
Effective Activation by Kynurenic Acid and Its Aminoalkylated Derivatives on M-Type K+ Current

 ,
, 
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
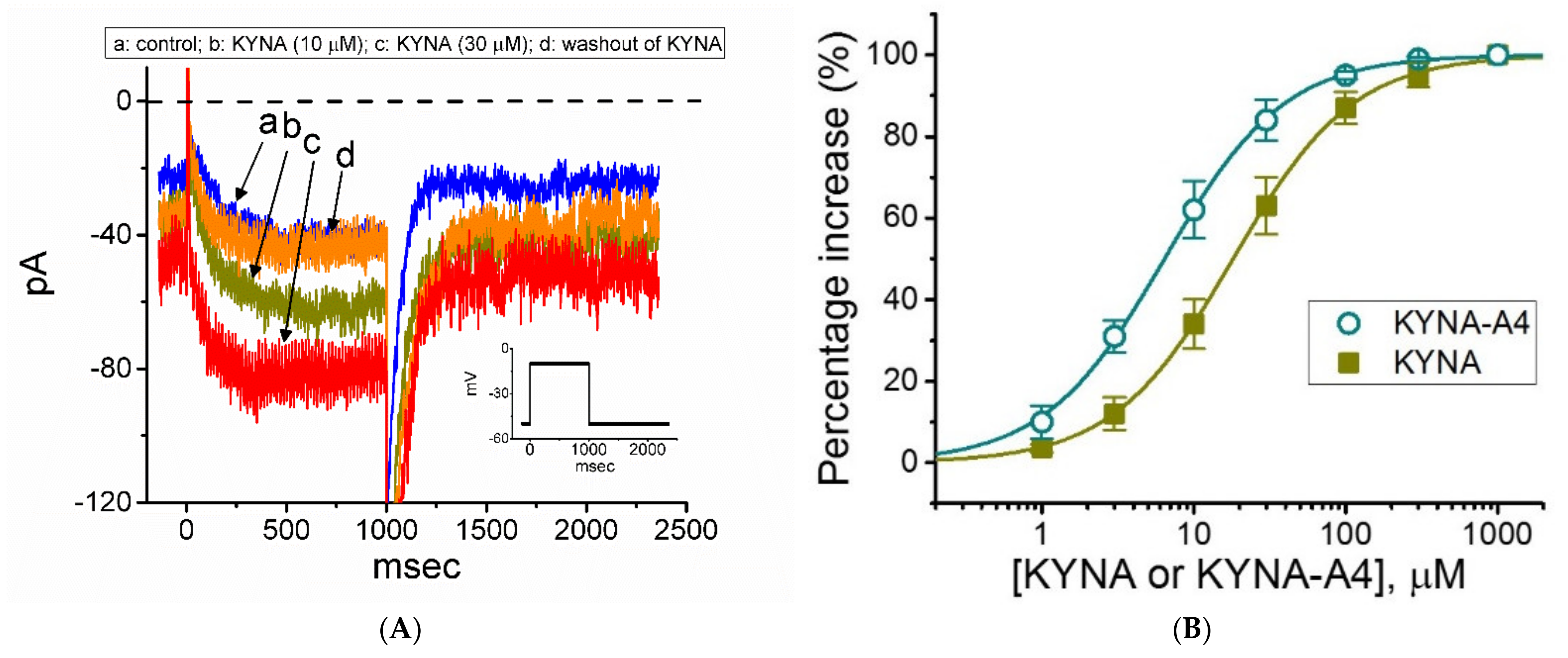
2.1. Effect of KYNA and KYNA Derivatives (i.e., KYNA-M1 and KYNA-A4) on M-Type K+ Current (IK(M)) in GH3 Cells
2.2. Steady-State Activation Curve of IK(M) in the Absence and Presence of KYNA or KYNA-A4 in GH3 Cells
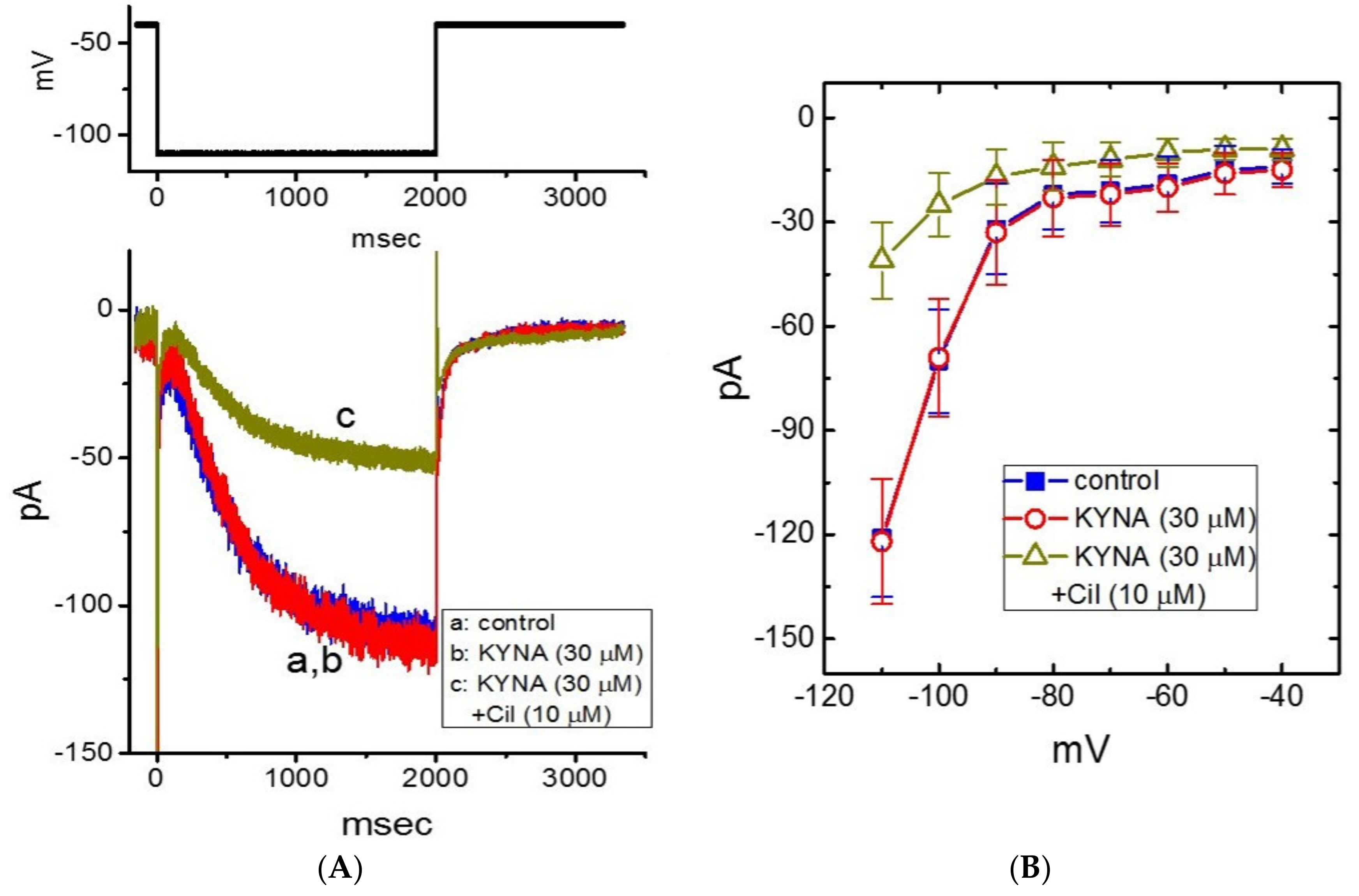
2.3. Effect of KYNA on Voltage-Dependent Hysteresis of IK(M) Elicited by Long Isosceles-Triangular Ramp Pulse
2.4. Effect of KYNA on M-Type K+ Channel (KM) Channels Recorded from GH3 Cells
2.5. Effect KYNA on Delayed-Rectified K+ Current (IK(DR)) in Pituitary Tumor (GH3) Cells
2.6. Lack of KYNA on Erg-Mediated K+ Current (IK(erg))
2.7. Inability of KYNA to Modify Hyperpolarization-Activated Cation Current (Ih)
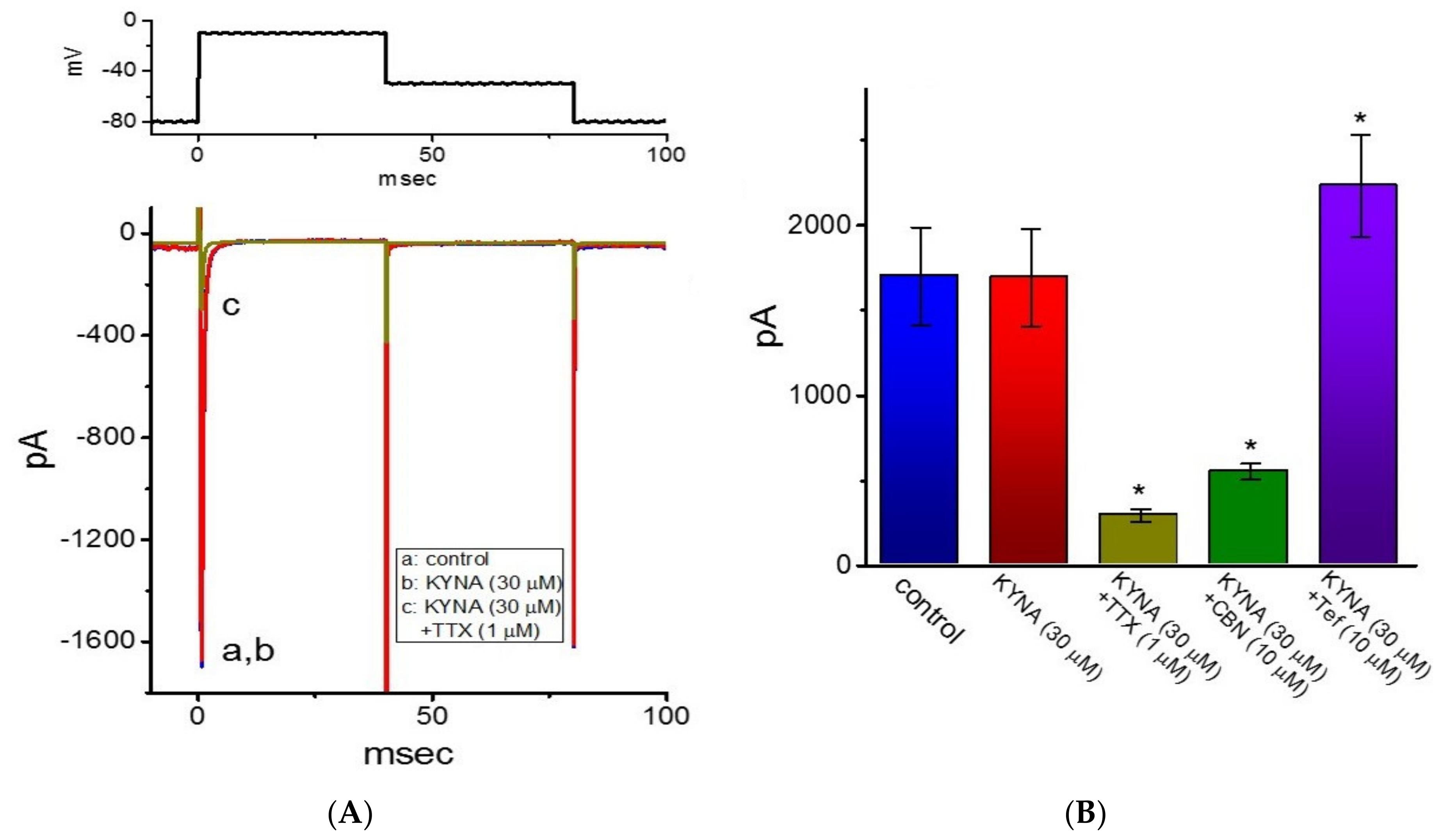
2.8. Ineffectiveness of KYNA in Modifying Voltage-Gated Na+ Current (INa) in GH3 Cells
2.9. Effect of KYNA or KYNA-A4 on Spontaneous Action Potentials (APs) Recorded from GH3 Cells
2.10. Effect of KYNA and KYNA Plus Linopirdine on IK(M) Measured from Hippocampal mHippoE-14 Neurons
3. Discussion
4. Materials and Methods
4.1. Drugs, Chemicals, and Solutions Used in This Study
4.2. Cell Preparations
4.3. Electrophysiological Measurements
4.4. Data Recordings
4.5. Data Analyses
4.6. Analyses of Single M-Type K+ (KM) Channels
4.7. Statistical Analyses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AP | Action potential |
| EC50 | The concentration required for 50% stimulation |
| HCN channel | Hyperpolarization-activated cyclic nucleotide-gated ion channel |
| Ih | Hyperpolarization-activated cation current |
| IK(DR) | Delayed-rectifier K+ current |
| I-V | Current versus voltage |
| IK(erg) | Erg-mediated K+ current |
| IK(M) | M-type K+ current |
| INa | Voltage-gated Na+ current |
| KM channel | M-type K+ channel |
| KYNA | Kynurenic acid |
| KYNA-M1 | 3-(morpholinomethyl)-4-oxo-1,4-dihydroquinoline-2-carboxylic acid |
| KYNA-A4 | N-(2-(dimethylamino)ethyl)-3-(morpholinomethyl)-4-oxo-1,4-dihydroquinoline-2-carboxamide |
| NMDAR | N-methyl-D-aspartate receptor |
| SEM | Standard error of mean |
| TEA | Tetraethylammonium chloride |
| TTX | Tetrodotoxin |
References
- Hilmas, C.; Pereira, E.F.; Alkondon, M.; Rassoulpour, A.; Schwarcz, R.; Albuquerque, E.X. The brain metabolite kynurenic acid inhibits alpha7 nicotinic receptor activity and increases non-alpha7 nicotinic receptor expression: Physiopathological implications. J. Neurosci. 2001, 21, 7463–7473. [Google Scholar] [CrossRef] [PubMed]
