Low Basicity as a Characteristic for Atypical Ligands of Serotonin Receptor 5-HT2

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. History of 5-HT2R Ligands with Low Basicity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
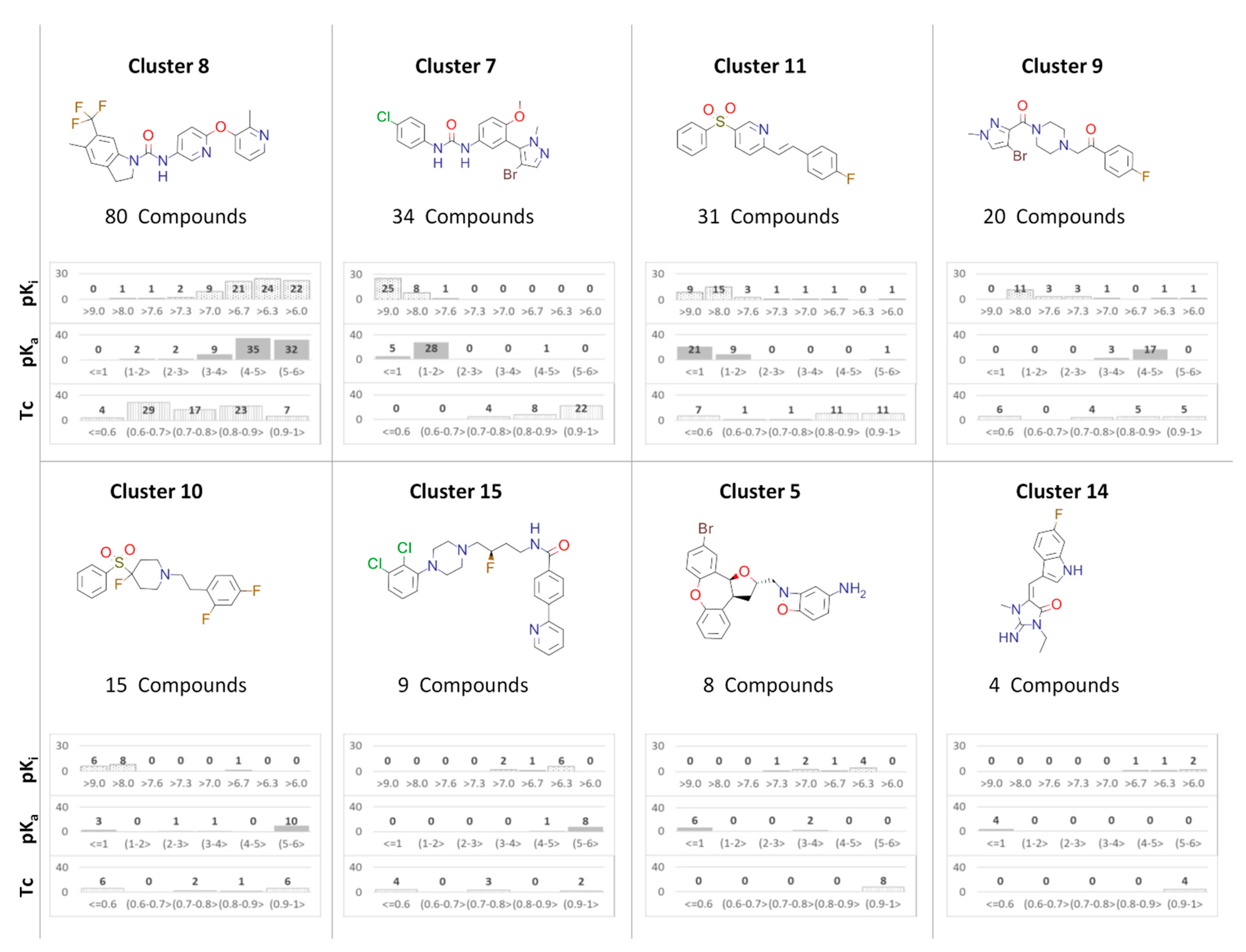
{kind=link}
| 5-HT2A/Cluster 8 [37,38,39,40,41,43,45], (80 cmpds) | 5-HT2A/Cluster 7 [45], (34 cmpds) | ||
 pKa = 5.44 |  pKa = 8.39 [43] |  pKa = 1.29 |  pKa = 9.68 [58] |
| 5-HT2A/Cluster 11 [26,50], (31 cmpds) | 5-HT2A/Cluster 9 [48], (20 cmpds) | ||
 pKa = 0.39 |  pKa = 7.00 [59] |  pKa = 4.23 |  pKa = 6.96 [48] |
| 5-HT2A/Cluster 10 [23,26,49], (15 cmpds) | 5-HT2A/Cluster 15 [60], (9 cmpds) | ||
 pKa = 5.88 |  pKa = 7.00 [59] |  pKa = 5.90 |  pKa = 7.33 [61] |
| 5-HT2A/Cluster 5 [62], (8 cmpds) | 5-HT2A/Cluster 14 [54], (4 cmpds) | ||
 pKa = 3.35 |  pKa = 8.80 [62] |  pKa = 9.33 |  pKa = 9.55 [63] |
| 5-HT2B/Cluster 16 [33,34,35,36,37], (88 cmpds) | 5-HT2B/Cluster 1, 2 [55], (33 cmpds) | ||
 pKa = 5.44 |  pKa = 8.48 [43] |  pKa = 5.47 |  pKa = 9.56 [64] |
| 5-HT2B/Cluster 22 [65] (20 cmpds) | 5-HT2B/Cluster 23 [53] (16 cmpds) | ||
 pKa = −4.71 |  pKa = 1.73 | ||
| 5-HT2B/Cluster 18 [46] (8 cmpds) | 5-HT2B/Cluster 10 [66] (4 cmpds) | ||
 pKa = 5.05 |  pKa = 1.58 | ||
| 5-HT2B/Cluster 13 [67] (4 cmpds) | 5-HT2B/Cluster 20 [57,62] (2 cmpds) | ||
 pKa = −4.56 |  pKa = 3.66 |  pKa = 9.75 [68] | |
| 5-HT2C/Cluster 16 [37,38,39,40,41,42,43,45], (150 cmpds) | 5-HT2C/Cluster 15 [44,45,46,47], (46 cmpds) | ||
 pKa = 4.33 |  pKa = 9.92 [54] |  pKa = 1.29 |  pKa = 10.04 [69] |
| 5-HT2C/Cluster 11 [23,49,50,59], (31 cmpds) | 5-HT2C/Cluster 14, 17 [54,55,56], (16 cmpds) | ||
 pKa = 0.36 |  pKa = 7.00 [59] |  pKa = 5.47 |  pKa = 9.55 [70] |
| 5-HT2C/Cluster 2 [53,71], (15 cmpds) | 5-HT2C/Cluster 9 [51,52,72], (13 cmpds) | ||
 pKa = 1.73 |  pKa = −1.44 |  pKa = 9.78 [73] | |
| 5-HT2C /Cluster 3 [62], (7 cmpds) | 5-HT2C /Cluster 4 [68], (1 cmpds) | ||
 pKa = 3.35 |  pKa = 8.80 [62] |  pKa = 5.71 |  pKa = 9.85 [68] |
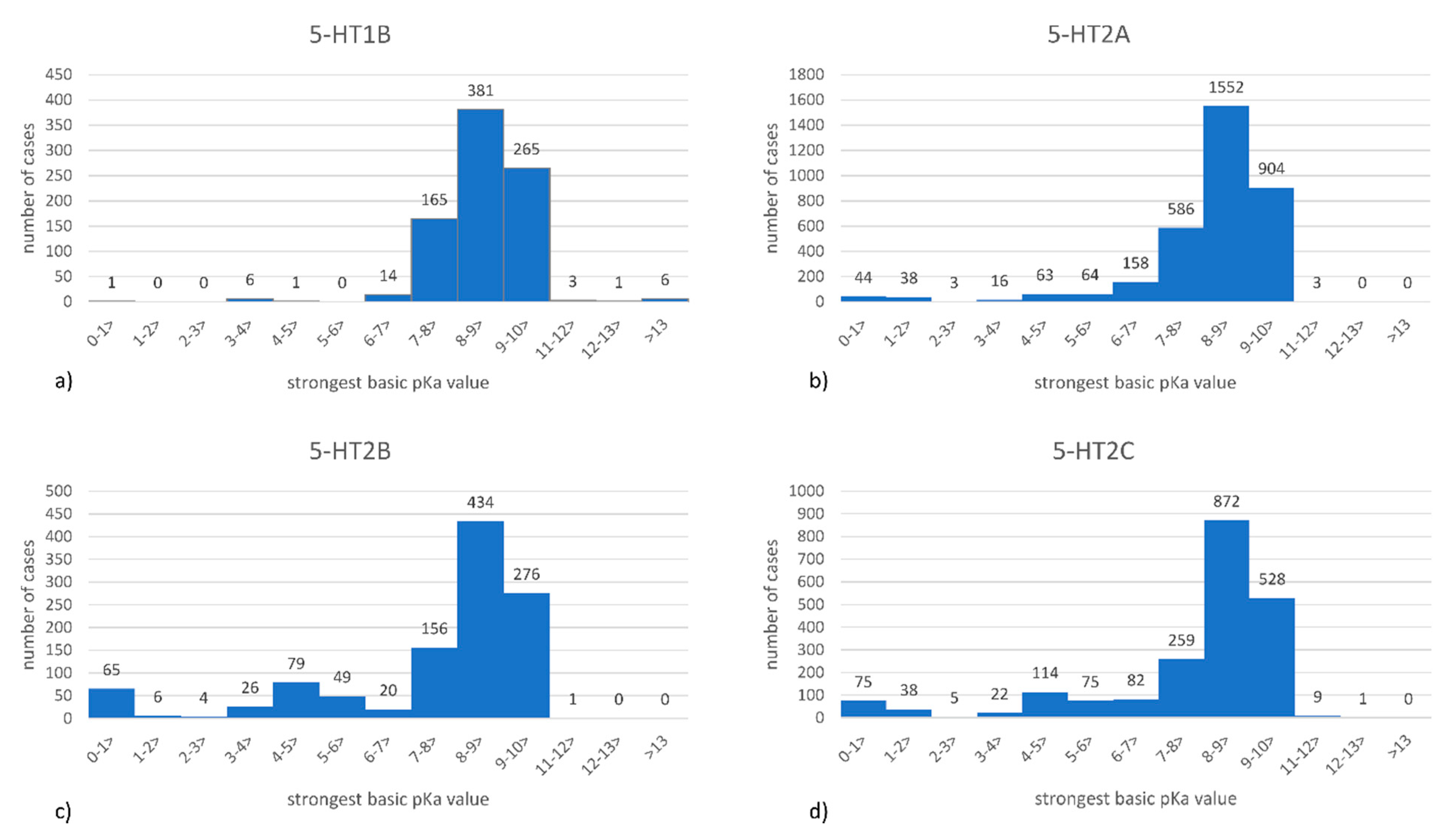
2.2. Datasets Analysis
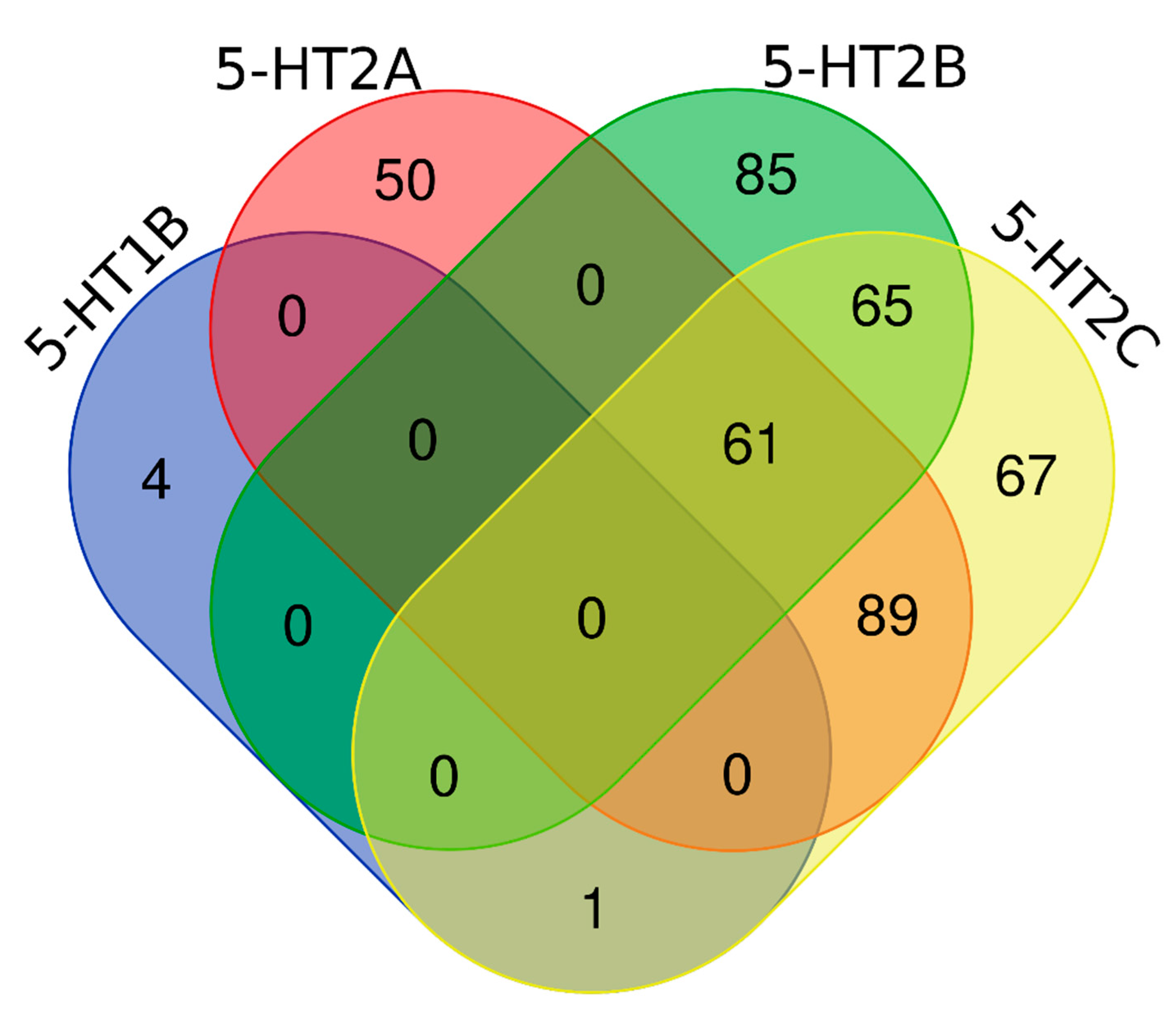
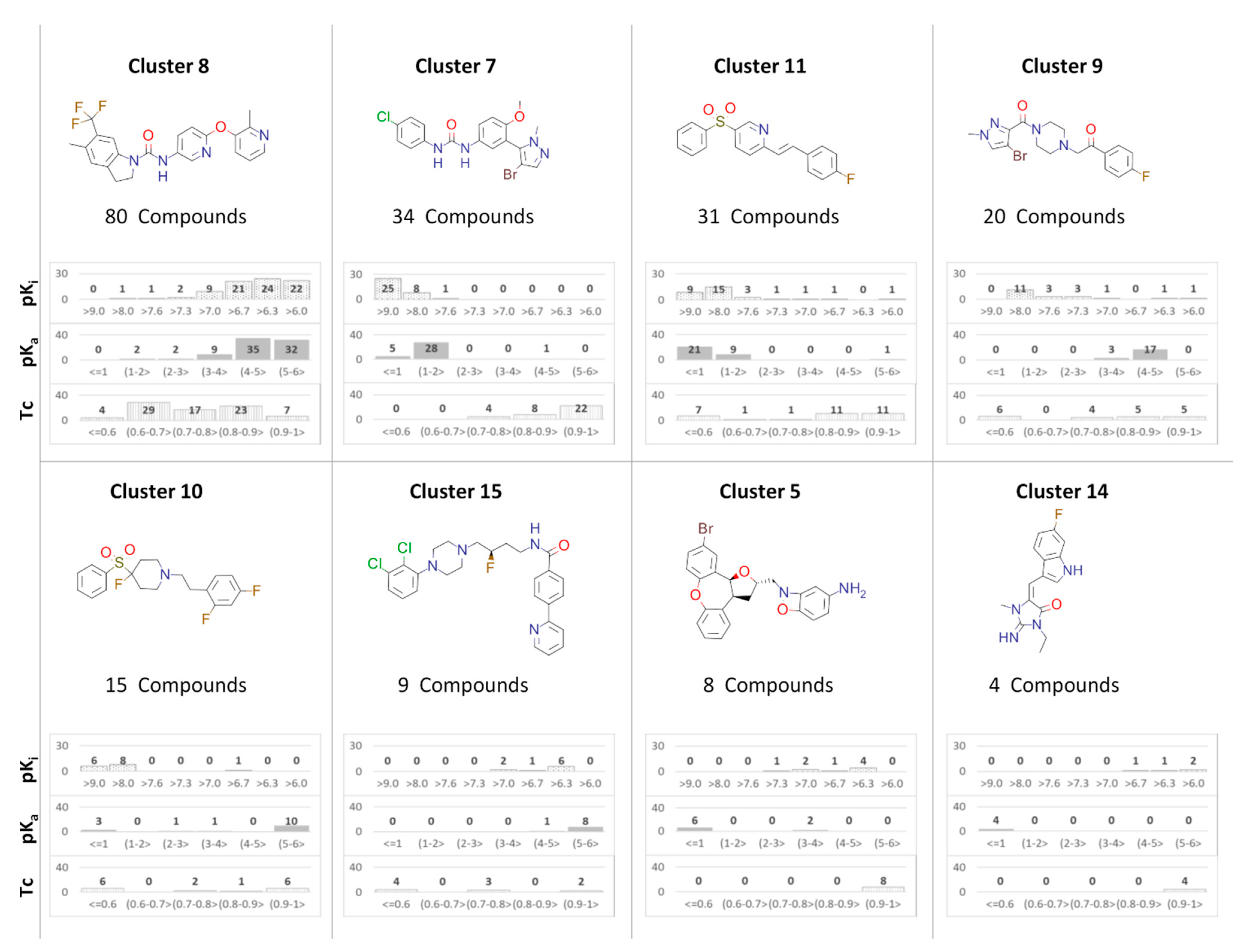
2.3. Analysis of Nonbasic Ligands by Clusters
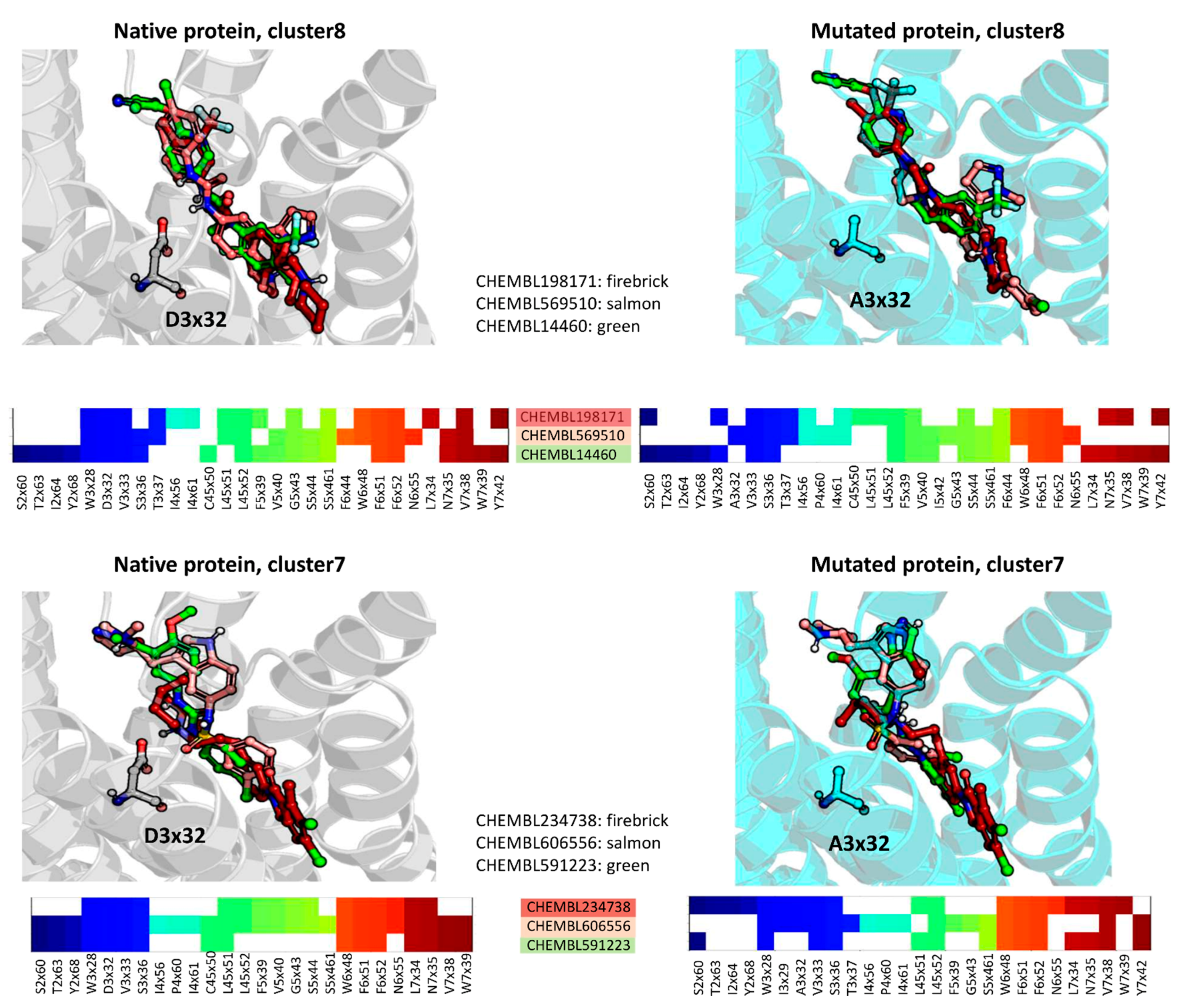
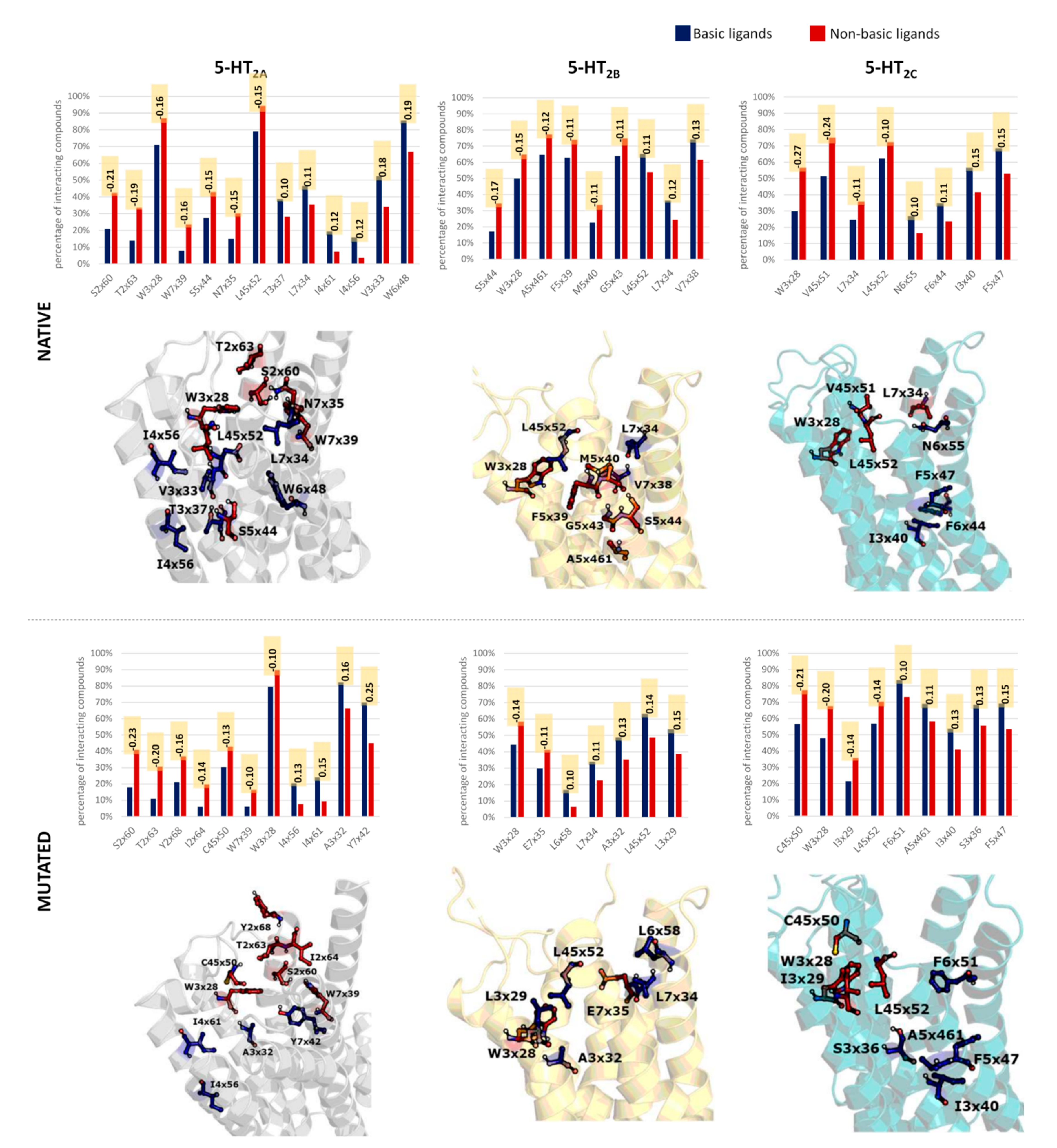
2.4. Docking Analysis—By Clusters
2.5. Docking Studies—Whole Dataset Analysis
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 5-HTRs | serotonin receptors |
| CNS | central nervous system |
| GPCRs | G protein-coupled receptors |
| TMDs | transmembrane domains |
| ICL | intracellular loop |
| ECL | extracellular loop |
| SDM | site-directed mutagenesis |
| Tc | Tanimoto coefficient |
| SIFt | Structural Interaction Fingerprint |
References
- De Deurwaerdère, P.; Di Giovanni, G. Serotonin in health and disease. Int. J. Mol. Sci. 2020, 21, 3500. [Google Scholar] [CrossRef] [PubMed]
- Tsegay, E.W.; Demise, D.G.; Hailu, N.A.; Gufue, Z.H. Serotonin Type 6 and 7 Receptors as a Novel Therapeutic Target for the Treatment of Schizophrenia. Neuropsychiatr Dis. Treat. 2020, 16, 2499–2509. [Google Scholar] [CrossRef]
- Štrac, D.Š.; Pivac, N.; Mück-Šeler, D. The serotonergic system and cognitive function. Transl. Neurosci. 2016, 7, 35–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artigas, F. Serotonin receptors involved in antidepressant effects. Pharmacol. Ther. 2013, 137, 119–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seyedabadi, M.; Fakhfouri, G.; Ramezani, V.; Mehr, S.E.; Rahimian, R. The role of serotonin in memory: Interactions with neurotransmitters and downstream signaling. Exp. Brain Res. 2014, 232, 723–738. [Google Scholar] [CrossRef]
