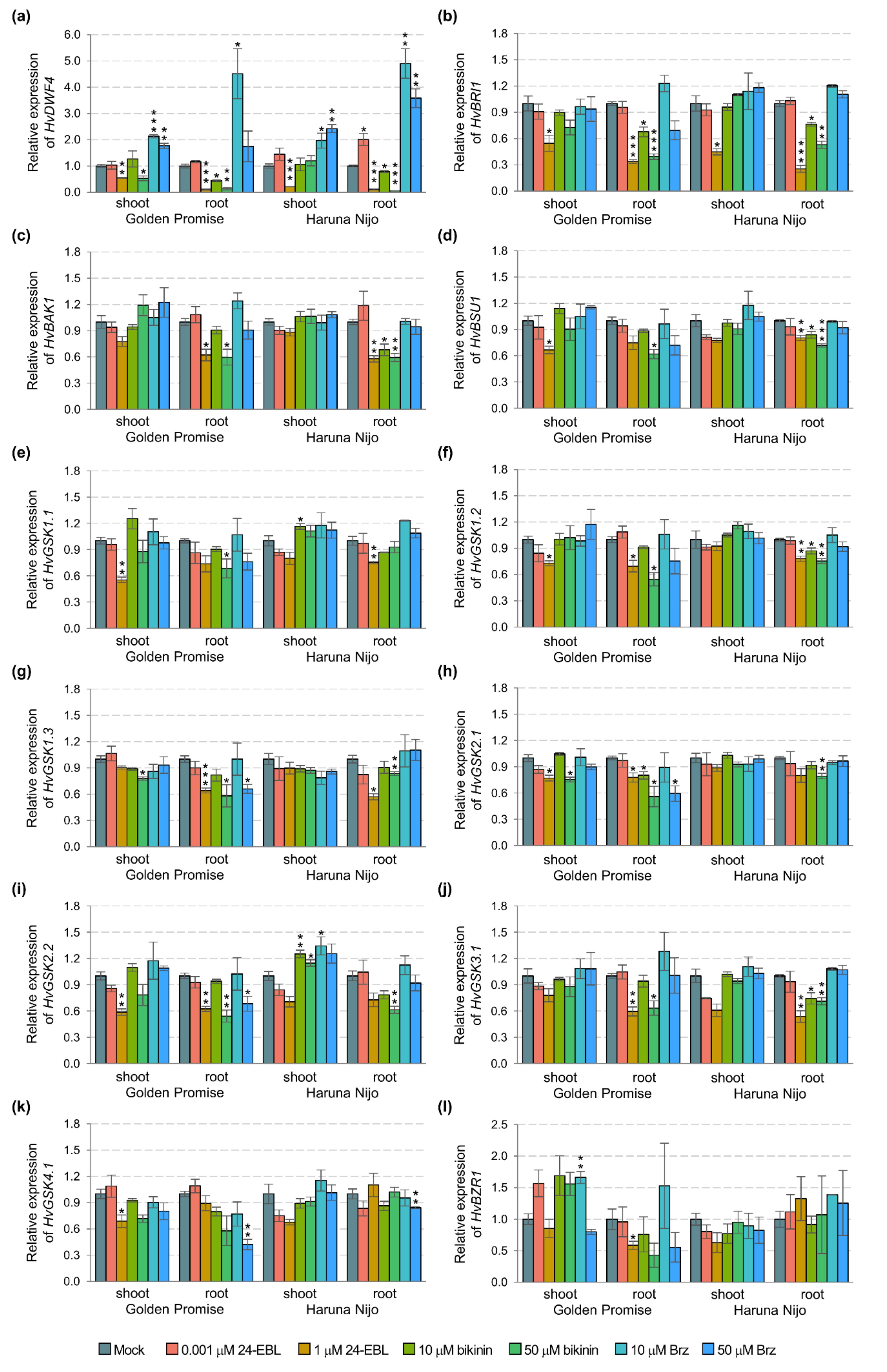
2.1. HvBZR1 Activity, Plant Phenotype, and BR-Related Gene Expression
2.2. Barley Response after 24-EBL, Bikinin, and Brz Treatment
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.3. Treatments and Growth Traits
4.4. Western Blot Detection of GSK2.1 and BZR1 Proteins
4.5. RNA Extraction, cDNA Synthesis, and Real-Time PCR Analysis
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bajguz, A. Metabolism of brassinosteroids in plants. Plant Physiol. Biochem. 2007, 45, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Goda, H.; Nakamura, A.; Takatsuto, S.; Fujioka, S.; Yoshida, S. Organ-specific expression of brassinosteroid-biosynthetic genes and distribution of endogenous brassinosteroids in Arabidopsis. Plant Physiol. 2003, 131, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bai, M.-Y.; Chong, K. Brassinosteroid-mediated regulation of agronomic traits in rice. Plant Cell Rep. 2014, 33, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Wu, C.; Wang, C.; Roh, J.; Zhang, L.; Chen, J.; Zhang, S.; Zhang, H.; Yang, C.; Hu, J. SLG controls grain size and leaf angle by modulating brassinosteroid homeostasis in rice. J. Exp. Bot. 2016, 67, 4241–4253. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, G.; Liu, D.; Niu, M.; Tong, H.; Chu, C. GSK2 stabilizes OFP3 to suppress brassinosteroid responses in rice. Plant J. 2020, 102, 1187–1201. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Bai, M.-Y.; Oh, E.; Zhu, J.-Y. Brassinosteroid signaling network and regulation of photomorphogenesis. Annu. Rev. Genet. 2012, 46, 701–724. [Google Scholar] [CrossRef]
- Duan, P.; Ni, S.; Wang, J.; Zhang, B.; Xu, R.; Wang, Y.; Chen, H.; Zhu, X.; Li, Y. Regulation of OsGRF4 by OsmiR396 controls grain size and yield in rice. Nat. Plants 2015, 2, 15203. [Google Scholar] [CrossRef]
- Duan, P.; Rao, Y.; Zeng, D.; Yang, Y.; Xu, R.; Zhang, B.; Dong, G.; Qian, Q.; Li, Y. SMALL GRAIN 1, which encodes a mitogen-activated protein kinase kinase 4, influences grain size in rice. Plant J. 2014, 77, 547–557. [Google Scholar] [CrossRef]
- Li, Y.; Fan, C.; Xing, Y.; Jiang, Y.; Luo, L.; Sun, L.; Shao, D.; Xu, C.; Li, X.; Xiao, J. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat. Genet. 2011, 43, 1266. [Google Scholar] [CrossRef]
- Wang, S.; Wu, K.; Yuan, Q.; Liu, X.; Liu, Z.; Lin, X.; Zeng, R.; Zhu, H.; Dong, G.; Qian, Q. Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 2012, 44, 950. [Google Scholar] [CrossRef]
- Nakagawa, H.; Tanaka, A.; Tanabata, T.; Ohtake, M.; Fujioka, S.; Nakamura, H.; Ichikawa, H.; Mori, M. Short grain1 decreases organ elongation and brassinosteroid response in rice. Plant Physiol. 2012, 158, 1208–1219. [Google Scholar] [CrossRef]