- Erhardt, S.; Olsson, S.K.; Engberg, G. Pharmacological manipulation of kynurenic acid: Potential in the treatment of psychiatric disorders. CNS Drugs 2009, 23, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Moroni, F.; Cozzi, A.; Sili, M.; Mannaioni, G. Kynurenic acid: A metabolite with multiple actions and multiple targets in brain and periphery. J. Neural Transm. 2012, 119, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Stone, T.W. Does kynurenic acid act on nicotinic receptors? An assessment of the evidence. J. Neurochem. 2020, 152, 627–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erhardt, S.; Schwieler, L.; Nilsson, L.; Linderholm, K.; Engberg, G. The kynurenic acid hypothesis of schizophrenia. Physiol. Behav. 2007, 92, 203–209. [Google Scholar] [CrossRef]
- Chen, Y.; Meininger, V.; Guillemin, G.J. Recent advances in the treatment of amyotrophic lateral sclerosis. Emphasis on kynurenine pathway inhibitors. Cent. Nerv. Syst. Agents Med. Chem. 2009, 9, 32–39. [Google Scholar]
- Mok, M.H.; Fricker, A.C.; Weil, A.; Kew, J.N. Electrophysiological characterisation of the actions of kynurenic acid at ligand-gated ion channels. Neuropharmacology 2009, 57, 242–249. [Google Scholar] [CrossRef]
- Potter, M.C.; Elmer, G.I.; Bergeron, R.; Albuquerque, E.X.; Guidetti, P.; Wu, H.Q.; Schwarcz, R. Reduction of endogenous kynurenic acid formation enhances extracellular glutamate, hippocampal plasticity, and cognitive behavior. Neuropsychopharmacology 2010, 35, 1734–1742. [Google Scholar]
- Cosi, C.; Mannaioni, G.; Cozzi, A.; Carla, V.; Sili, M.; Cavone, L.; Maratea, D.; Moroni, F. G-protein coupled receptor 35 (GPR35) activation and inflammatory pain: Studies on the antinociceptive effects of kynurenic acid and zaprinast. Neuropharmacology 2011, 60, 1227–1231. [Google Scholar] [CrossRef]
- Demeter, I.; Nagy, K.; Gellert, L.; Vecsei, L.; Fulop, F.; Toldi, J. A novel kynurenic acid analog (SZR104) inhibits pentylenetetrazole-induced epileptiform seizures. An electrophysiological study: Special issue related to kynurenine. J. Neural Transm. 2012, 119, 151–154. [Google Scholar] [CrossRef]
- Vecsei, L.; Horvath, Z.; Tuka, B. Old and New Neuroendocrine Molecules: Somatostatin, Cysteamine, Pantethine and Kynurenine. Ideggyógyászati Szemle 2014, 67, 107–112. [Google Scholar] [PubMed]
