1. Introduction
Mesenchymal Stromal Cells (MSC) are multipotent, self-renewing cells which have been shown to exhibit immuno-modulatory, anti-inflammatory and regenerative capacities at sites of inflammation [
1,
2,
3]. MSCs can be easily isolated from various types of tissues, such as the bone marrow, the umbilical cord or adipose tissue, and can furthermore be differentiated into osteoblasts, adipocytes and chondrocytes in vitro [
4]. Therefore, MSCs have been used as an experimental therapeutic agent in graft-versus-host disease (GVHD) [
5] and Crohn’s disease [
6]. Reported safety properties of MSCs have initiated interest on their administration at or around the time of solid organ transplantation (SOT), with an aim to alleviate ischemia reperfusion injury as well as the risk for subsequent rejection episodes [
7,
8,
9]. Although it was previously assumed that MSCs act by differentiating into functional cells [
10], evidence suggests that MSCs are more likely to act in a paracrine manner [
11]. Via the secretion of cytokines, growth factors and prostaglandins with immune-modulatory and regenerative function, MSCs may target resident cells. MSCs secrete a range of anti-inflammatory factors, including Interleukin-10 (IL-10), Transforming growth factor-beta (TGF-beta), hepatocyte growth factor (HGF), nitric oxide (NO), Histocompatibility antigen-G (HLA-G) and Prostaglandin E2 (PGE2), that are all indicated to play some role in their immunomodulatory effect [
9,
12,
13]. As MSCs are a mixed population of cells, there may be subtypes of cells with enhanced immunosuppressive capacity [
14]. Several reports have suggested ‘MSC-preconditioned’ macrophage activation to be partly responsible for the beneficial effects [
15,
16,
17]. Whether these result from direct cell-cell interactions in the sense of an MSC trans-differentiation at sites of damage, or from paracrine mechanisms, remains unclear.
In SOT, macrophages, amongst other cells, are key initiators of the early innate immune response contributing to ischemia reperfusion injury (IRI), by sending inflammatory signals to other effector cells as well as by a release of soluble mediators. Production of cytokines and chemokines then contributes to an influx of leukocytes, and further promotion of inflammation. Equally however, following activation, macrophages and dendritic cell subpopulations subsequently contribute to the resolution of injury [
18]. Depending on the microenvironmental signals, two major phenotypical/functional sub-populations have been identified. The M1 macrophage represents an activated/inflammatory phenotype, whereas the M2 macrophage represents an activated/regenerative phenotype [
19]. Therefore, and due to the fact that in vitro macrophage models are very well established, we aimed to investigate the effects of supernatants from different MSC phenotypes on macrophages in vitro.
Several research groups around the world are investigating the effects of MSCs as a therapeutic agent prior to or after SOT, using systemic infusion techniques, or ex-vivo organ machine perfusion techniques in order to precondition organs prior to transplantation. In animal studies, with an aim to trace cells after experiments, often MSCs from genetically modified donors are used, e.g., transgenic animals positive for the expression of green fluorescent protein [
20], which for us, raised the question whether any genetic modification, or even a variation in age of donors would have an influence on the immunomodulatory capacities of extracted MSCs. So far, these genetic determinants have not been investigated. We hypothesized, that genetic modification of donor rats for MSCs, as well as the donor age, might have an influence on the immunosuppressive capacities of MSCs. Therefore, our objective was to conduct an in vitro study to investigate paracrine effects of supernatants from MSCs extracted from genetically modified donors and age groups on BMDMs with or without prior inflammatory stimuli.
3. Discussion
The immunosuppressive properties of MSCs have been extensively studied both in vitro and in animal models of immune-mediated disorders. Clinical trials are currently underway in which MSCs are employed to treat various human immunological diseases as well as inflammatory processes, e.g., ischemia reperfusion injury [
21]. The molecular mechanisms leading to immunomodulatory and anti-inflammatory effects of MSCs on different immune cells are still unclear. In that context, ‘MSC- preconditioned’ macrophage activation has been proposed to play a role by some groups in the past [
15,
16,
22]. In order to investigate mechanisms of actions of MSCs, research groups focus on labelling of MSCs prior to their use in in-vivo experiments. Methods of labelling include transfection techniques applied on the cells, or use of transgenic donor animals, classically positive for green fluorescence [
20]. Whether genome interfering labelling methods could alter the function, e.g., immunomodulatory capacities of extracted MSCs, has however not been investigated so far. Indeed, the creators of the
GFP+ transgenic WKY rats used for our experiments, cannot exclude an effect of the genetic manipulation on splicing machineries within the genome, and hence, functional alterations further downstream [
20].
A study conducted by Behmoaras et al. in 2014 found, that soluble factors present in supernatants of mesangial cells (MC) from
WKY rats could differentiate BMDM into active/inflammatory M1, or active/regenerative M2 phenotypes, depending on the genetic background [
23] of the MCs used. MCs and MSCs are of similar origin, therefore we hypothesized to potentially find similar differences in MSCs extracted from donors with a genetic modification. Furthermore, we hypothesized, that not only a genetic modification of MSC donors might influence the effect of MSCs on BMDMs in vitro, but also the age of respective donors. Studies have demonstrated that properties and functionalities of MSCs can be influenced by intrinsic factors including aging and that MSCs extracted from young and elder individuals have diverse properties [
24,
25]. In regards to MSCs as a potential therapeutic agent to alleviate ischemia-reperfusion injury in solid organ transplantation, an aim would be to identify the MSC geno- and phenotype with the most promising anti-inflammatory potential. To our knowledge this is the first study investigating MSC lineage and donor animal age.