- Corvalan, C.; An, G.; Choe, S. The Rice propiconazole resistant 1-D mutant, with activated expression of a DPb transcription factor gene, exhibits increased seed yields. bioRxiv 2021. [Google Scholar] [CrossRef]
- Bittner, T.; Campagne, S.; Neuhaus, G.; Rensing, S.A.; Fischer-Iglesias, C. Identification and characterization of two wheat Glycogen Synthase Kinase 3/SHAGGY-like kinases. BMC Plant Biol. 2013, 13, 64. [Google Scholar] [CrossRef]
- Hartwig, T.; Corvalan, C.; Best, N.B.; Budka, J.S.; Zhu, J.Y.; Choe, S.; Schulz, B. Propiconazole is a specific and accessible brassinosteroid (BR) biosynthesis inhibitor for Arabidopsis and maize. PLoS ONE 2012, 7, e36625. [Google Scholar] [CrossRef]
- Gruszka, D. Exploring the Brassinosteroid Signaling in Monocots Reveals Novel Components of the Pathway and Implications for Plant Breeding. Int. J. Mol. Sci. 2020, 21, 354. [Google Scholar] [CrossRef]
- Cui, X.-Y.; Gao, Y.; Guo, J.; Yu, T.-F.; Zheng, W.-J.; Liu, Y.-W.; Chen, J.; Xu, Z.-S.; Ma, Y.-Z. BES/BZR Transcription Factor TaBZR2 Positively Regulates Drought Responses by Activation of TaGST1. Plant Physiol. 2019, 180, 605–620. [Google Scholar] [CrossRef]
- Corvalan, C.; Choe, S. Identification of brassinosteroid genes in Brachypodium distachyon. BMC Plant Biol. 2017, 17, 5. [Google Scholar] [CrossRef]
- Groszyk, J.; Szechyńska-Hebda, M. Effects of 24-Epibrassinolide, Bikinin, and Brassinazole on Barley Growth under Salinity Stress Are Genotype- and Dose-Dependent. Agronomy 2021, 11, 259. [Google Scholar] [CrossRef]
- Castorina, G.; Consonni, G. The Role of Brassinosteroids in Controlling Plant Height in Poaceae: A Genetic Perspective. Int. J. Mol. Sci. 2020, 21, 1191. [Google Scholar] [CrossRef]
- Nolan, T.M.; Vukašinović, N.; Liu, D.; Russinova, E.; Yin, Y. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2020, 32, 295–318. [Google Scholar] [CrossRef]
- Oh, M.-H.; Honey, S.H.; Tax, F.E. The Control of Cell Expansion, Cell Division, and Vascular Development by Brassinosteroids: A Historical Perspective. Int. J. Mol. Sci. 2020, 21, 1743. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Li, J. Regulation of Three Key Kinases of Brassinosteroid Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 4340. [Google Scholar] [CrossRef] [PubMed]
- He, J.-X.; Gendron, J.M.; Sun, Y.; Gampala, S.S.; Gendron, N.; Sun, C.Q.; Wang, Z.-Y. BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses. Science 2005, 307, 1634–1638. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Vafeados, D.; Tao, Y.; Yoshida, S.; Asami, T.; Chory, J. A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell 2005, 120, 249–259. [Google Scholar] [CrossRef]