- Bagasrawala, I.; Zecevic, N.; Radonjic, N.V. N-Methyl D-Aspartate Receptor Antagonist Kynurenic Acid Affects Human Cortical Development. Front. Neurosci. 2016, 10, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahomi, R.B.; Nam, M.H.; Rankenberg, J.; Rakete, S.; Houck, J.A.; Johnson, G.C.; Stankowska, D.L.; Pantcheva, M.B.; MacLean, P.S.; Nagaraj, R.H. Kynurenic Acid Protects Against Ischemia/Reperfusion-Induced Retinal Ganglion Cell Death in Mice. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, M.; Terramani, T.; Lynch, G.; Baudry, M. A glycine site associated with N-methyl-D-aspartic acid receptors: Characterization and identification of a new class of antagonists. J. Neurochem. 1989, 52, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Mourao, A.A.; de Mello, A.B.S.; Dos Santos Moreira, M.C.; Rodrigues, K.L.; Lopes, P.R.; Xavier, C.H.; Gomes, R.M.; Freiria-Oliveira, A.H.; Blanch, G.T.; Colombari, E.; et al. Median preoptic nucleus excitatory neurotransmitters in the maintenance of hypertensive state. Brain Res. Bull. 2018, 142, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Lajko, E.; Tuka, B.; Fulop, F.; Krizbai, I.; Toldi, J.; Magyar, K.; Vecsei, L.; Kohidai, L. Kynurenic acid and its derivatives are able to modulate the adhesion and locomotion of brain endothelial cells. J. Neural Transm. 2018, 125, 899–912. [Google Scholar] [CrossRef]
- Lorinczi, B.; Csampai, A.; Fulop, F.; Szatmari, I. Synthesis of New C-3 Substituted Kynurenic Acid Derivatives. Molecules 2020, 25. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, K.; Feng, G.G.; Li, J.; Akahori, T.; Yasuda, Y.; Nakamura, E.; Hatakeyama, N.; Fujiwara, Y.; Kinoshita, H. Kynurenine causes vasodilation and hypotension induced by activation of KCNQ-encoded voltage-dependent K(+) channels. J. Pharmacol. Sci. 2015, 129, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Resta, F.; Masi, A.; Sili, M.; Laurino, A.; Moroni, F.; Mannaioni, G. Kynurenic acid and zaprinast induce analgesia by modulating HCN channels through GPR35 activation. Neuropharmacology 2016, 108, 136–143. [Google Scholar]
- Chen, H.; Smith, P.A. M-currents in frog sympathetic ganglion cells: Manipulation of membrane phosphorylation. Br. J. Pharmacol. 1992, 105, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Sankaranarayanan, S.; Simasko, S.M. Characterization of an M-like current modulated by thyrotropin-releasing hormone in normal rat lactotrophs. J. Neurosci. 1996, 16, 1668–1678. [Google Scholar] [CrossRef] [PubMed]