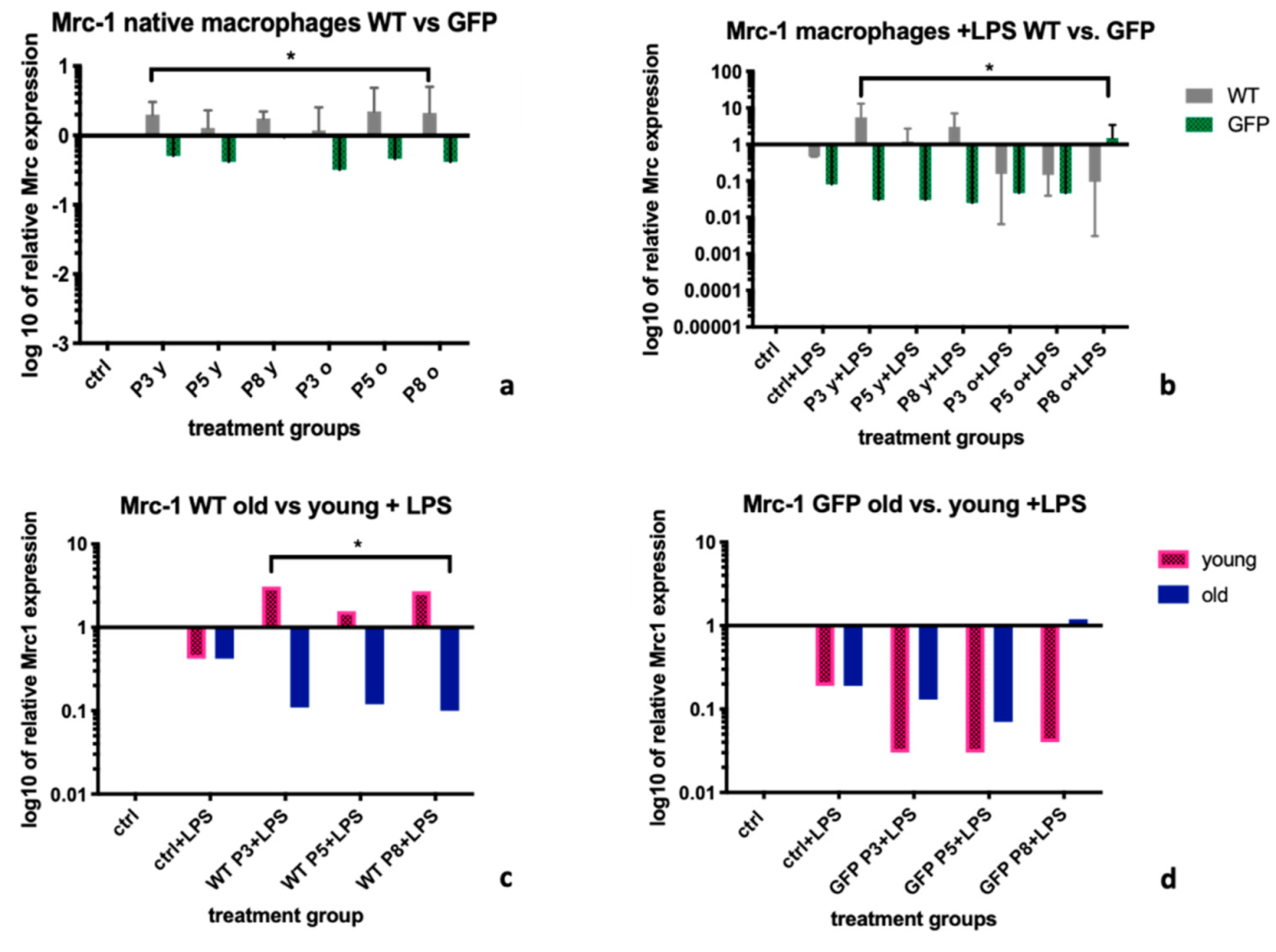
Mrc-1 in unstimulated BMDMs was up-regulated after treatment with supernatants from MSC-wt cells, not from MSC-GFP+ cells (
Figure 1a). After LPS stimulation, the Mrc-1 downregulation within the control groups was only reversed by supernatants from MSC-wt cells, not from MSC-GFP+ cells (
Figure 1b). We furthermore found this effect to be stronger after treatment of LPS stimulated BMDM with supernatants from young MSC-wt cells (
Figure 1c). Therefore, in regard to Mrc-1 as marker for the polarization of BMDM, and Mrc-1 upregulation to indicate presence of the more regenerative M2 macrophage phenotype, we found supernatants from young MSC-wt donors to have the most anti-inflammatory effect.