- Goda, H.; Shimada, Y.; Asami, T.; Fujioka, S.; Yoshida, S. Microarray analysis of brassinosteroid-regulated genes in Arabidopsis. Plant Physiol. 2002, 130, 1319–1334. [Google Scholar] [CrossRef]
- Bai, M.Y.; Zhang, L.Y.; Gampala, S.S.; Zhu, S.W.; Song, W.Y.; Chong, K.; Wang, Z.Y. Functions of OsBZR1 and 14-3-3 proteins in brassinosteroid signaling in rice. Proc. Natl. Acad. Sci. USA 2007, 104, 13839–13844. [Google Scholar] [CrossRef]
- He, Y.; Hong, G.; Zhang, H.; Tan, X.; Li, L.; Kong, Y.; Sang, T.; Xie, K.; Wei, J.; Li, J. The OsGSK2 kinase integrates brassinosteroid and jasmonic acid signaling by interacting with OsJAZ4. Plant Cell 2020, 32, 2806–2822. [Google Scholar] [CrossRef]
- Hughes, P.W. OsGSK2 Integrates Jasmonic Acid and Brassinosteroid Signaling in Rice. Am. Soc. Plant. Biol. 2020, 32, 2669–2670. [Google Scholar] [CrossRef]
- Xiao, Y.; Liu, D.; Zhang, G.; Tong, H.; Chu, C. Brassinosteroids regulate OFP1, a DLT interacting protein, to modulate plant architecture and grain morphology in rice. Front. Plant Sci. 2017, 8, 1698. [Google Scholar] [CrossRef]
- Tong, H.; Jin, Y.; Liu, W.; Li, F.; Fang, J.; Yin, Y.; Qian, Q.; Zhu, L.; Chu, C. DWARF AND LOW-TILLERING, a new member of the GRAS family, plays positive roles in brassinosteroid signaling in rice. Plant J. 2009, 58, 803–816. [Google Scholar] [CrossRef]
- Tong, H.; Liu, L.; Jin, Y.; Du, L.; Yin, Y.; Qian, Q.; Zhu, L.; Chu, C. DWARF AND LOW-TILLERING acts as a direct downstream target of a GSK3/SHAGGY-like kinase to mediate brassinosteroid responses in rice. Plant Cell 2012, 24, 2562–2577. [Google Scholar] [CrossRef]
- Tong, H.; Chu, C. Roles of DLT in fine modulation on brassinosteroid response in rice. Plant Signal. Behav. 2014, 4, 438–439. [Google Scholar] [CrossRef][Green Version]
- Zhang, C.; Xu, Y.; Guo, S.; Zhu, J.; Huan, Q.; Liu, H.; Wang, L.; Luo, G.; Wang, X.; Chong, K. Dynamics of brassinosteroid response modulated by negative regulator LIC in rice. PLoS Genet. 2012, 8, e1002686. [Google Scholar] [CrossRef]
- Qiao, S.; Sun, S.; Wang, L.; Wu, Z.; Li, C.; Li, X.; Wang, T.; Leng, L.; Tian, W.; Lu, T. The RLA1/SMOS1 transcription factor functions with OsBZR1 to regulate brassinosteroid signaling and rice architecture. Plant Cell 2017, 29, 292–309. [Google Scholar] [CrossRef]
- Gruszka, D.; Janeczko, A.; Dziurka, M.; Pociecha, E.; Oklestkova, J.; Szarejko, I. Barley Brassinosteroid Mutants Provide an Insight into Phytohormonal Homeostasis in Plant Reaction to Drought Stress. Front Plant Sci. 2016, 7, 1824. [Google Scholar] [CrossRef]