- Maia, T.V.; Cano-Colino, M. The Role of Serotonin in Orbitofrontal Function and Obsessive-Compulsive Disorder. Clin. Psychol. Sci. 2015, 3, 460–482. [Google Scholar] [CrossRef] [Green Version]
- Villalón, C.M. The Role of Serotonin Receptors in the Control of Cardiovascular Function. In The Serotonin System: History, Neuropharmacology, and Pathology; Tricklebank, M., Daly, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Palacios, J.M. Serotonin receptors in brain revisited. Brain Res. 2016, 1645, 46–49. [Google Scholar] [CrossRef]
- Ohno, Y. Serotonin Receptors as the Therapeutic Target for Central Nervous System Disorders. In Serotonin: The Mediator That Spans Evolution; Pilowsky, P.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Hoyer, D. Targeting the 5-HT system: Potential side effects. Neuropharmacology 2020, 179, 108233. [Google Scholar] [CrossRef]
- Madia, V.N.; Messore, A.; Saccoliti, F.; Tudino, V.; De Leo, A.; De Vita, D.; Bortolami, M.; Scipione, L.; Pindinello, I.; Costi, R.; et al. Tegaserod for the Treatment of Irritable Bowel Syndrome. Antiinflamm. Antiallergy. Agents Med. Chem. 2020, 19, 342–369. [Google Scholar] [CrossRef]
- Magnusson, J.L.; Cummings, K.J. Central serotonin and the control of arterial blood pressure and heart rate in infant rats: Influence of sleep state and sex. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R313–R321. [Google Scholar] [CrossRef] [PubMed]
- Ott, M.; Mannchen, J.K.; Jamshidi, F.; Werneke, U. Management of severe arterial hypertension associated with serotonin syndrome: A case report analysis based on systematic review techniques. Ther. Adv. Psychopharmacol. 2019, 9, 2045125318818814. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Denna, T.H.; Storkersen, J.N.; Gerriets, V.A. Beyond a neurotransmitter: The role of serotonin in inflammation and immunity. Pharmacol. Res. 2019, 140, 100–114. [Google Scholar] [CrossRef] [PubMed]
- Heath, M.J.; Hen, R. Serotonin receptors. Genetic insights into serotonin function. Curr. Biol. 1995, 5, 997–999. [Google Scholar] [CrossRef] [Green Version]
- Nichols, D.E.; Nichols, C.D. Serotonin receptors. Chem. Rev. 2008, 108, 1614–1641. [Google Scholar] [CrossRef]
- Bachman, J. Chapter Ninteen-Site-Directed Mutagenesis. In Methods in Enzymology; Abelson, J., Simon, M., Verdine, G., Pyle, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Impellizzeri, A.A.R.; Pappalardo, M.; Basile, L.; Manfra, O.; Andressen, K.W.; Krobert, K.A.; Messina, A.; Levy, F.O.; Guccione, S. Identification of essential residues for binding and activation in the human 5-HT7(a) serotonin receptor by molecular modeling and site-directed mutagenesis. Front. Behav. Neurosci. 2015, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Boess, F.G.; Monsma, F.J.; Sleight, A.J. Identification of residues in transmembrane regions III and VI that contribute to the ligand binding site of the serotonin 5-HT6 receptor. J. Neurochem. 1998, 71, 2169–2177. [Google Scholar] [CrossRef]
- Xie, J.; Dernovici, S.; Ribeiro, P. Mutagenesis analysis of the serotonin 5-HT2C receptor and a Caenorhabditis elegans 5-HT2 homologue: Conserved residues of helix 4 and helix 7 contribute to agonist-dependent activation of 5-HT2 receptors. J. Neurochem. 2005, 92, 375–387. [Google Scholar] [CrossRef]
