
High-Temperature Conditions Promote Soybean Flowering through the Transcriptional Reprograming of Flowering Genes in the Photoperiod Pathway

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Climatron Condition Affects Soybean Growth and Development Such as Flowering
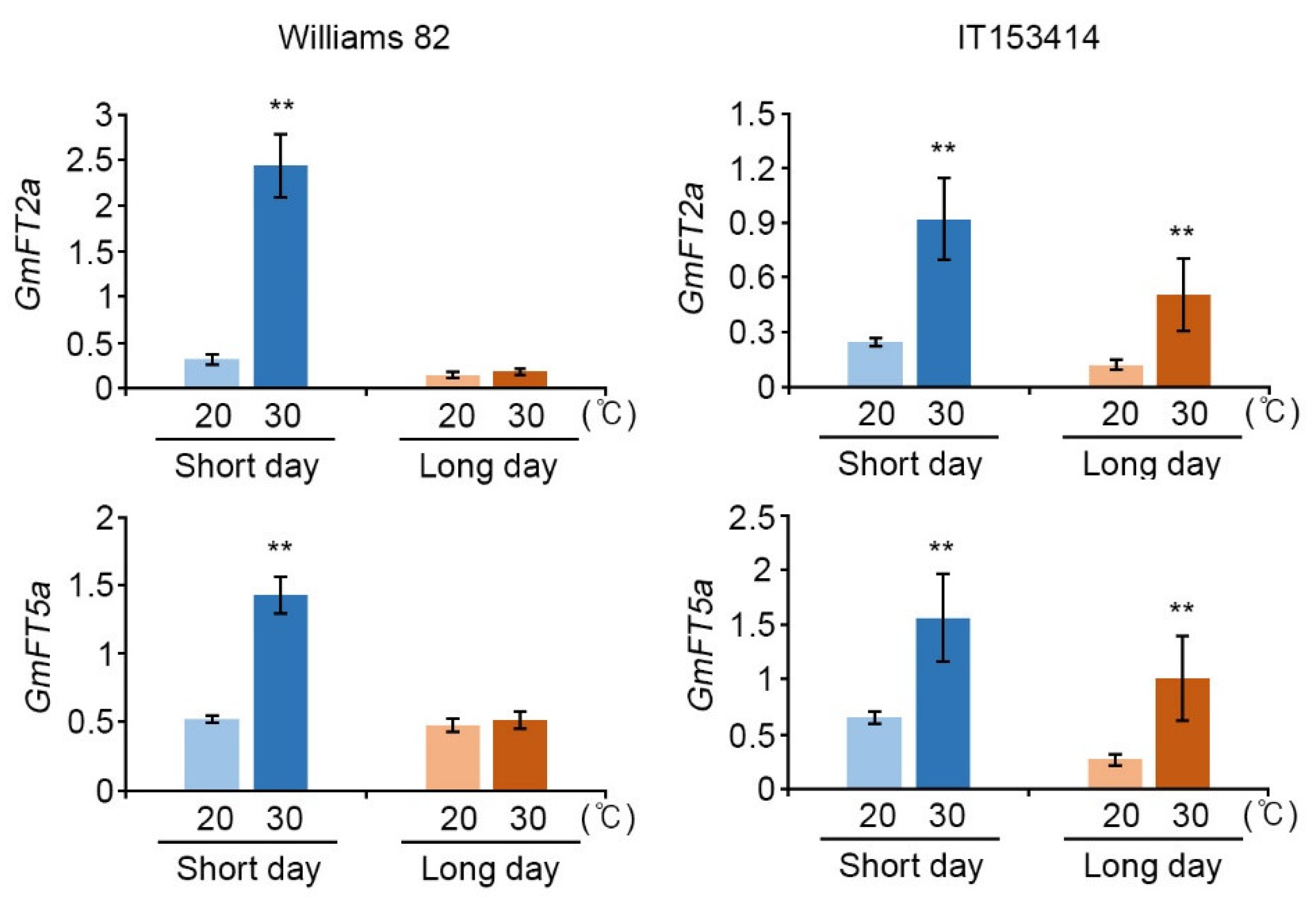
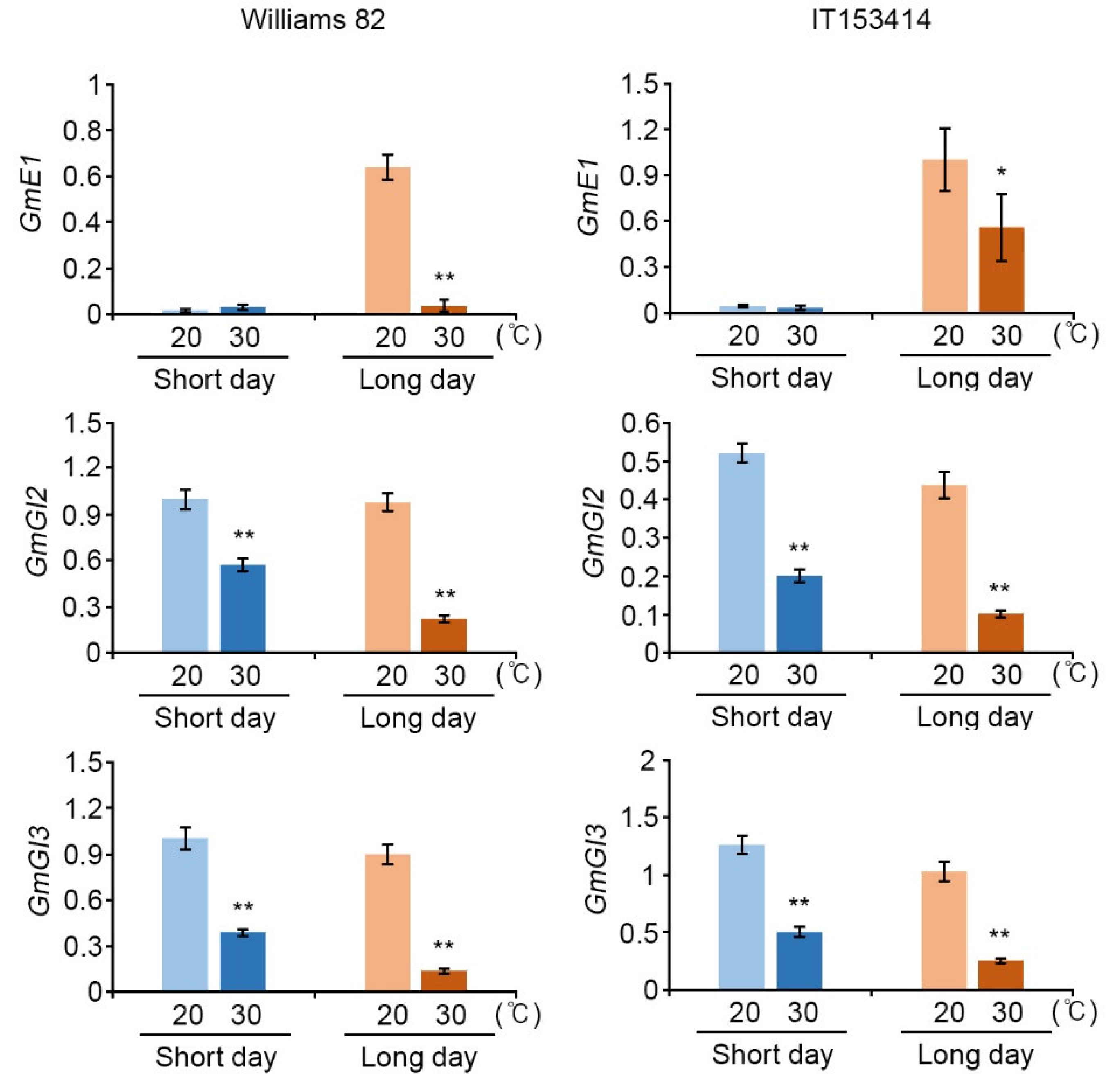
2.2. High-Temperature Conditions Regulated Expression of Soybean Flowering Genes in Photoperiod Pathway
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Semi-Quantitative Reverse Transcription Polymerase Chain Reaction (RT-PCR) and Quantitative Real Time PCR (qRT-PCR)
4.3. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lean, J.L.; Rind, D.H. How will Earth’s surface temperature change in future decades? Geophys. Res. Lett. 2009, 36, 1–5. [Google Scholar] [CrossRef]
- Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Khoury, C.K.; Müller, J.V.; Toll, J. Adapting agriculture to climate change: A global initiative to collect, conserve, and use crop wild relatives. Agroecol. Sustain. Food Syst. 2014, 38, 369–377. [Google Scholar] [CrossRef]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate extremes: Observations, modeling, and impacts. Science 2000, 289, 2068–2075. [Google Scholar] [CrossRef] [PubMed]
- Rogelj, J.; Shindell, D.; Jiang, K.; Fifita, S.; Forster, P.; Ginzburg, V.; Handa, C.; Kheshgi, H.; Kobayashi, S.; Kriegler, E.; et al. Mitigation pathways compatible with 1.5 °C in the context of sustainable development. In Global Warming of 1.5°C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways; International Institute for Applied Systems Analysis (IIASA): Laxenburg, Austria, 2018; p. 82. [Google Scholar]
- Leung, J.Y.S.; Russell, B.D.; Connell, S.D. Adaptive responses of marine gastropods to heatwaves. One Earth 2019, 1, 374–381. [Google Scholar] [CrossRef]
- White, J.W.; Hoogenboom, G.; Kimball, B.A.; Wall, G.W. Methodologies for simulating impacts of climate change on crop production. F. Crop. Res. 2011, 124, 357–368. [Google Scholar] [CrossRef]
- Lippmann, R.; Babben, S.; Menger, A.; Delker, C.; Quint, M. Development of wild and cultivated plants under global warming conditions. Curr. Biol. 2019, 29, R1326–R1338. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K. Plant environmental stress responses for survival and biomass enhancement. In Climate Change and Plant Abiotic Stress Tolerance; Wiley: Hoboken, NJ, USA, 2013; pp. 79–108. [Google Scholar] [CrossRef]