- Oh, E.; Zhu, J.Y.; Bai, M.Y.; Arenhart, R.A.; Sun, Y.; Wang, Z.Y. Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. eLife 2014, 3, e03031. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Z.; Xu, Y.; Joo, S.H.; Kim, S.K.; Xue, Z.; Xu, Z.; Wang, Z.; Chong, K. OsGSR1 is involved in crosstalk between gibberellins and brassinosteroids in rice. Plant J. 2009, 57, 498–510. [Google Scholar] [CrossRef]
- Yang, X.; Bai, Y.; Shang, J.; Xin, R.; Tang, W. The antagonistic regulation of abscisic acid-inhibited root growth by brassinosteroids is partially mediated via direct suppression of ABSCISIC ACID INSENSITIVE 5 expression by BRASSINAZOLE RESISTANT 1. Plant Cell Environ. 2016, 39, 1994–2003. [Google Scholar] [CrossRef]
- Sahni, S.; Prasad, B.D.; Liu, Q.; Grbic, V.; Sharpe, A.; Singh, S.P.; Krishna, P. Overexpression of the brassinosteroid biosynthetic gene DWF4 in Brassica napus simultaneously increases seed yield and stress tolerance. Sci. Rep. 2016, 6, 28298. [Google Scholar] [CrossRef]
- Kim, T.W.; Guan, S.; Sun, Y.; Deng, Z.; Tang, W.; Shang, J.X.; Sun, Y.; Burlingame, A.L.; Wang, Z.Y. Brassinosteroid signal transduction from cell-surface receptor kinases to nuclear transcription factors. Nat. Cell Biol. 2009, 11, 1254–1260. [Google Scholar] [CrossRef]
- Wang, Z.-Y.; Nakano, T.; Gendron, J.; He, J.; Chen, M.; Vafeados, D.; Yang, Y.; Fujioka, S.; Yoshida, S.; Asami, T. Nuclear-localized BZR1 mediates brassinosteroid-induced growth and feedback suppression of brassinosteroid biosynthesis. Dev. Cell 2002, 2, 505–513. [Google Scholar] [CrossRef]
- Gruszka, D.; Szarejko, I.; Maluszynski, M. New allele of HvBRI1 gene encoding brassinosteroid receptor in barley. J. Appl. Genet. 2011, 52, 257–268. [Google Scholar] [CrossRef]
- Gruszka, D.; Szarejko, I.; Maluszynski, M. Identification of barley DWARF gene involved in brassinosteroid synthesis. Plant Growth Regul. 2011, 65, 343–358. [Google Scholar] [CrossRef]
- Saidi, Y.; Hearn, T.J.; Coates, J.C. Function and evolution of ‘green’ GSK3/Shaggy-like kinases. Trends Plant Sci. 2012, 17, 39–46. [Google Scholar] [CrossRef]
- Yoo, M.-J.; Albert, V.A.; Soltis, P.S.; Soltis, D.E. Phylogenetic diversification of glycogen synthase kinase 3/SHAGGY-like kinase genes in plants. BMC Plant Biol. 2006, 6, 3. [Google Scholar] [CrossRef]
- Groszyk, J.; Yanushevska, Y.; Zielezinski, A.; Nadolska-Orczyk, A.; Karlowski, W.M.; Orczyk, W. Annotation and profiling of barley GLYCOGEN SYNTHASE3/Shaggy-like genes indicated shift in organ-preferential expression. PLoS ONE 2018, 13, e0199364. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Biju, S.; Bhardwaj, R. 24-Epibrassinolide; an active brassinolide and its role in salt stress tolerance in plants: A review. Plant Physiol. Biochem. 2018, 130, 69–79. [Google Scholar] [CrossRef]