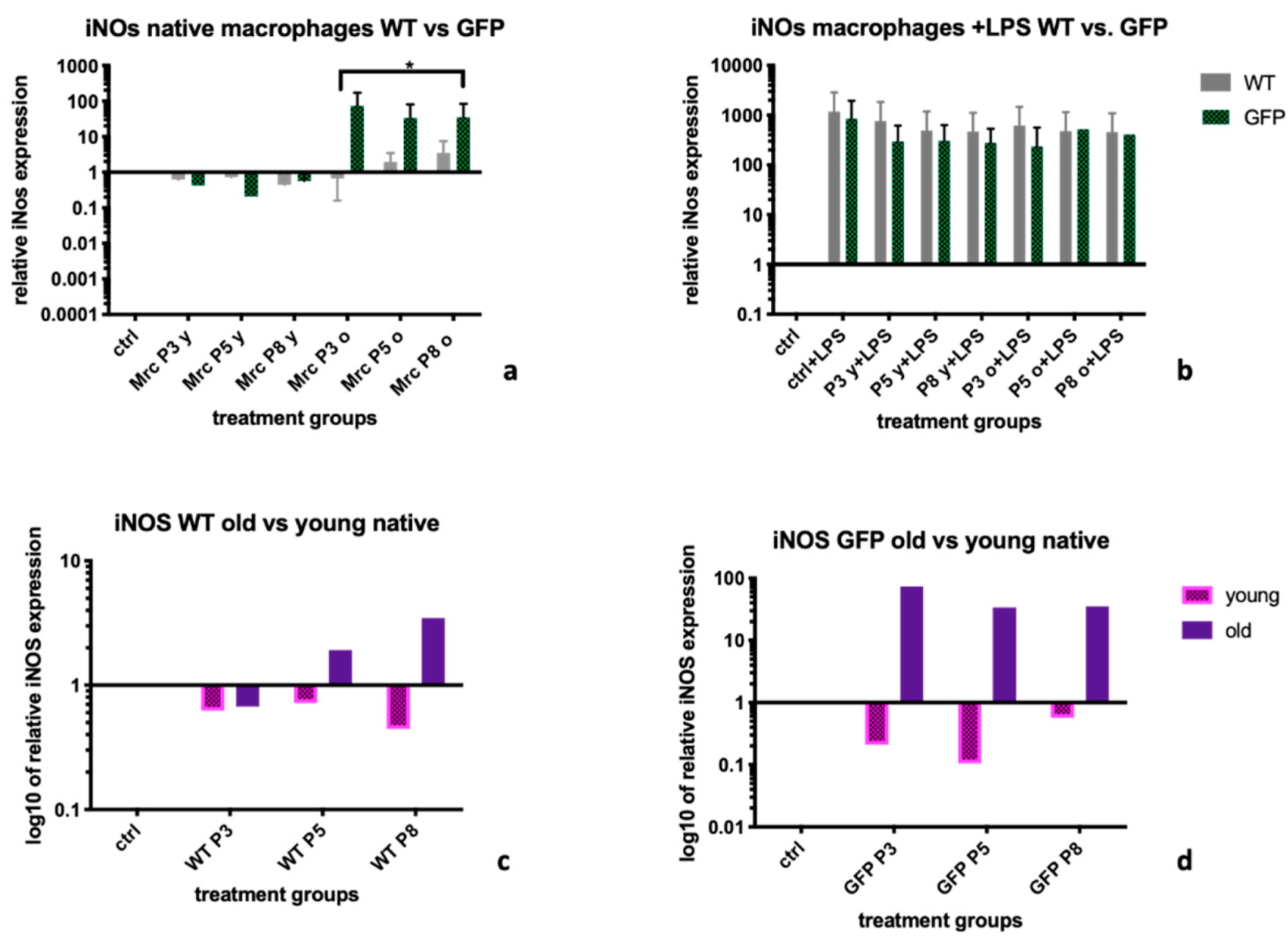
For iNOs, a marker for a more pro-inflammatory BMDM phenotype M1, we observed a better down regulation after treatment with supernatants from MSC-wt than from MSC-GFP+ cells, which was significant for supernatants from old donors (
Figure 2a). Sub-group analysis did however not show significant differences between effects from supernatants from young cells versus old cells, for neither of the two, MSC-wt or MSC-GFP+ genotypes of MSCs (
Figure 2c,d). Interestingly, after stimulation of BMDM with LPS, none of the supernatant treatments led to an efficient downregulation of iNOS (
Figure 2b). We suggest that the capacity for anti-inflammatory effects by chemokines in supernatants might also be influenced by the inflammatory milieu the BMDM are in, and that multiple factors contribute to this. Without cells present in the supernatants, the anti-inflammatory capacity might be too weak to counteract in a pro-inflammatory environment.