- Manivet, P.; Schneider, B.; Smith, J.C.; Choi, D.S.; Maroteaux, L.; Kellermann, O.; Launay, J.M. The serotonin binding site of human and murine 5-HT 2B receptors. Molecular modeling and site-directed mutagenesis. J. Biol. Chem. 2002, 277, 17170–17178. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Ha, H.-J.; Chi, D.; Lee, H. Serotonin Receptor and Transporter Ligands-Current Status. Curr. Med. Chem. 2001, 8, 999–1034. [Google Scholar] [CrossRef]
- Ladduwahetty, T.; Boase, A.L.; Mitchinson, A.; Quin, C.; Patel, S.; Chapman, K.; MacLeod, A.M. A new class of selective, non-basic 5-HT2A receptor antagonists. Bioorg. Med. Chem. Lett. 2006, 16, 3201–3204. [Google Scholar] [CrossRef]
- Staroń, J.; Kurczab, R.; Warszycki, D.; Satała, G.; Krawczyk, M.; Bugno, R.; Lenda, T.; Popik, P.; Hogendorf, A.S.; Hogendorf, A.; et al. Virtual screening-driven discovery of dual 5-HT6/5-HT2A receptor ligands with pro-cognitive properties. Eur. J. Med. Chem. 2019, 111857. [Google Scholar] [CrossRef] [PubMed]
- Staroń, J.; Mordalski, S.; Warszycki, D.; Satała, G.; Hogendorf, A.; Bojarski, A.J. Pyrano[2,3,4-cd]indole as a Scaffold for Selective Nonbasic 5-HT6R Ligands. ACS Med. Chem. Lett. 2017, 8, 390–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladduwahetty, T.; Gilligan, M.; Humphries, A.; Merchant, K.J.; Fish, R.; McAlister, G.; Ivarsson, M.; Dominguez, M.; O’Connor, D.; MacLeod, A.M. Non-basic ligands for aminergic GPCRs: The discovery and development diaryl sulfones as selective, orally bioavailable 5-HT2A receptor antagonists for the treatment of sleep disorders. Bioorg. Med. Chem. Lett. 2010, 20, 3708–3712. [Google Scholar] [CrossRef] [PubMed]
- Jastrzebski, S.; Podlewska, S.; Czarnecki, W.M.; Bojarski, A.J. Quo vadis G protein-coupled receptor ligands? A tool for analysis of the emergence of new groups of compounds over time. Bioorg. Med. Chem. Lett. 2017, 27, 626–631. [Google Scholar] [CrossRef]
- Vandenberg, J.I.; Perry, M.D.; Perrin, M.J.; Mann, S.A.; Ke, Y.; Hill, A.P. hERG K(+) channels: Structure, function, and clinical significance. Physiol. Rev. 2012, 92, 1393–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wacker, D.; Wang, C.; Katritch, V.; Han, G.W.; Huang, X.-P.; Vardy, E.; McCorvy, J.D.; Jiang, Y.; Chu, M.; Siu, F.Y.; et al. Structural features for functional selectivity at serotonin receptors. Science 2013, 340, 615–619. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Jiang, Y.; Ma, J.; Wu, H.; Wacker, D.; Katritch, V.; Han, G.W.; Liu, W.; Huang, X.P.; Vardy, E.; et al. Structural Basis for Molecular Recognition at Serotonin Receptors. Science 2013, 340, 610–614. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wacker, D.; Gati, C.; Han, G.W.; James, D.; Wang, D.; Nelson, G.; Weierstall, U.; Katritch, V.; Barty, A.; et al. Serial femtosecond crystallography of G protein-coupled receptors. Science 2013, 342, 1521–1524. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Gaulton, A.; Bellis, L.J.; Bento, A.P.; Chambers, J.; Davies, M.; Hersey, A.; Light, Y.; McGlinchey, S.; Michalovich, D.; Al-Lazikani, B.; et al. ChEMBL: A large-scale bioactivity database for drug discovery. Nucleic Acids Res. 2011, 40, D1100–D1107. [Google Scholar] [CrossRef] [Green Version]