- Selyanko, A.A.; Hadley, J.K.; Wood, I.C.; Abogadie, F.C.; Delmas, P.; Buckley, N.J.; London, B.; Brown, D.A. Two types of K(+) channel subunit, Erg1 and KCNQ2/3, contribute to the M-like current in a mammalian neuronal cell. J. Neurosci. 1999, 19, 7742–7756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.A.; Passmore, G.M. Neural KCNQ (Kv7) channels. Br. J. Pharmacol. 2009, 156, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.T.; Tseng, Y.T.; Lo, Y.C.; Wu, S.N. Ability of naringenin, a bioflavonoid, to activate M-type potassium current in motor neuron-like cells and to increase BKCa-channel activity in HEK293T cells transfected with alpha-hSlo subunit. BMC Neurosci. 2014, 15, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.T.; Liu, P.Y.; Gao, Z.H.; Lee, S.W.; Lee, W.K.; Wu, S.N. Evidence for the Effectiveness of Remdesivir (GS-5734), a Nucleoside-Analog Antiviral Drug in the Inhibition of I K(M) or I K(DR) and in the Stimulation of I MEP. Front. Pharmacol. 2020, 11, 1091. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Hsiao, H.T.; Wang, J.C.; Liu, Y.C.; Wu, S.N. Parecoxib, a selective blocker of cyclooxygenase-2, directly inhibits neuronal delayed-rectifier K(+) current, M-type K(+) current and Na(+) current. Eur. J. Pharmacol. 2019, 844, 95–101. [Google Scholar] [CrossRef]
- Smith, P.A.; Chen, H.; Kurenny, D.E.; Selyanko, A.A.; Zidichouski, J.A. Regulation of the M current: Transduction mechanism and role in ganglionic transmission. Can. J. Physiol. Pharmacol. 1992, 70 (Suppl. S12–8). [Google Scholar] [CrossRef]
- Frohlich, H.; Boini, K.M.; Seebohm, G.; Strutz-Seebohm, N.; Ureche, O.N.; Foller, M.; Eichenmuller, M.; Shumilina, E.; Pathare, G.; Singh, A.K.; et al. Hypothyroidism of gene-targeted mice lacking Kcnq1. Pflugers Arch. 2011, 461, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Friedman, A.K.; Juarez, B.; Ku, S.M.; Zhang, H.; Calizo, R.C.; Walsh, J.J.; Chaudhury, D.; Zhang, S.; Hawkins, A.; Dietz, D.M.; et al. KCNQ channel openers reverse depressive symptoms via an active resilience mechanism. Nat. Commun. 2016, 7, 11671. [Google Scholar] [CrossRef] [Green Version]
- Vanhoof-Villalba, S.L.; Gautier, N.M.; Mishra, V.; Glasscock, E. Pharmacogenetics of KCNQ channel activation in 2 potassium channelopathy mouse models of epilepsy. Epilepsia 2018, 59, 358–368. [Google Scholar] [CrossRef] [Green Version]
- Haick, J.M.; Byron, K.L. Novel treatment strategies for smooth muscle disorders: Targeting Kv7 potassium channels. Pharmacol. Ther. 2016, 165, 14–25. [Google Scholar] [CrossRef] [PubMed]