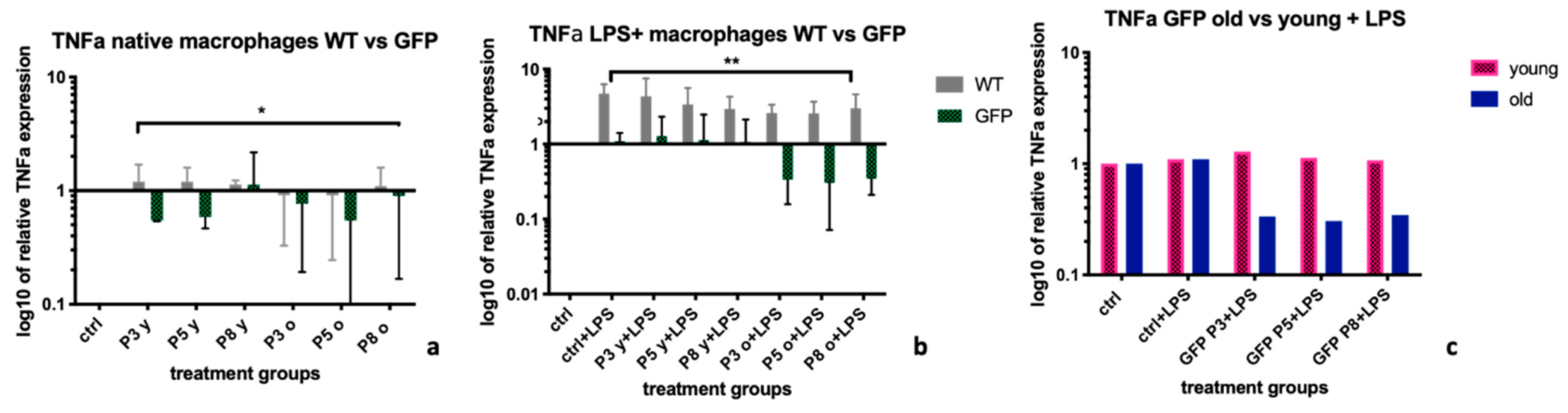
TNFα is a key feature of activated/inflammatory M1 macrophages [
26,
27]. Contrary to findings for iNOS, supernatants from MSC-GFP+ cells, led to significantly better suppression of TNFα expression levels, both in native (
Figure 3a) and LPS stimulated macrophages (
Figure 3b), than supernatants from MSC-wt cells. The best down-regulation was achieved by supernatants from MSC-GFP+ old cells (
Figure 3c). This confirms our theory that supernatants from cells of genetically modified donors can produce different chemokine patterns and therefore, regulate different cytokines in different ways. It might also be dependent on the microenvironment and the activation state of the macrophage, which effect will be observed.
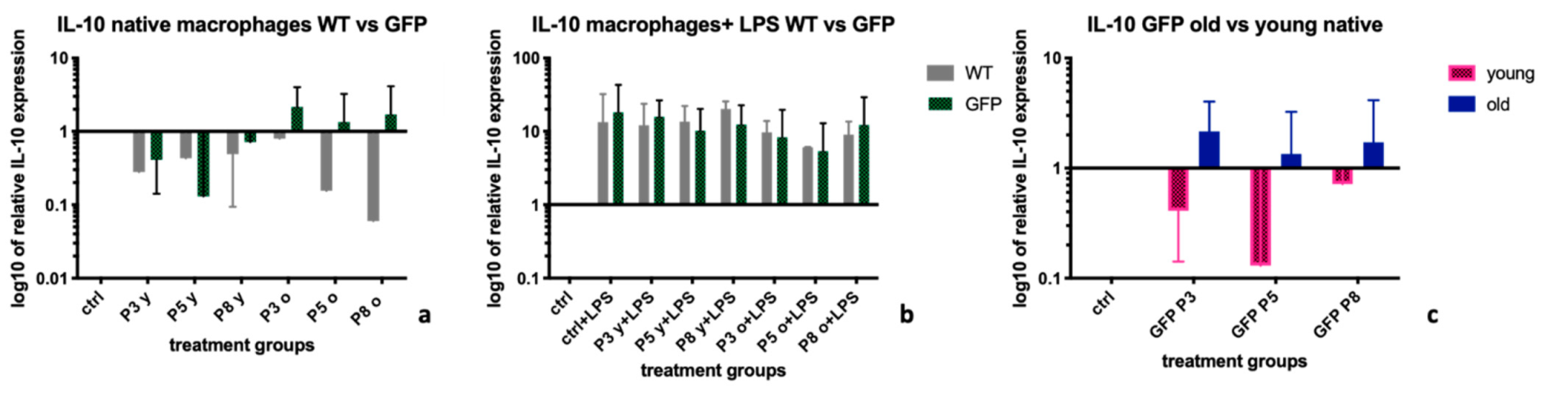
IL-10, as a marker for the anti-inflammatory M2 macrophage phenotype [
28], was more up-regulated by supernatants from MSC-GFP+ cells in untreated macrophages, than by supernatants from MSC-wt cells (
Figure 4a), without reaching statistical significance. Especially supernatants from old MSC-GFP cells seemed to up-regulate
IL-10 expression levels, however in a sub-group analysis between supernatants from young and old MSC-GFP+ cells, no significant difference was detected (
Figure 4c). Also after LPS stimulation of macrophages, we did not observe any essential differences in
IL-10 expression levels
(Figure 4b).
Limitations of our study are that the supernatant transfer experiment was repeated three times, respective PCRs were repeated twice, and each sample was run in duplicates. Therefore, we acknowledge the statistical challenges and the resulting range of results, with sometimes high interquartile ranges. However, we think that the number of repetitions of the experiment as well as of PCR reactions, can also be acknowledged as a strength of the study.