- Kennett, G.A.; Wood, M.D.; Glen, A.; Grewal, S.; Forbes, I.; Gadre, A.; Blackburn, T.P. In vivo properties of SB 200646A, a 5-HT2C/2B receptor antagonist. Br. J. Pharmacol. 1994, 111, 1476–5381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, I.T.; Kennett, G.A.; Gadre, A.; Ham, P.; Hayward, C.J.; Martin, R.T.; Thompson, M.; Wood, M.D.; Baxter, G.S.; Glen, A.; et al. N-(1-Methyl-5-indolyl)-N′-(3-pyridyl)urea Hydrochloride: The First Selective 5-HT1C Receptor Antagonist. J. Med. Chem. 1993, 36, 1104–1107. [Google Scholar] [CrossRef] [PubMed]
- Fludzinski, P.; Wittenauer, L.A.; Schenck, K.W.; Cohen, M.L. 2,3-Dialkyl(diraethylamino)indoles: Interaction with 5HT1, 5HT2, and Rat Stomach Fundal Serotonin Receptors. J. Med. Chem. 1986, 29, 2415–2418. [Google Scholar] [CrossRef] [PubMed]
- Bromidge, S.M.; Duckworth, M.; Forbes, I.T.; Ham, P.; King, F.D.; Thewlis, K.M.; Blaney, F.E.; Naylor, C.B.; Blackburn, T.P.; Kennett, G.A.; et al. 6-chloro-5-methyl-1-[[2-[(2-methyl-3-pyridyl)oxy]-5-pyridyl]carbamoyl]- indoline (SB-242084): The first selective and brain penetrant 5-HT(2C) receptor antagonist. J. Med. Chem. 1997, 40, 3494–3496. [Google Scholar] [CrossRef] [PubMed]
- Bromidge, S.M.; Dabbs, S.; Davies, S.; Duckworth, D.M.; Forbes, I.T.; Jones, G.E.; Jones, J.; King, F.D.; Saunders, D.V.; Blackburn, T.P.; et al. 1-[2-[(Heteroaryloxy)heteroaryl]carbamoyl]indolines: Novel and selective 5-HT(2C) receptor inverse agonists with potential as antidepressant/anxiolytic agents. Bioorg. Med. Chem. Lett. 2000, 10, 1863–1866. [Google Scholar] [CrossRef]
- Bromidge, S.M.; Davies, S.; Duckworth, D.M.; Forbes, I.T.; Jones, G.E.; Jones, J.; King, F.D.; Blackburn, T.P.; Holland, V.; Kennett, G.A.; et al. 1-[2-[(Heteroarylmethoxy)aryl]carbamoyl]indolines are selective and orally active 5-HT(2C) receptor inverse agonists. Bioorg. Med. Chem. Lett. 2000, 10, 1867–1870. [Google Scholar] [CrossRef]
- Bromidge, S.M.; Dabbs, S.; Davies, D.T.; Davies, S.; Duckworth, D.M.; Forbes, I.T.; Gaster, L.M.; Ham, P.; Jones, G.E.; King, F.D.; et al. Biarylcarbamoylindolines are novel and selective 5-HT(2C) receptor inverse agonists: Identification of 5-methyl-1-[[2-[(2-methyl-3- pyridyl)oxy]5-pyridyl]carbamoyl]-6-trifluoromethylindoline (SB-243213) as a potential antidepressant/anxiolytic agent. J. Med. Chem. 2000, 43, 1123–1134. [Google Scholar] [CrossRef]
- Bromidge, S.M.; Dabbs, S.; Davies, D.T.; Duckworth, D.M.; Forbes, I.T.; Ham, P.; Jones, G.E.; King, F.D.; Saunders, D.V.; Starr, S.; et al. Novel and selective 5-HT(2C/2B) receptor antagonists as potential anxiolytic agents: Synthesis, quantitative structure-Activity relationships, and molecular modeling of substituted 1-(3- pyridylcarbamoyl)indolines. J. Med. Chem. 1998, 41, 1598–1612. [Google Scholar] [CrossRef]
- Forbes, I.T.; Ham, P.; Booth, D.H.; Martin, R.T.; Thompson, M.; Baxter, G.S.; Blackburn, T.P.; Glen, A.; Kennett, G.A.; Wood, M.D. 5-Methyl-1-(3-pyridylcarbamoyl)-1,2,3,5-tetrahydropyrrolo[2,3-f]indole: A Novel 5-HT2C/5-HT2B Receptor Antagonist with Improved Affinity, Selectivity, and Oral Activity. J. Med. Chem. 1995, 38, 2524–2530. [Google Scholar] [CrossRef]
- Goodacre, C.J.; Bromidge, S.M.; Clapham, D.; King, F.D.; Lovell, P.J.; Allen, M.; Campbell, L.P.; Holland, V.; Riley, G.J.; Starr, K.R.; et al. A series of bisaryl imidazolidin-2-ones has shown to be selective and orally active 5-HT2C receptor antagonists. Bioorg. Med. Chem. Lett. 2005, 15, 4989–4993. [Google Scholar] [CrossRef]
- Forbes, I.T.; Jones, G.E.; Murphy, O.E.; Holland, V.; Baxter, G.S. N-(1-Methyl-5-indolyl)-N′-(3-methyl-5-isothiazolyl)urea: A Novel, High-Affinity 5-HT2B Receptor Antagonist. J. Med. Chem. 1995, 38, 855–857. [Google Scholar] [CrossRef] [PubMed]