- De Rybel, B.; Audenaert, D.; Vert, G.; Rozhon, W.; Mayerhofer, J.; Peelman, F.; Coutuer, S.; Denayer, T.; Jansen, L.; Nguyen, L.; et al. Chemical inhibition of a subset of Arabidopsis thaliana GSK3-like kinases activates brassinosteroid signaling. Chem. Biol. 2009, 16, 594–604. [Google Scholar] [CrossRef]
- Bajguz, A.; Chmur, M.; Gruszka, D. Comprehensive overview of the brassinosteroid biosynthesis pathways: Substrates, products, inhibitors, and connections. Front. Plant Sci. 2020, 11, 1034. [Google Scholar] [CrossRef]
- Min, H.J.; Cui, L.H.; Oh, T.R.; Kim, J.H.; Kim, T.W.; Kim, W.T. OsBZR 1 turnover mediated by Os SK 22-regulated U-box E3 ligase Os PUB 24 in rice BR response. Plant J. 2019, 99, 426–438. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, J.-Q.; Zhang, X.; Zhou, J.; Jiang, Z.; Huang, P.; Tang, Z.; Bao, Y.; Cheng, J.; Tang, H. Rice qGL3/OsPPKL1 functions with the GSK3/SHAGGY-like kinase OsGSK3 to modulate brassinosteroid signaling. Plant Cell 2019, 31, 1077–1093. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Liang, W.; Cui, X.; Chen, M.; Yin, C.; Luo, Z.; Zhu, J.; Lucas, W.J.; Wang, Z.; Zhang, D. Brassinosteroids promote development of rice pollen grains and seeds by triggering expression of Carbon Starved Anther, a MYB domain protein. Plant J. 2015, 82, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Ji, Y.; Hu, J.; Guo, R.; Sun, S.; Wang, X. Strigolactones and brassinosteroids antagonistically regulate the stability of the D53–OsBZR1 complex to determine FC1 expression in rice tillering. Mol. Plant 2020, 13, 586–597. [Google Scholar] [CrossRef] [PubMed]
- UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Howe, K.L.; Contreras-Moreira, B.; De Silva, N.; Maslen, G.; Akanni, W.; Allen, J.; Alvarez-Jarreta, J.; Barba, M.; Bolser, D.M.; Cambell, L. Ensembl Genomes 2020—enabling non-vertebrate genomic research. Nucleic Acids Res. 2020, 48, D689–D695. [Google Scholar] [CrossRef]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef]
- Wang, H.; Jiao, X.; Kong, X.; Liu, Y.; Chen, X.; Fang, R.; Yan, Y. The histone deacetylase HDA703 interacts with OsBZR1 to regulate rice brassinosteroid signaling, growth and heading date through repression of Ghd7 expression. Plant J. 2020, 104, 447–459. [Google Scholar] [CrossRef]
- Jeong, Y.J.; Corvalán, C.; Kwon, S.I.; Choe, S. Analysis of anti-BZR1 antibody reveals the roles BES1 in maintaining the BZR1 levels in Arabidopsis. J. Plant Biol. 2015, 58, 87–95. [Google Scholar] [CrossRef]