In summary, we have observed similar results as described by Behmoaras et al. for mesangial cells. In our case, different paracrine effects were observed for supernatants from MSCs from a genetically modified donor rat, expressing GFP under the control of elongation factor 1α, as well as from MSCs of rats of different age groups. Different passages showed similar results. Looking at Mrc-1 and iNOS regulations to start with, it seemed like MSC-wt cells were more promising as inhibitor cells of inflammatory cascades in a paracrine manner. However looking at TNFα and IL-10 expression levels, supernatants from MSC-GFP+ cells seemed to be more promising in regards to promotion of anti-inflammatory cascades within macrophages.
For future experiments, it would be sensible to measure the exact concentrations of cytokines present in the supernatants, and also, to perform a similar experiment on a direct cell-cell interaction level in parallel. This might help to further understand the mechanisms of action of MSCs. Nevertheless, different phenotypes of MSCs depending on their age and the presence of genetic modifications of donor animals should be considered in in vivo studies.
4. Materials and Methods
All studies were performed with Ethical approval and approval under the Animal Scientific Procedures Act (1986). Experiments were performed under Project Licence Number PB1C4696D, granted by the Home Office, UK.
4.1. Extraction and Culture of Bone Marrow Derived Mesenchymal Stromal Cells
MSCs were extracted from the bone marrow of
wild-type Wystar Kyoto rats (WKY-wt) and from
Wystar Kyoto rats positive for the expression of green fluorescent protein
(WKY-GFP+). This specific rat had been genetically modified to express GFP under the control of elongation factor 1α, by Dr. Anna Garcia-Diaz et al [
20]. Donor rats were either 6 weeks (young), or 6 months (old) of age, respectively. After sacrifice of the donor rats according to the Schedule 1 protocol, femurs and tibias were retrieved and bone marrow was flushed into fresh Falcon tubes using sterile Hank’s Balanced Salt Solution (HBSS; Gibco, Thermo Fisher Scientific, Waltham, MA, USA) at 4 °C. Cell suspensions were washed twice with HBSS, supernatants were discarded and pellets containing cells were resuspended in 1 mL of MesenCult