- Teegarden, B.R.; Li, H.; Jayakumar, H.; Strah-Pleynet, S.; Dosa, P.I.; Selaya, S.D.; Kato, N.; Elwell, K.H.; Davidson, J.; Cheng, K.; et al. Discovery of 1-[3-(4-Bromo-2-methyl-2H-pyrazol-3-yl)-4-methoxyphenyl]-3-(2, 4-difluorophenyl)urea (Nelotanserin) and Related 5-Hydroxytryptamine 2A Inverse Agonists for the Treatment of Insomnia. J. Med. Chem. 2010, 53, 1923–1936. [Google Scholar] [CrossRef] [PubMed]
- Porter, R.A.; Chan, W.N.; Coulton, S.; Johns, A.; Hadley, M.S.; Widdowson, K.; Jerman, J.C.; Brough, S.J.; Coldwell, M.; Smart, D.; et al. 1,3-Biarylureas as selective non-peptide antagonists of the orexin-1 receptor. Bioorg. Med. Chem. Lett. 2001, 11, 1907–1910. [Google Scholar] [CrossRef]
- Lin, X.; Huang, X.P.; Chen, G.; Whaley, R.; Peng, S.; Wang, Y.; Zhang, G.; Wang, S.X.; Wang, S.; Roth, B.L.; et al. Life beyond kinases: Structure-based discovery of sorafenib as nanomolar antagonist of 5-HT receptors. J. Med. Chem. 2012, 55, 5749–5759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Ullman, B.; Choi, J.S.K.; Cherrier, M.; Strah-Pleynet, S.; Decaire, M.; Dosa, P.I.; Feichtinger, K.; Teegarden, B.R.; Frazer, J.M.; et al. Synthesis and in vivo evaluation of phenethylpiperazine amides: Selective 5-hydroxytryptamine2A receptor antagonists for the treatment of insomnia. J. Med. Chem. 2010, 53, 5696–5706. [Google Scholar] [CrossRef]
- Fish, L.R.; Gilligan, M.T.; Humphries, A.C.; Ivarsson, M.; Ladduwahetty, T.; Merchant, K.J.; O’Connor, D.; Patel, S.; Philipps, E.; Vargas, H.M.; et al. 4-Fluorosulfonylpiperidines: Selective 5-HT2A ligands for the treatment of insomnia. Bioorg. Med. Chem. Lett. 2005, 15, 3665–3669. [Google Scholar] [CrossRef]
- Wilson, K.J.; van Niel, M.B.; Cooper, L.; Bloomfield, D.; O’Connor, D.; Fish, L.R.; MacLeod, A.M. 2,5-Disubstituted pyridines: The discovery of a novel series of 5-HT2A ligands. Bioorg. Med. Chem. Lett. 2007, 17, 2643–2648. [Google Scholar] [CrossRef]
- Ettaoussi, M.; Sabaouni, A.; Rami, M.; Boutin, J.A.; Delagrange, P.; Renard, P.; Spedding, M.; Caignard, D.H.; Berthelot, P.; Yous, S. Design, synthesis and pharmacological evaluation of new series of naphthalenic analogues as melatoninergic (MT 1/MT 2) and serotoninergic 5-HT 2C dual ligands (I). Eur. J. Med. Chem. 2012, 49, 310–323. [Google Scholar] [CrossRef]
- Ettaoussi, M.; Pérès, B.; Jarry, C.; Nosjean, O.; Boutin, J.A.; Gohier, A.; Mannoury La Cour, C.; Caignard, D.H.; Delagrange, P.; Berthelot, P.; et al. Synthesis, chiral resolution, absolute configuration assignment and pharmacological evaluation of a series of melatoninergic ligands. MedChemComm 2014, 5, 1303–1308. [Google Scholar] [CrossRef]
- Tosh, D.K.; Ciancetta, A.; Warnick, E.; Crane, S.; Gao, Z.G.; Jacobson, K.A. Structure-Based Scaffold Repurposing for G Protein-Coupled Receptors: Transformation of Adenosine Derivatives into 5HT2B/5HT2C Serotonin Receptor Antagonists. J. Med. Chem. 2016, 59, 11006–11026. [Google Scholar] [CrossRef] [Green Version]