- Mishra, A.K.; Agrawal, S.B.; Agrawal, M. Rising Atmospheric Carbon Dioxide and Plant Responses: Current and Future Consequences; Elsevier Inc.: Amsterdam, The Netherlands, 2019; ISBN 9780128164839. [Google Scholar]
- Terzić, D.; Popović, V.; Tatić, M.; Vasileva, V.; Đekić, V.; Ugrenović, V.; Popović, S.; Avdić, P. Soybean area, yield and production in world. In Proceedings of the XXII Eco-Conference Ecological Movement, Novi Sad, Serbia, 26–28 September 2018; pp. 135–145. [Google Scholar]
- Roy, R.; Finck, A.; Blair, G.; Tandon, H. Plant Nutrition for food security. A guide for integrated nutrient management. In Fertilizer and Plant Nutrition Bulletin 16; FAO, Ed.; Food and Agricultural Organization of United Nationals: Rome, Italy, 2006; pp. 235–348. ISBN 92-5-105490-8. [Google Scholar]
- Hassan, S.M. Soybean, nutrition and health. In Soybean-Bio-Active Compounds; InTech: London, UK, 2013; pp. 453–473. [Google Scholar]
- Li, Y.H.; Zhou, G.; Ma, J.; Jiang, W.; Jin, L.G.; Zhang, Z.; Guo, Y.; Zhang, J.; Sui, Y.; Zheng, L.; et al. De novo assembly of soybean wild relatives for pan-genome analysis of diversity and agronomic traits. Nat. Biotechnol. 2014, 32, 1045–1052. [Google Scholar] [CrossRef]
- Watanabe, S.; Harada, K.; Abe, J. Genetic and molecular bases of photoperiod responses of flowering in soybean. Breed. Sci. 2011, 61, 531–543. [Google Scholar] [CrossRef]
- Amasino, R. Seasonal and developmental timing of flowering. Plant J. 2010, 61, 1001–1013. [Google Scholar] [CrossRef]
- Song, Y.H.; Shim, J.S.; Kinmonth-Schultz, H.A.; Imaizumi, T. Photoperiodic flowering: Time measurement mechanisms in leaves. Annu. Rev. Plant Biol. 2015, 66, 441–464. [Google Scholar] [CrossRef]
- Turck, F.; Fornara, F.; Coupland, G. Regulation and identity of florigen: Flowering locus T moves center stage. Annu. Rev. Plant Biol. 2008, 59, 573–594. [Google Scholar] [CrossRef] [PubMed]
- Suárez-López, P.; Wheatley, K.; Robson, F.; Onouchi, H.; Valverde, F.; Coupland, G. Constans mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 2001, 410, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- Golembeski, G.S.; Kinmonth-Schultz, H.A.; Song, Y.H.; Imaizumi, T. Photoperiodic Flowering Regulation in Arabidopsis Thaliana; Elsevier: Amsterdam, The Netherlands, 2014; Volume 72, ISBN 9780124171626. [Google Scholar]