TM MSC Basal Medium (StemCell Technologies, Cambridge, UK) before being added into flasks containing 24 mL full MSC culture medium containing MSC stimulatory supplement and 0.5% Penicillin/Streptomycin (Invitrogen, Carlsbad, CA, USA). At a confluence of 80%, cells were trypsinized, counted and split.
4.2. Confirmation of MSC Phenotype
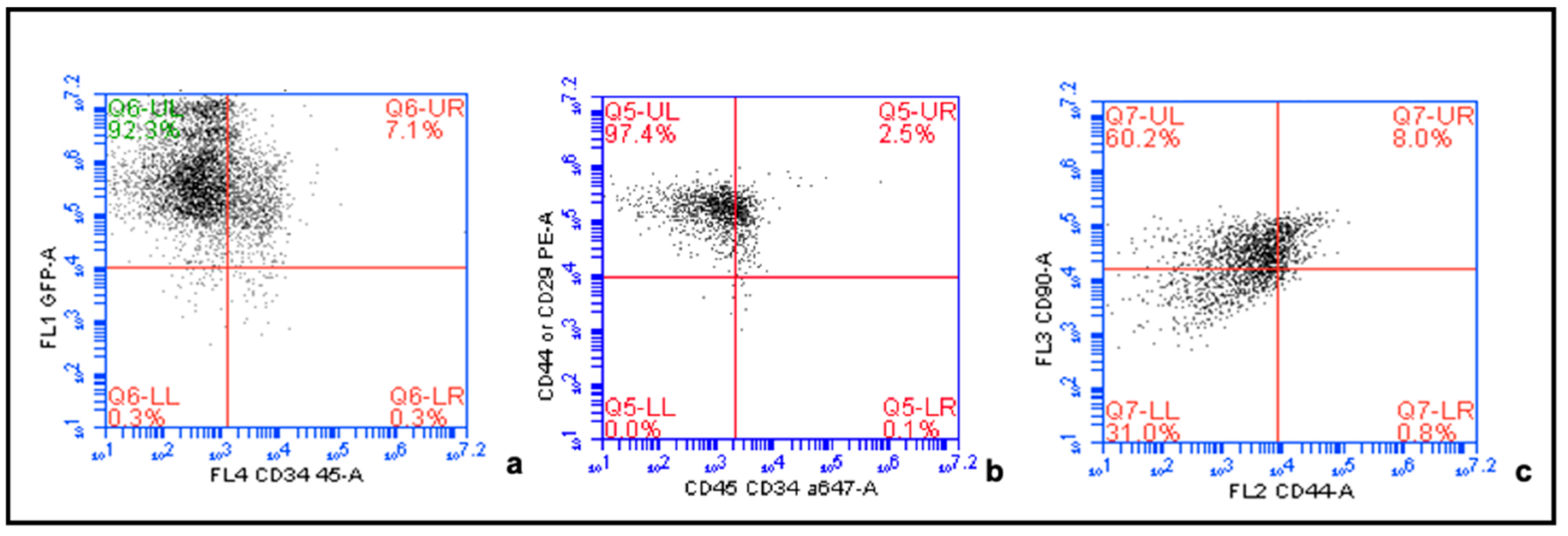
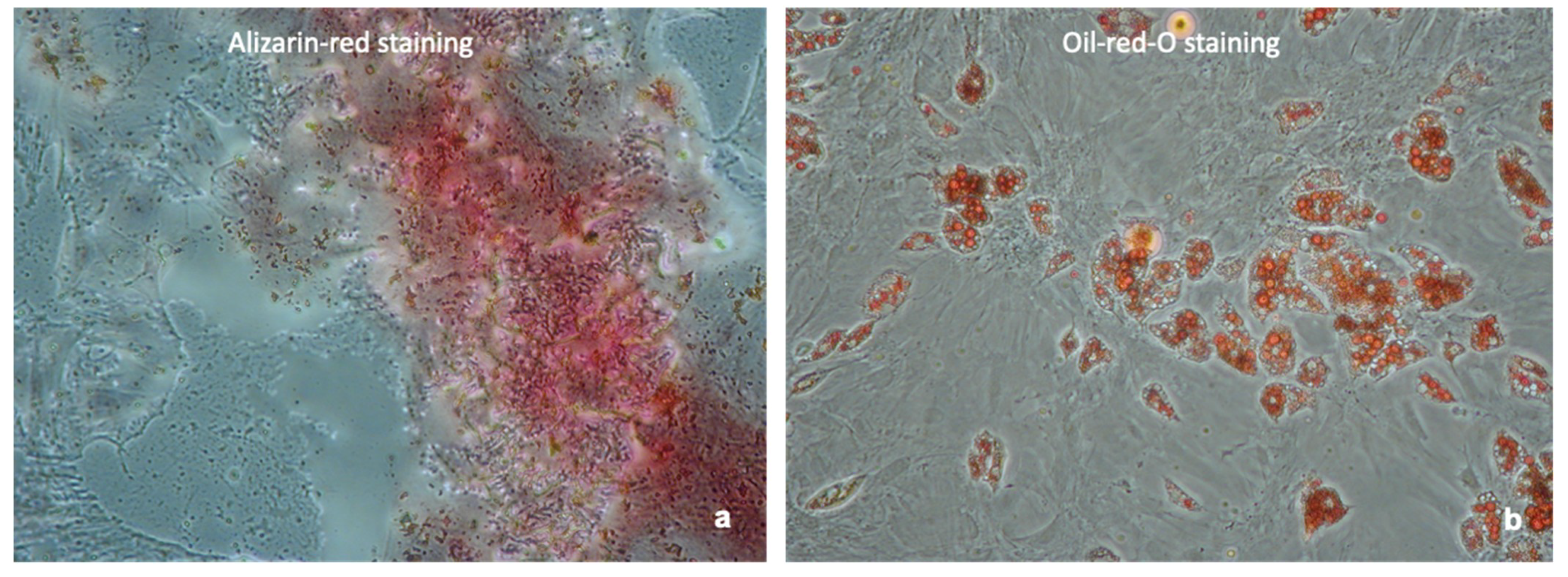
MSC phenotype for all MSC cultures was proven by demonstration of their differentiation capacities into adipocytes and osteocytes. Rat MSC Adipogenic Bullet kit medium (Lonza, Basel, Switzerland) was added to the cells for adipogenic induction, and rat Osteogenic Bullet kit medium (Lonza) for osteogenic differentiation. Differentiation of the cells into adipocytes was confirmed by Oil-Red-O staining and formation of bone matrix was confirmed by Alizarin Red staining. Furthermore, MSC phenotype was confirmed by Flow cytometry. Cells were shown to express the cell surface markers CD29 (antibody: PE anti-mouse/rat CD29, clone: HMβ1-1, Bio Legend Cat: 102207, Conc: 0.2 mg/mL), CD90 (antibody: PerCP anti-rat CD90/mouse, CD90.1 (Thy-1.1) clone: OX-7, BioLegend Cat: 202512; Lot: B171081, Conc: 0.2 mg/mL), and CD44 (antibody: RPE mouse anti-rat CD44, clone OX-50, Bio Rad Cat: MCA643PE), and to lack the expression of CD34 (antibody: Alexa Fluor 647 anti-mouse CD34, clone: ICO115, Novus Biologicals Cat: NBP2-33076AF647; Conc: 0.75 mg/mL) and CD45 (antibody: Alexa Fluor 647 anti-rat CD45, clone: OX-1, BioLegend Cat: 202212; Conc: 0.5 mg/mL) for MSC phenotyping (
Figure A2).
4.3. MSC Supernatants
For the purpose of supernatant transfer experiments, for each group and passage of MSC cultures (P1-P10), triplicates of 2.5 × 105 cells were placed into a 24 well plate in 500 µL of MesenCultTM full culture medium (StemCell, Catalogue number Catalog #05513) for 24 h. Thereafter, supernatants from all the wells were collected and stored on −80 °C until used for the experiments. The following groups of supernatants were obtained for experiments:
MSC-wt young, P1-P10: supernatants from MSCs in culture, extracted from a 6 week old WKY-wt rat, passages 1–10.