- Cummings, D.F.; Canseco, D.C.; Sheth, P.; Johnson, J.E.; Schetz, J.A. Synthesis and structure-affinity relationships of novel small molecule natural product derivatives capable of discriminating between serotonin 5-HT1A, 5-HT2A, 5-HT2C receptor subtypes. Bioorg. Med. Chem. 2010, 18, 4783–4792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewellyn, K.; Bialonska, D.; Loria, M.J.; White, S.W.; Sufka, K.J.; Zjawiony, J.K. In vitro structure-activity relationships of aplysinopsin analogs and their in vivo evaluation in the chick anxiety-depression model. Bioorg. Med. Chem. 2013, 21, 7083–7390. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.F.; Schetz, J.A.; Kelly, M.; Peng, J.N.; Ang, K.K.H.; Flotow, H.; Leong, C.Y.; Ng, S.B.; Buss, A.D.; Wilkins, S.P.; et al. New antiinfective and human 5-HT2 receptor binding natural and semisynthetic compounds from the Jamaican sponge Smenospongia aurea. J. Nat. Prod. 2002, 65, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Lebar, M.D.; Hahn, K.N.; Mutka, T.; Maignan, P.; McClintock, J.B.; Amsler, C.D.; Van Olphen, A.; Kyle, D.E.; Baker, B.J. CNS and antimalarial activity of synthetic meridianin and psammopemmin analogs. Bioorg. Med. Chem. 2011, 19, 5756–5762. [Google Scholar] [CrossRef] [PubMed]
- Cole, D.C.; Lennox, W.J.; Lombardi, S.; Ellingboe, J.W.; Bernotas, R.C.; Tawa, G.J.; Mazandarani, H.; Smith, D.L.; Zhang, G.; Coupet, J.; et al. Discovery of 5-arylsulfonamido-3-(pyrrolidin-2-ylmethyl)-1H-indole derivatives as potent, selective 5-HT6 receptor agonists and antagonists. J. Med. Chem. 2005, 48, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, S.R.; Burkamp, F.; Blurton, P.; Cheng, S.K.F.; Clarkson, R.; O’Connor, D.; Spinks, D.; Tudge, M.; Van Niel, M.B.; Patel, S.; et al. 4-(Phenylsulfonyl)piperidines: Novel, selective, and bioavailable 5-HT2A receptor antagonists. J. Med. Chem. 2002, 45, 492–503. [Google Scholar] [CrossRef]
- Banala, A.K.; Levy, B.A.; Khatri, S.S.; Furman, C.A.; Roof, R.A.; Mishra, Y.; Griffin, S.A.; Sibley, D.R.; Luedtke, R.R.; Newman, A.H. N -(3-Fluoro-4-(4-(2-methoxy or 2,3-dichlorophenyl)piperazine-1-yl)butyl) arylcarboxamides as selective dopamine D3 receptor ligands: Critical role of the carboxamide linker for d3 receptor selectivity. J. Med. Chem. 2011, 54, 3581–3594. [Google Scholar] [CrossRef] [Green Version]
- Grundt, P.; Prevatt, K.M.; Cao, J.; Taylor, M.; Floresca, C.Z.; Choi, J.K.; Jenkins, B.G.; Luedtke, R.R.; Newman, A.H. Heterocyclic analogues of N-(4-(4-(2,3-dichlorophenyl)piperazin-1-yl)butyl) arylcarboxamides with functionalized linking chains as novel dopamine D3 receptor ligands: Potential substance abuse therapeutic agents. J. Med. Chem. 2007, 50, 4135–4146. [Google Scholar] [CrossRef]
- Bartolomé, J.M.; Alcudia, A.; Andrés, J.I.; Cid, J.M.; García, M.; Megens, A.; Toledo, M.A.; Trabanco, A.A. Novel 2-N,N-dimethylaminomethyl-2,3,3a,12b-tetrahydrodibenzo[b,f]furo[2,3- d]oxepin derivatives displaying combined norepinephrine reuptake inhibition and 5-HT2A/2C receptor antagonism. Bioorg. Med. Chem. Lett. 2005, 15, 2898–2901. [Google Scholar] [CrossRef]
- Blough, B.E.; Landavazo, A.; Partilla, J.S.; Decker, A.M.; Page, K.M.; Baumann, M.H.; Rothman, R.B. Alpha-ethyltryptamines as dual dopamine-serotonin releasers. Bioorg. Med. Chem. Lett. 2014, 24, 4754–4758. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.A.; El-Alfy, A.T.; Ezel, K.; Radwan, M.O.; Shilabin, A.G.; Kochanowska-Karamyan, A.J.; Abd-Alla, H.I.; Otsuka, M.; Hamann, M.T. Marine inspired 2-(5-halo-1H-indol-3-yl)-N,N-dimethylethanamines as modulators of serotonin receptors: An example illustrating the power of bromine as part of the uniquely marine chemical space. Mar. Drugs 2017, 15, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.J.; Saubern, S.; MacDonald, J.M.; Huang, X.P.; Setola, V.; Roth, B.L. N-Tetrahydrothiochromenoisoxazole-1-carboxamides as selective antagonists of cloned human 5-HT2B. Bioorg. Med. Chem. Lett. 2010, 20, 5488–5490. [Google Scholar] [CrossRef] [PubMed]