- Zhao, M.; Tang, S.; Zhang, H.; He, M.; Liu, J.; Zhi, H.; Sui, Y.; Liu, X.; Jia, G.; Zhao, Z. DROOPY LEAF1 controls leaf architecture by orchestrating early brassinosteroid signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 21766–21774. [Google Scholar] [CrossRef]
- Kim, E.-J.; Lee, S.-H.; Park, C.-H.; Kim, S.-H.; Hsu, C.-C.; Xu, S.; Wang, Z.-Y.; Kim, S.-K.; Kim, T.-W. Plant U-box40 mediates degradation of the brassinosteroid-responsive transcription factor BZR1 in Arabidopsis roots. Plant Cell 2019, 31, 791–808. [Google Scholar] [CrossRef]
- Yang, M.; Wang, X. Multiple ways of BES1/BZR1 degradation to decode distinct developmental and environmental cues in plants. Mol. Plant 2017, 10, 915–917. [Google Scholar] [CrossRef]
- Yang, M.; Li, C.; Cai, Z.; Hu, Y.; Nolan, T.; Yu, F.; Yin, Y.; Xie, Q.; Tang, G.; Wang, X. SINAT E3 ligases control the light-mediated stability of the brassinosteroid-activated transcription factor BES1 in Arabidopsis. Dev. Cell 2017, 41, 47–58.e44. [Google Scholar] [CrossRef]
- Rozhon, W.; Wang, W.; Berthiller, F.; Mayerhofer, J.; Chen, T.; Petutschnig, E.; Sieberer, T.; Poppenberger, B.; Jonak, C. Bikinin-like inhibitors targeting GSK3/Shaggy-like kinases: Characterisation of novel compounds and elucidation of their catabolism in planta. BMC Plant Biol. 2014, 14, 172. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.; Zheng, X.; Wu, F.; Lin, Q.; Heng, Y.; Tian, P.; Cheng, Z.; Yu, X.; Zhou, K. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice. Nat. Plants 2017, 3, 17043. [Google Scholar] [CrossRef]
- Vukašinović, N.; Wang, Y.; Vanhoutte, I.; Fendrych, M.; Guo, B.; Kvasnica, M.; Jiroutová, P.; Oklestkova, J.; Strnad, M.; Russinova, E. Local brassinosteroid biosynthesis enables optimal root growth. Nat. Plants 2021, 7, 619–632. [Google Scholar] [CrossRef]
- Lee, H.-S.; Kim, Y.; Pham, G.; Kim, J.W.; Song, J.-H.; Lee, Y.; Hwang, Y.-S.; Roux, S.J.; Kim, S.-H. Brassinazole resistant 1 (BZR1)-dependent brassinosteroid signalling pathway leads to ectopic activation of quiescent cell division and suppresses columella stem cell differentiation. J. Exp. Bot. 2015, 66, 4835–4849. [Google Scholar] [CrossRef]
- Chaiwanon, J.; Wang, Z.Y. Spatiotemporal brassinosteroid signaling and antagonism with auxin pattern stem cell dynamics in Arabidopsis roots. Curr. Biol. 2015, 25, 1031–1042. [Google Scholar] [CrossRef]
- Vilarrasa-Blasi, J.; González-García, M.-P.; Frigola, D.; Fàbregas, N.; Alexiou, K.G.; López-Bigas, N.; Rivas, S.; Jauneau, A.; Lohmann, J.U.; Benfey, P.N. Regulation of plant stem cell quiescence by a brassinosteroid signaling module. Dev. Cell 2014, 30, 36–47. [Google Scholar] [CrossRef]