MSC-GFP+ young, P1-P10: supernatants from MSCs in culture, extracted from a 6 week old WKY-GFP+ rat, passages 1–10.
MSC-wt old, P1-P10: supernatants from MSCs in culture, extracted from a 6 month old WKY-wt rat, passages 1–10.
MSC-GFP+ old, P1-P10: supernatants from MSCs in culture, extracted from a 6 month old WKY-GFP+ rat, passages 1–10.
4.4. Extraction of Bone Marrow Derived Macrophages
L929 conditioned culture medium for macrophage culture was prepared and filtered with a sterile filter. Bone marrow derived macrophages were obtained from a WKY-wt rat. The animal was sacrificed according to the Schedule 1 protocol. Femurs and the tibias were retrieved, and bone marrow was flushed into a Falcon tube in a laminar flow hood. The content was spun for 5 min at 1500 rpm and a temperature of 4 °C. The supernatant was discarded and the pellet containing the cells was resuspended in 10 mL of HBSS. Red blood cell lysis was performed. Cells were spun again for 5 min at 1500 rpm, 4 °C, the supernatant was discarded and cells were resuspended in 1 mL of full culture media, which per 375 mL contained: 240 mL DMEM (+L-glu), 3 g of Hepes 25 mM, 125 mL L929 conditioned media (25%) and 10 mL Pen/Strep (100 U/mL, 100 ug/mL). The cells were dispensed equally into 3 large petri dishes containing 25 mL BMDM full culture medium and cultured for 5 days in the 37 °C, 5%CO2 incubator. After 5 days, BMDM full culture medium was removed and dissociation buffer was added to cover the surface of the cells before incubating for 10–20 min at 37 °C, 5%CO2. Cells were then collected and spun down for 5 min at 1500 rpm. The supernatants were discarded and 1 mL of BMDM full culture medium was added, respectively, to resuspend the cells. Cells were counted with the use of a haemocytometer and 2.5 × 105 cells per well were seeded into 24 well plates in 400 µL of BMDM full culture medium, respectively. Cells were incubated over night to adhere to the plastic again.
4.5. Supernatant Transfer Experiments
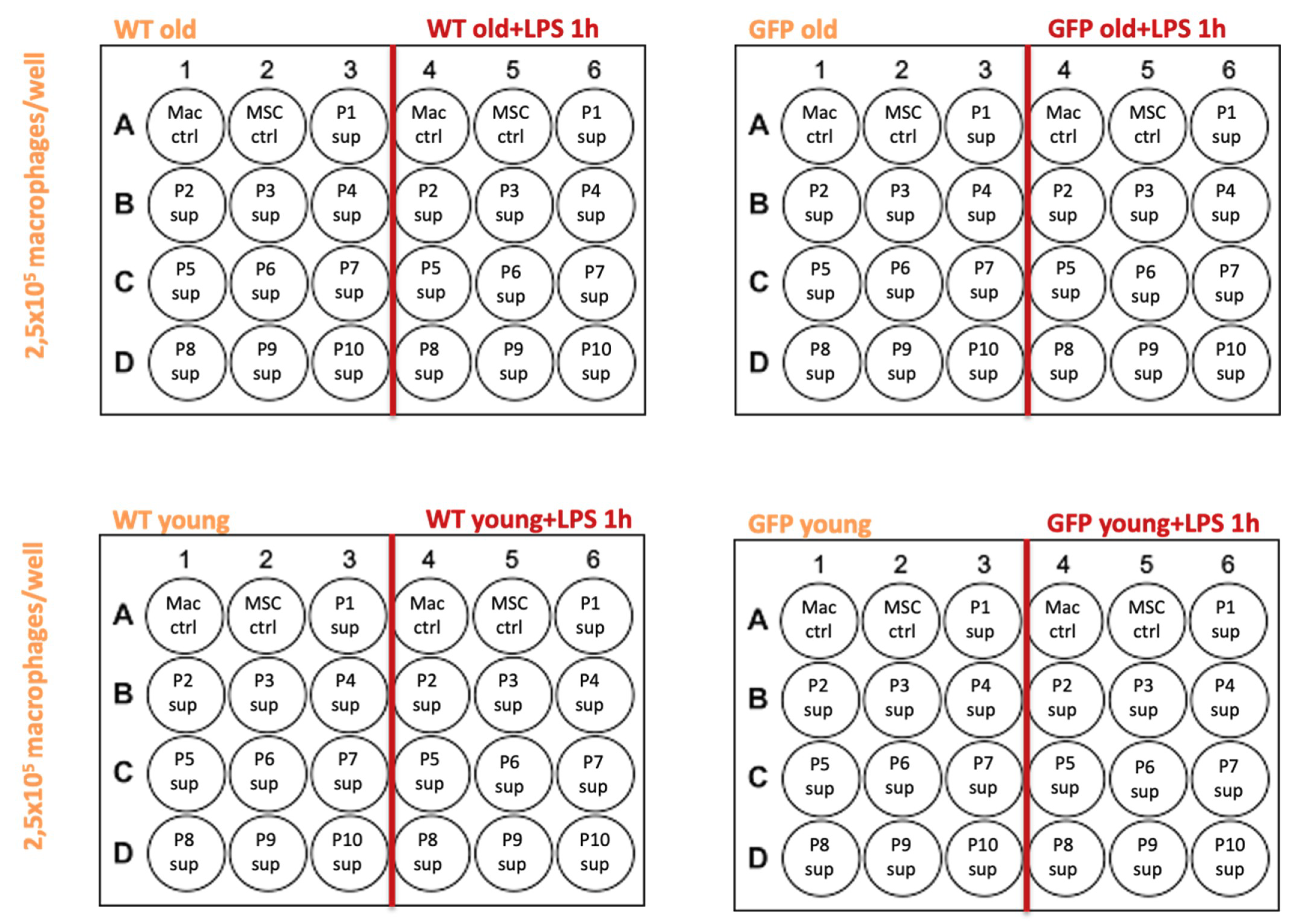
After confirmation of the viability of BMDMs in all wells of the 24 well plates by light microscopy, wells were washed with HBSS. Macrophage control samples (Mac ctrl) were treated with 400 µL of the DMEM based medium alone. MSC control samples (Mac MSC ctrl) contained 150 µL DMEM based medium + 250 µL plain MSC growth medium which was freshly prepared in order to rule out that any effect could come from any of the ingredients of the MSC growth medium (MesenCult proliferation kit, Stem Cell Technologies, UK). Cells in treated wells within columns 4–6 of a 24 well plate, underwent treatment with LPS for 1 h, respectively. Wells in columns 1–3 underwent change of media only. Macrophages were treated with 250 µL of the supernatants from MSCs P1-P10. Each well contained 150 µL DMEM based medium and 250 µL of MSC supernatant.
Figure A3 shows a scheme of the experiment, highlighting respective treatment groups within the 24 well plates. The plates were left in an incubator at 37 °C and 5%CO
2 over-night. Thereafter the macrophages were washed with HBSS and the cells from each well were lysed using 250 µL of Trizol reagent for RNA extraction.
4.6. qRT-PCR