- Ivachtchenko, A.V.; Dmitriev, D.E.; Golovina, E.S.; Kadieva, M.G.; Koryakova, A.G.; Kysil, V.M.; Mitkin, O.D.; Okun, I.M.; Tkachenko, S.E.; Vorobiev, A.A. (3-Phenylsulfonylcycloalkano[e and d]pyrazolo[1,5-a]pyrimidin-2-yl)amines: Potent and selective antagonists of the serotonin 5-HT6 receptor. J. Med. Chem. 2010, 53, 5186–5196. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, D.L.; Saddler, T.; Auerbach, S.S. An Overview of National Toxicology Program’s Toxicogenomic Applications: DrugMatrix and ToxFX. In Challenges and Advances in Computational Chemistry and Physics; Hong, H., Ed.; Springer: Berlin, Germany, 2019. [Google Scholar]
- Sard, H.; Kumaran, G.; Morency, C.; Roth, B.L.; Toth, B.A.; He, P.; Shuster, L. SAR of psilocybin analogs: Discovery of a selective 5-HT2C agonist. Bioorg. Med. Chem. Lett. 2005, 15, 4555–4559. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.C.; Jensen, N.H.; Kurome, T.; Kadari, S.; Manzano, M.L.; Malberg, J.E.; Caldarone, B.; Roth, B.L.; Kozikowski, A.P. Selective 5-hydroxytryptamine 2c receptor agonists derived from the lead compound tranylcypromine: Identification of drugs with antidepressant-like action. J. Med. Chem. 2009, 52, 1885–1902. [Google Scholar] [CrossRef] [Green Version]
- Rickli, A.; Moning, O.D.; Hoener, M.C.; Liechti, M.E. Receptor interaction profiles of novel psychoactive tryptamines compared with classic hallucinogens. Eur. Neuropsychopharmacol. 2016, 26, 1327–1337. [Google Scholar] [CrossRef] [Green Version]
- Tosh, D.K.; Janowsky, A.; Eshleman, A.J.; Warnick, E.; Gao, Z.G.; Chen, Z.; Gizewski, E.; Auchampach, J.A.; Salvemini, D.; Jacobson, K.A. Scaffold Repurposing of Nucleosides (Adenosine Receptor Agonists): Enhanced Activity at the Human Dopamine and Norepinephrine Sodium Symporters. J. Med. Chem. 2017, 60, 3109–3123. [Google Scholar] [CrossRef] [Green Version]
- Landagaray, E.; Ettaoussi, M.; Duroux, R.; Boutin, J.A.; Caignard, D.H.; Delagrange, P.; Melnyk, P.; Berthelot, P.; Yous, S. Melatonergic ligands: Design, synthesis and pharmacological evaluation of novel series of naphthofuranic derivatives. Eur. J. Med. Chem. 2016, 109, 360–370. [Google Scholar] [CrossRef]
- Vangveravong, S.; Kanthasamy, A.; Lucaites, V.L.; Nelson, D.L.; Nichols, D.E. Synthesis and serotonin receptor affinities of a series of trans-2- (indol-3-yl)cyclopropylamine derivatives. J. Med. Chem. 1998, 41, 4995–5001. [Google Scholar] [CrossRef]
- Bajusz, D.; Racz, A.; Heberger, K. Why is Tanimoto index an appropriate choice for fingerprint-based similarity calculations? J. Cheminform. 2015, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- InstantJChem version 15.3.30.0, licensed by ChemAxon. 2015. Available online: www.chemaxon.com (accessed on 31 August 2017).
- Sastry, M.; Lowrie, J.F.; Dixon, S.L.; Sherman, W. Large-scale systematic analysis of 2D fingerprint methods and parameters to improve virtual screening enrichments. J. Chem. Inf. Model. 2010, 50, 771–784. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Gardner, S.P.; Sutcliffe, M.J. An automated approach for defining core atoms and domains in an ensemble of NMR-derived protein structures. Protein Eng. 1997, 10, 737–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LigPrep, Schrödinger Release 2019-3; LLC: New York, NY, USA, 2019.