- So, E.C.; Foo, N.P.; Ko, S.Y.; Wu, S.N. Bisoprolol, Known to Be a Selective beta(1)-Receptor Antagonist, Differentially but Directly Suppresses IK(M) and IK(erg) in Pituitary Cells and Hippocampal Neurons. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.S.; Lai, M.C.; Liu, P.Y.; Lo, Y.C.; Huang, C.W.; Wu, S.N. Characterization of the Inhibitory Effect of Gastrodigenin and Gastrodin on M-type K(+) Currents in Pituitary Cells and Hippocampal Neurons. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannikko, R.; Pandey, S.; Larsson, H.P.; Elinder, F. Hysteresis in the voltage dependence of HCN channels: Conversion between two modes affects pacemaker properties. J. Gen. Physiol. 2005, 125, 305–326. [Google Scholar] [CrossRef] [Green Version]
- Villalba-Galea, C.A. Hysteresis in voltage-gated channels. Channels 2017, 11, 140–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.T.; Lo, Y.C.; Wu, S.N. Characterization of Convergent Suppression by UCL-2077 (3-(Triphenylmethylaminomethyl)pyridine), Known to Inhibit Slow Afterhyperpolarization, of erg-Mediated Potassium Currents and Intermediate-Conductance Calcium-Activated Potassium Channels. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [Green Version]
- Villalba-Galea, C.A.; Chiem, A.T. Hysteretic Behavior in Voltage-Gated Channels. Front. Pharmacol. 2020, 11, 579596. [Google Scholar] [CrossRef]
- So, E.C.; Liu, P.Y.; Wu, S.N. Effectiveness in the inhibition of dapagliflozin and canagliflozin on M-type K(+) current and alpha-methylglucoside-induced current in pituitary tumor (GH3) and pheochromocytoma PC12 cells. Eur. J. Pharmacol. 2020, 879, 173141. [Google Scholar] [CrossRef]
- Wu, S.N.; Li, H.F.; Jan, C.R.; Shen, A.Y. Inhibition of Ca2+-activated K+ current by clotrimazole in rat anterior pituitary GH3 cells. Neuropharmacology 1999, 38, 979–989. [Google Scholar]
- Barros, F.; del Camino, D.; Pardo, L.A.; Palomero, T.; Giraldez, T.; de la Pena, P. Demonstration of an inwardly rectifying K+ current component modulated by thyrotropin-releasing hormone and caffeine in GH3 rat anterior pituitary cells. Pflugers Arch. 1997, 435, 119–129. [Google Scholar]
- Badzynska, B.; Zakrocka, I.; Turski, W.A.; Olszynski, K.H.; Sadowski, J.; Kompanowska-Jezierska, E. Kynurenic acid selectively reduces heart rate in spontaneously hypertensive rats. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.L.; Lu, T.J.; Wu, S.N. Effectiveness in Block by Dexmedetomidine of Hyperpolarization-Activated Cation Current, Independent of Its Agonistic Effect on alpha2-Adrenergic Receptors. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Lu, T.L.; Lu, T.J.; Wu, S.N. Inhibitory Effective Perturbations of Cilobradine (DK-AH269), A Blocker of HCN Channels, on the Amplitude and Gating of Both Hyperpolarization-Activated Cation and Delayed-Rectifier Potassium Currents. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.J.; Lin, M.W.; Lin, A.A.; Wu, S.N. Riluzole-induced block of voltage-gated Na+ current and activation of BKCa channels in cultured differentiated human skeletal muscle cells. Life Sci. 2008, 82, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Wu, B.; Zhang, W.; Ji, Y.H.; Wang, Q.; Tan, Z.Y. Inhibitory Effects of Columbianadin on Nociceptive Behaviors in a Neuropathic Pain Model, and on Voltage-Gated Calcium Currents in Dorsal Root Ganglion Neurons in Mice. Front. Pharmacol. 2019, 10, 1522. [Google Scholar] [PubMed]
- Wu, S.N.; Wu, Y.H.; Chen, B.S.; Lo, Y.C.; Liu, Y.C. Underlying mechanism of actions of tefluthrin, a pyrethroid insecticide, on voltage-gated ion currents and on action currents in pituitary tumor (GH3) cells and GnRH-secreting (GT1-7) neurons. Toxicology 2009, 258, 70–77. [Google Scholar] [CrossRef]
- Chen, T.S.; Lai, M.C.; Hung, T.Y.; Lin, K.M.; Huang, C.W.; Wu, S.N. Pioglitazone, a PPAR-gamma Activator, Stimulates BKCa but Suppresses IK M in Hippocampal Neurons. Front. Pharmacol. 2018, 9, 977. [Google Scholar] [CrossRef]
- Lai, M.C.; Tzeng, R.C.; Huang, C.W.; Wu, S.N. The Novel Direct Modulatory Effects of Perampanel, an Antagonist of AMPA Receptors, on Voltage-Gated Sodium and M-type Potassium Currents. Biomolecules 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Nagy, K.; Plangar, I.; Tuka, B.; Gellert, L.; Varga, D.; Demeter, I.; Farkas, T.; Kis, Z.; Marosi, M.; Zadori, D.; et al. Synthesis and biological effects of some kynurenic acid analogs. Bioorg. Med. Chem. 2011, 19, 7590–7596. [Google Scholar]
- Hsiao, H.T.; Liu, Y.C.; Liu, P.Y.; Wu, S.N. Concerted suppression of Ih and activation of IK(M) by ivabradine, an HCN-channel inhibitor, in pituitary cells and hippocampal neurons. Brain Res. Bull. 2019, 149, 11–20. [Google Scholar] [CrossRef]
- Wu, S.N.; Hsu, M.C.; Liao, Y.K.; Wu, F.T.; Jong, Y.J.; Lo, Y.C. Evidence for inhibitory effects of flupirtine, a centrally acting analgesic, on delayed rectifier k(+) currents in motor neuron-like cells. Evid. Based Complement. Alternat. Med. 2012, 2012, 148403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villalobos, C.; Garcia-Sancho, J. Glutamate increases cytosolic calcium in GH3 pituitary cells acting via a high-affinity glutamate transporter. FASEB J. 1995, 9, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, F.P.; Fox, B.K.; Chan, W.Y.; Davis, L.K.; Andres, M.A.; Hirano, T.; Grau, E.G.; Cooke, I.M. Ionotropic glutamate receptor activation increases intracellular calcium in prolactin-releasing cells of the adenohypophysis. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E1188–E1196. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Simonavicius, N.; Wu, X.; Swaminath, G.; Reagan, J.; Tian, H.; Ling, L. Kynurenic acid as a ligand for orphan G protein-coupled receptor GPR35. J. Biol. Chem. 2006, 281, 22021–22028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkondon, M.; Pereira, E.F.; Todd, S.W.; Randall, W.R.; Lane, M.V.; Albuquerque, E.X. Functional G-protein-coupled receptor 35 is expressed by neurons in the CA1 field of the hippocampus. Biochem. Pharmacol. 2015, 93, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Haitin, Y.; Attali, B. The C-terminus of Kv7 channels: A multifunctional module. J. Physiol. 2008, 586, 1803–1810. [Google Scholar] [PubMed]
- Labro, A.J.; Priest, M.F.; Lacroix, J.J.; Snyders, D.J.; Bezanilla, F. Kv3.1 uses a timely resurgent K(+) current to secure action potential repolarization. Nat. Commun. 2015, 6, 10173. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.D.; Whetsell, W.O., Jr.; Crowley, W.R. Quinolinic acid stimulates luteinizing hormone secretion in female rats: Evidence for involvement of N-methyl-D-aspartate-preferring receptors. Exp. Brain Res. 1985, 59, 57–61. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.; McKenzie, G.; Witting, P.K.; Stasch, J.P.; Hahn, M.; Changsirivathanathamrong, D.; Wu, B.J.; Ball, H.J.; Thomas, S.R.; et al. Kynurenine is an endothelium-derived relaxing factor produced during inflammation. Nat. Med. 2010, 16, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Harsing, L.G., Jr.; Solyom, S.; Salamon, C. The role of glycineB binding site and glycine transporter (GlyT1) in the regulation of [3H]GABA and [3H]glycine release in the rat brain. Neurochem. Res. 2001, 26, 915–923. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Dunaway, E.P.; Bhattacharya, S.; Bloemer, J.; Buabeid, M.; Escobar, M.; Suppiramaniam, V.; Dhanasekaran, M. Impaired ILK Function Is Associated with Deficits in Hippocampal Based Memory and Synaptic Plasticity in a FASD Rat Model. PLoS ONE 2015, 10, e0135700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Muthuchamy, M.; Reddy, D.S. Atomic Force Microscopy Protocol for Measurement of Membrane Plasticity and Extracellular Interactions in Single Neurons in Epilepsy. Front. Aging Neurosci. 2016, 8, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carver, C.M.; Chuang, S.H.; Reddy, D.S. Zinc Selectively Blocks Neurosteroid-Sensitive Extrasynaptic deltaGABAA Receptors in the Hippocampus. J. Neurosci. 2016, 36, 8070–8077. [Google Scholar] [CrossRef] [PubMed]