For each well, RNA extraction was performed with the TRIzol method (Thermo Fisher Scientific, UK). DNA was digested using DNase I (Sigma-Aldrich, UK). The concentration and purity of RNA samples was assessed using the A260:A280 ratio and A260:A230 ratio, respectively using a Nano drop 2000c Spectrophotometer (Thermo Fisher Scientific). Quantitative real time PCR (qRT-PCR) was performed using the iScript Sybr Green Supermix (BioRad, Hertfordshire, UK). Optimal Primer combinations and PCR programmes were tested before performance of the actual qRT-PCR. PCRs were performed in duplicates in 96-well PCR plates (Thermo Fisher Scientific, UK) on an Eppendorf Mastercycler® RealPlex. PCRs were performed with the following protocol:
3 min at 95 °C (hot start), followed by 40 cycles of the following:
15 s at 95 °C denaturation step
20 s at 60.4 °C primer annealing step
40 s at 72 °C elongation step, followed by
Melting curve
Hold at 4 °C
The housekeeping gene
Hypoxanthine-Guanine Phosphoribosyltransferase (HPRT) was used as positive control. Primer pair sequences for these genes as well as their product lengths are shown in
Table 1. All primer pairs were designed using Primer blast (NCBI) and were synthesised by Sigma-Aldrich. The relative quantification of a target gene in the PCR reaction compared to the control was calculated using the comparative Ct method ΔΔCt. Specific primer combinations for qRT-PCR are shown in
Table 1.
4.7. MTT Viability Assay
The MTT viability Assay was performed on macrophages, mimicking circumstances of respective supernatant transfer experiments, in order to investigate whether the treatment led to cell death that could cause altered results. The MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) reagent was diluted 1:10 in culture media. In a 96-well plate, rat bone marrow derived macrophages (BMDM) underwent the same respective treatments as macrophages in 24-well plates for previously described experiments. Supernatant were removed from cells without disturbing the cell sheet and 100μL MTT solution per well were added. Cells were incubated overnight at 37 °C in the incubator. Subsequently an equal volume of SDS MTT solution (10% SDS, 0.01M HCl) was added and incubated at 37 °C for at least 3 h or overnight. The plate was read on an ELISA plate reader at 592 nm and ODs were compared.
4.8. Statistical Evaluation
The supernatant transfer experiment was repeated three times and qRT-PCRs were performed in duplicates and repeated twice, respectively. Relative mRNA expression levels of selected cytokines were calculated using the ΔΔCt method and using the ubiquitously expressed
Hypoxanthine-Guanine Phosphoribosyltransferase (HPRT) as a housekeeping gene. Relative expressions were calculated in comparison to the respective control samples (e.g., macrophage control, LPS control, macrophage MSC control- see
Figure A4). Mean values from the three experiments were calculated, respectively, and statistical analyses between groups were performed using Mann Whitney U test as well as ANOVA. In order to simplify the results, for subgroup analyses, passages were plotted together to result in one mean value, respectively (this resulted in columns without markers for a standard deviation as only one mean value was used for analysis). Statistical analyses were conducted for all subgroups but in this manuscript, with consciousness to an otherwise overwhelming set of data, only the most interesting/significant findings are demonstrated and visualized by graphs. GraphPad Prism was used to perform all statistical analyses.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}