- Kimura, K.T.; Asada, H.; Inoue, A.; Kadji, F.M.N.; Im, D.; Mori, C.; Arakawa, T.; Hirata, K.; Nomura, Y.; Nomura, N.; et al. Structures of the 5-HT 2A receptor in complex with the antipsychotics risperidone and zotepine. Nat. Struct. Mol. Biol. 2019, 26, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; McCorvy, J.D.; Harpsøe, K.; Lansu, K.; Yuan, S.; Popov, P.; Qu, L.; Pu, M.; Che, T.; Nikolajsen, L.F.; et al. 5-HT2C Receptor Structures Reveal the Structural Basis of GPCR Polypharmacology. Cell 2018, 172, 719–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glide, Schrödinger Release 2019-3; LLC: New York, NY, USA, 2019.
- Deng, Z.; Chuaqui, C.; Singh, J. Structural interaction fingerprint (SIFt): A novel method for analyzing three-dimensional protein-ligand binding interactions. J. Med. Chem. 2004, 47, 337–344. [Google Scholar] [CrossRef]
- Mordalski, S.; Kosciolek, T.; Kristiansen, K.; Sylte, I.; Bojarski, A.J. Protein binding site analysis by means of structural interaction fingerprint patterns. Bioorg. Med. Chem. Lett. 2011, 21, 6816–6819. [Google Scholar] [CrossRef]
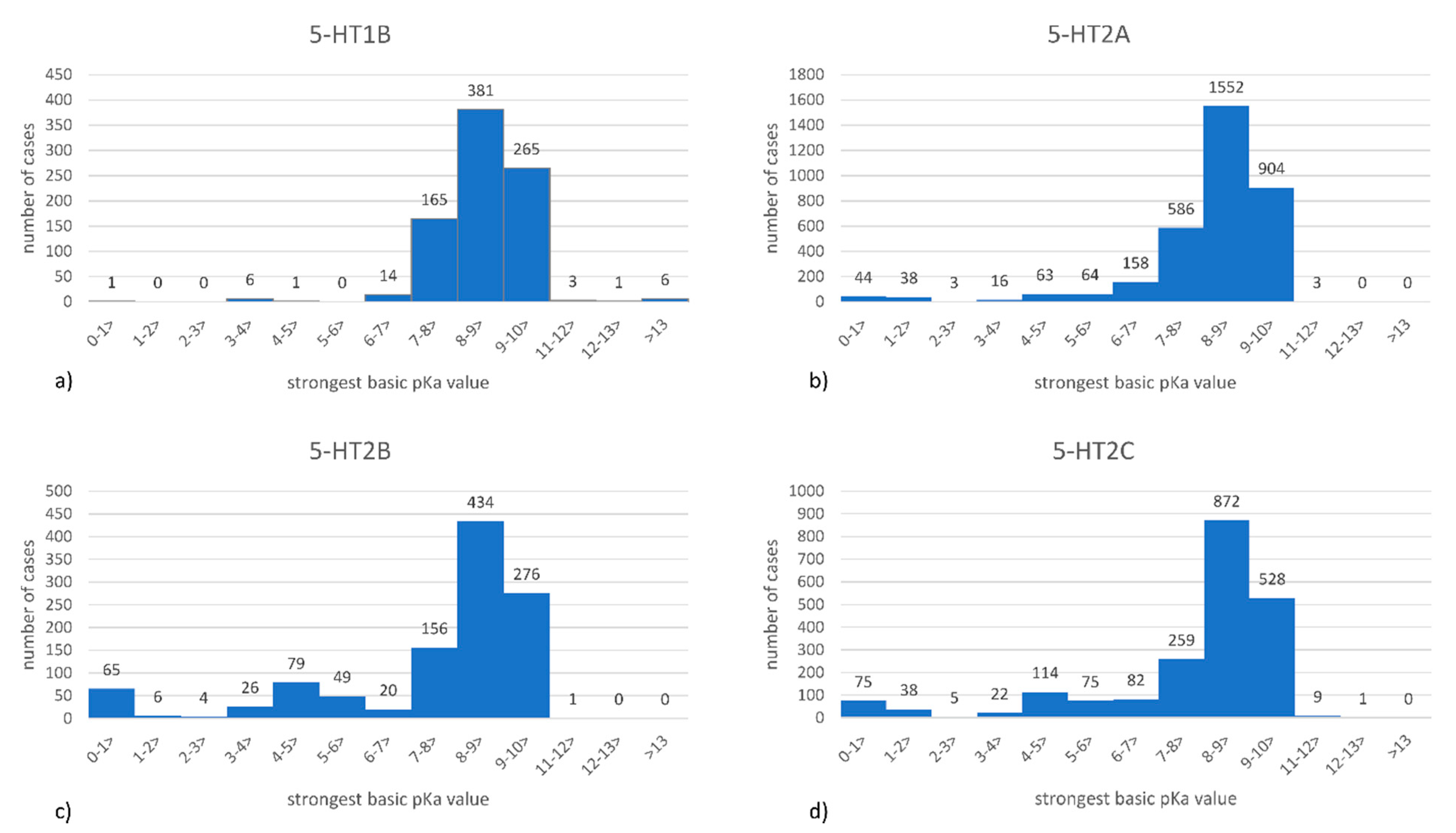



| 5-HT1B | 5-HT2A | 5-HT2B | 5-HT2C | |
|---|---|---|---|---|
| Low basicity | 8 (1.04%) | 228 (7.66%) | 229 (22.50%) | 329 (17.06%) |
| High basicity | 584 (75.75%) | 2005 (67.35%) | 613 (60.22%) | 1258 (65.25%) |
| Total number of ligands | 771 | 2977 | 1018 | 1928 |
| Ligand Structure | CHEMBLID | pKa | 5-HT2AR Ki [nM] | 5-HT2BR Ki [nM] | 5-HT2CR Ki [nM] |
|---|---|---|---|---|---|
 | CHEMBL54707 | 1.91 | 316 | 10 | 0.5 |
 | CHEMBL294030 | 1.21 | 100 | 50 | 1.26 |
 | CHEMBL240045 | No ionizable atom | 180 | 170 | 390 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podlewska, S.; Bugno, R.; Lacivita, E.; Leopoldo, M.; Bojarski, A.J.; Handzlik, J. Low Basicity as a Characteristic for Atypical Ligands of Serotonin Receptor 5-HT2. Int. J. Mol. Sci. 2021, 22, 1035. https://doi.org/10.3390/ijms22031035
Podlewska S, Bugno R, Lacivita E, Leopoldo M, Bojarski AJ, Handzlik J. Low Basicity as a Characteristic for Atypical Ligands of Serotonin Receptor 5-HT2. International Journal of Molecular Sciences. 2021; 22(3):1035. https://doi.org/10.3390/ijms22031035
Chicago/Turabian StylePodlewska, Sabina, Ryszard Bugno, Enza Lacivita, Marcello Leopoldo, Andrzej J. Bojarski, and Jadwiga Handzlik. 2021. "Low Basicity as a Characteristic for Atypical Ligands of Serotonin Receptor 5-HT2" International Journal of Molecular Sciences 22, no. 3: 1035. https://doi.org/10.3390/ijms22031035
APA StylePodlewska, S., Bugno, R., Lacivita, E., Leopoldo, M., Bojarski, A. J., & Handzlik, J. (2021). Low Basicity as a Characteristic for Atypical Ligands of Serotonin Receptor 5-HT2. International Journal of Molecular Sciences, 22(3), 1035. https://doi.org/10.3390/ijms22031035