- Carver, C.M.; Hastings, S.D.; Cook, M.E.; Shapiro, M.S. Functional responses of the hippocampus to hyperexcitability depend on directed, neuron-specific KCNQ2 K(+) channel plasticity. Hippocampus 2020, 30, 435–455. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.H.; Reddy, D.S. 3beta-Methyl-Neurosteroid Analogs Are Preferential Positive Allosteric Modulators and Direct Activators of Extrasynaptic delta-Subunit gamma-Aminobutyric Acid Type A Receptors in the Hippocampus Dentate Gyrus Subfield. J. Pharmacol. Exp. Ther. 2018, 365, 583–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalby, N.O.; Falk-Petersen, C.B.; Leurs, U.; Scholze, P.; Krall, J.; Frolund, B.; Wellendorph, P. Silencing of spontaneous activity at alpha4beta1/3delta GABAA receptors in hippocampal granule cells reveals different ligand pharmacology. Br. J. Pharmacol. 2020, 177, 3975–3990. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.R.; Zhou, H.F.; Liu, B.; Jiang, X.F.; Xu, J.Y.; Jiang, M. The Effect of Bushen Zhuyun Prescription on Gonadotropic Hormone Secretion and Key Transcription Factor Expression in Rat Pituitary Cells. Nanosci. Nanotechnol. Lett. 2017, 9, 1596–1603. [Google Scholar]
- Wu, S.N.; Chern, J.H.; Shen, S.; Chen, H.H.; Hsu, Y.T.; Lee, C.C.; Chan, M.H.; Lai, M.C.; Shie, F.S. Stimulatory actions of a novel thiourea derivative on large-conductance, calcium-activated potassium channels. J. Cell. Physiol. 2017, 232, 3409–3421. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo, Y.-C.; Lin, C.-L.; Fang, W.-Y.; Lőrinczi, B.; Szatmári, I.; Chang, W.-H.; Fülöp, F.; Wu, S.-N. Effective Activation by Kynurenic Acid and Its Aminoalkylated Derivatives on M-Type K+ Current. Int. J. Mol. Sci. 2021, 22, 1300. https://doi.org/10.3390/ijms22031300
Lo Y-C, Lin C-L, Fang W-Y, Lőrinczi B, Szatmári I, Chang W-H, Fülöp F, Wu S-N. Effective Activation by Kynurenic Acid and Its Aminoalkylated Derivatives on M-Type K+ Current. International Journal of Molecular Sciences. 2021; 22(3):1300. https://doi.org/10.3390/ijms22031300
Chicago/Turabian StyleLo, Yi-Ching, Chih-Lung Lin, Wei-Yu Fang, Bálint Lőrinczi, István Szatmári, Wan-Hsuan Chang, Ferenc Fülöp, and Sheng-Nan Wu. 2021. "Effective Activation by Kynurenic Acid and Its Aminoalkylated Derivatives on M-Type K+ Current" International Journal of Molecular Sciences 22, no. 3: 1300. https://doi.org/10.3390/ijms22031300