- Kim, S.-K.; Chang, S.C.; Lee, E.J.; Chung, W.-S.; Kim, Y.-S.; Hwang, S.; Lee, J.S. Involvement of brassinosteroids in the gravitropic response of primary root of maize. Plant Physiol. 2000, 123, 997–1004. [Google Scholar] [CrossRef]
- Kim, T.W.; Lee, S.M.; Joo, S.H.; Yun, H.S.; Lee, Y.; Kaufman, P.B.; Kirakosyan, A.; Kim, S.H.; Nam, K.H.; Lee, J.S. Elongation and gravitropic responses of Arabidopsis roots are regulated by brassinolide and IAA. Plant Cell Environ. 2007, 30, 679–689. [Google Scholar] [CrossRef]
- Li, L.; Xu, J.; Xu, Z.-H.; Xue, H.-W. Brassinosteroids stimulate plant tropisms through modulation of polar auxin transport in Brassica and Arabidopsis. Plant Cell 2005, 17, 2738–2753. [Google Scholar] [CrossRef]
- Inahashi, H.; Shelley, I.J.; Yamauchi, T.; Nishiuchi, S.; Takahashi-Nosaka, M.; Matsunami, M.; Ogawa, A.; Noda, Y.; Inukai, Y. OsPIN2, which encodes a member of the auxin efflux carrier proteins, is involved in root elongation growth and lateral root formation patterns via the regulation of auxin distribution in rice. Physiol. Plant. 2018, 164, 216–225. [Google Scholar] [CrossRef]
- Roddick, J.G. Comparative root growth inhibitory activity of four brassinosteroids. Phytochemistry 1994, 37, 1277–1281. [Google Scholar] [CrossRef]
- Kutschera, U.; Wang, Z.-Y. Brassinosteroid action in flowering plants: A Darwinian perspective. J. Exp. Bot. 2012, 63, 3511–3522. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Organ | Parameters | Genotype | Treatment | ||||
|---|---|---|---|---|---|---|---|
| Mean Square | F | p | Mean Square | F | p | ||
| Shoot | length | 129.407 *** | 237.227 | 0.0000 | 6.445 *** | 11.815 | 0.0000 |
| fresh weight | 2.371 × 10−2 *** | 129.722 | 0.0000 | 1.004 × 10−3 *** | 5.493 | 0.0000 | |
| dry weight | 1.584 × 10−4 *** | 95.728 | 0.0000 | 7.180 × 10−6 *** | 4.339 | 0.0002 | |
| RWC | 4.747 ** | 10.340 | 0.0018 | 2.084 *** | 4.539 | 0.0001 | |
| HvDWF4 | 3.937 × 10−7 ** | 8.924 | 0.0046 | 6.764 × 10−7 *** | 15.330 | 0.0000 | |
| HvBRI1 | 1.912 × 10−3 | 2.345 | 0.1328 | 3.513 × 10−3 *** | 4.308 | 0.0007 | |
| HvBAK1 | 4.246 × 10−3 *** | 572.952 | 0.0000 | 2.322 × 10−5 ** | 3.133 | 0.0069 | |
| HvBSU1 | 2.924 × 10−3 *** | 47.809 | 0.0000 | 1.269 × 10−4 | 2.075 | 0.0591 | |
| HvGSK1.1 | 5.645 × 10−3 * | 4.271 | 0.0447 | 3.885 × 10−3 * | 2.939 | 0.0101 | |
| HvGSK1.2 | 5.311 × 10−2 *** | 30.925 | 0.0000 | 1.850 × 10−3 | 1.077 | 0.3963 | |
| HvGSK1.3 | 0.101 *** | 203.986 | 0.0000 | 1.273 × 10−3 * | 2.560 | 0.0219 | |
| HvGSK2.1 | 0.131 *** | 79.846 | 0.0000 | 3.586 × 10−3 * | 2.180 | 0.0477 | |
| HvGSK2.2 | 1.915 × 10−2 *** | 99.207 | 0.0000 | 7.749 × 10−4 ** | 4.015 | 0.0012 | |
| HvGSK3.1 | 4.833 × 10−3 *** | 39.596 | 0.0000 | 2.983 × 10−4 * | 2.444 | 0.0278 | |