- Cao, D.; Takeshima, R.; Zhao, C.; Liu, B.; Jun, A.; Kong, F. Molecular mechanisms of fowering under long days and stem growth habit in soybean. J. Exp. Bot. 2017, 68, 1873–1884. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Shin, J.H.; Kang, Y.J.; Shim, S.R.; Lee, S.H. Divergence of flowering genes in soybean. J. Biosci. 2012, 37, 857–870. [Google Scholar] [CrossRef] [PubMed]
- Cober, E.R.; Morrison, M.J. Regulation of seed yield and agronomic characters by photoperiod sensitivity and growth habit genes in soybean. Theor. Appl. Genet. 2010, 120, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Zhai, H.; Liu, B.; Kong, F.; Yuan, X.; Wu, H.; Cober, E.R.; Harada, K. Molecular identification of genes controlling flowering time, maturity, and photoperiod response in soybean. Plant. Syst. Evol. 2012, 298, 1217–1227. [Google Scholar] [CrossRef]
- Xia, Z.; Watanabe, S.; Yamada, T.; Tsubokura, Y.; Nakashima, H.; Zhai, H.; Anai, T.; Sato, S.; Yamazaki, T.; Lü, S.; et al. Positional cloning and characterization reveal the molecular basis for soybean maturity locus E1 that regulates photoperiodic flowering. Proc. Natl. Acad. Sci. USA 2012, 109. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, T.; Wright, L.; Fujiwara, S.; Cremer, F.; Lee, K.; Onouchi, H.; Mouradov, A.; Fowler, S.; Kamada, H.; Putterill, J.; et al. Distinct roles of GIGANTEA in promoting flowering and regulating circadian rhythms in Arabidopsis. Plant. Cell 2005, 17, 2255–2270. [Google Scholar] [CrossRef]
- Zhao, C.; Takeshima, R.; Zhu, J.; Xu, M.; Sato, M.; Watanabe, S.; Kanazawa, A.; Liu, B.; Kong, F.; Yamada, T.; et al. A recessive allele for delayed flowering at the soybean maturity locus E9 is a leaky allele of FT2a, a flowering locus T ortholog. BMC Plant Biol. 2016, 16. [Google Scholar] [CrossRef]
- Kong, F.; Liu, B.; Xia, Z.; Sato, S.; Kim, B.M.; Watanabe, S.; Yamada, T.; Tabata, S.; Kanazawa, A.; Harada, K.; et al. Two coordinately regulated homologs of flowering locus T are involved in the control of photoperiodic flowering in Soybean. Plant Physiol. 2010, 154, 1220–1231. [Google Scholar] [CrossRef]