| HvGSK4.1 | 1.091 × 10−2 *** | 71.189 | 0.0000 | 4.283 × 10−4 * | 2.794 | 0.0136 | |
| HvBZR1 | 6.562 × 10−6 *** | 68.418 | 0.0000 | 1.873 × 10−7 | 1.953 | 0.0756 | |
| Root | length | 4.400 | 2.093 | 0.1512 | 49.539 *** | 23.562 | 0.0000 |
| fresh weight | 7.634 × 10−5 | 0.438 | 0.5096 | 4.306 × 10−3 *** | 24.715 | 0.0000 | |
| dry weight | 9.600 × 10−6 ** | 7.274 | 0.0082 | 1.508 × 10−5 *** | 11.428 | 0.0000 | |
| RWC | 13.306 ** | 7.328 | 0.0080 | 20.335 *** | 11.199 | 0.0000 | |
| HvDWF4 | 0.226 | 1.008 | 0.3209 | 0.218 | 0.974 | 0.4688 | |
| HvBRI1 | 3.745 × 10−3 | 0.222 | 0.6393 | 2.975 × 10−2 | 1.760 | 0.1121 | |
| HvBAK1 | 4.879 × 10−2 | 3.178 | 0.0817 | 1.548 × 10−2 | 1.008 | 0.4442 | |
| HvBSU1 | 1.788 × 10−2 | 1.430 | 0.2384 | 1.163 × 10−2 | 0.930 | 0.5017 | |
| HvGSK1.1 | 5.917 × 10−2 * | 4.195 | 0.0467 | 1.689 × 10−2 | 1.193 | 0.3233 | |
| HvGSK1.2 | 3.394 × 10−2 | 2.637 | 0.1117 | 1.371 × 10−2 | 1.065 | 0.4051 | |
| HvGSK1.3 | 8.160 × 10−3 | 0.539 | 0.4669 | 1.589 × 10−2 | 1.049 | 0.4156 | |
| HvGSK2.1 | 0.111 ** | 9.515 | 0.0036 | 9.758 × 10−3 | 0.838 | 0.5747 | |
| HvGSK2.2 | 4.686 × 10−2 | 2.950 | 0.0931 | 1.463 × 10−2 | 0.921 | 0.5087 | |
| HvGSK3.1 | 3.283 × 10−2 | 1.368 | 0.2485 | 2.429 × 10−2 | 1.012 | 0.4412 | |
| HvGSK4.1 | 2.724 × 10−2 | 2.160 | 0.1489 | 1.036 × 10−2 | 0.821 | 0.5882 | |
| HvBZR1 | 9.796 × 10−4 | 0.519 | 0.4751 | 1.752 × 10−2 | 0.928 | 0.5031 | |
| Shoot-to-root ratio | by length | 0.677 *** | 31.750 | 0.0000 | 0.621 *** | 29.098 | 0.0000 |
| by fresh weight | 4.545 *** | 180.744 | 0.0000 | 1.361 *** | 54.135 | 0.0000 | |
| by dry weight | 6.411 *** | 115.179 | 0.0000 | 1.111 *** | 19.953 | 0.0000 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Groszyk, J.; Szechyńska-Hebda, M. Brassinazole Resistant 1 Activity Is Organ-Specific and Genotype-Dependent in Barley Seedlings. Int. J. Mol. Sci. 2021, 22, 13572. https://doi.org/10.3390/ijms222413572
Groszyk J, Szechyńska-Hebda M. Brassinazole Resistant 1 Activity Is Organ-Specific and Genotype-Dependent in Barley Seedlings. International Journal of Molecular Sciences. 2021; 22(24):13572. https://doi.org/10.3390/ijms222413572
Chicago/Turabian StyleGroszyk, Jolanta, and Magdalena Szechyńska-Hebda. 2021. "Brassinazole Resistant 1 Activity Is Organ-Specific and Genotype-Dependent in Barley Seedlings" International Journal of Molecular Sciences 22, no. 24: 13572. https://doi.org/10.3390/ijms222413572
APA StyleGroszyk, J., & Szechyńska-Hebda, M. (2021). Brassinazole Resistant 1 Activity Is Organ-Specific and Genotype-Dependent in Barley Seedlings. International Journal of Molecular Sciences, 22(24), 13572. https://doi.org/10.3390/ijms222413572