- Song, Y.H.; Ito, S.; Imaizumi, T. Flowering time regulation: Photoperiod- and temperature-sensing in leaves. Trends Plant Sci. 2013, 18, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Price, B.W.; Haider, W.; Seufferheld, G.; Nelson, R.; Hanzawa, Y. Functional and evolutionary characterization of the CONSTANS gene family in short-day photoperiodic flowering in soybean. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Li, Y.; Lu, S.; Wang, J.; Nan, H.; Li, X.; Shi, D.; Fang, C.; Zhai, H.; Yuan, X.; et al. GmCOL1a and GmCOL1b function as flowering repressors in soybean under long-day conditions. Plant Cell Physiol. 2015, 56, 2409–2422. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef]
- Eberhard, S.; Finazzi, G.; Wollman, F.A. The dynamics of photosynthesis. Annu. Rev. Genet. 2008, 42, 463–515. [Google Scholar] [CrossRef]
- Habekotté, B. Quantitative analysis of pod formation, seed set and seed filling in winter oilseed rape (Brassica napus L.) under field conditions. F. Crop. Res. 1993, 35, 21–33. [Google Scholar] [CrossRef]
- Bunce, J.A. Elevated carbon dioxide effects on reproductive phenology and seed yield among soybean cultivars. Crop. Sci. 2015, 55, 339–343. [Google Scholar] [CrossRef]
- Sreeharsha, R.V.; Sekhar, K.M.; Reddy, A.R. Delayed flowering is associated with lack of photosynthetic acclimation in Pigeon pea (Cajanus cajan L.) grown under elevated CO2. Plant Sci. 2015, 231, 82–93. [Google Scholar] [CrossRef]
- Shimono, H.; Okada, M.; Yamakawa, Y.; Nakamura, H.; Kobayashi, K.; Hasegawa, T. Genotypic variation in rice yield enhancement by elevated CO2 relates to growth before heading, and not to maturity group. J. Exp. Bot. 2009, 60, 523–532. [Google Scholar] [CrossRef]
- Cleland, E.E.; Chiariello, N.R.; Loarie, S.R.; Mooney, H.A.; Field, C.B. Diverse responses of phenology to global changes in a grassland ecosystem. Proc. Natl. Acad. Sci. USA 2006, 103, 13740–13744. [Google Scholar] [CrossRef]
- Valverde, F. CONSTANS and the evolutionary origin of photoperiodic timing of flowering. J. Exp. Bot. 2011, 62, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Hu, R.; Zhang, X.; Wang, X.; Zhang, W.; Zhang, Q.; Ma, J.; Fu, Y.F. Conserved CO-FT regulons contribute to the photoperiod flowering control in soybean. BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Ort, D.R. More than taking the heat: Crops and global change. Curr. Opin. Plant Biol. 2010, 13, 240–247. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Siebers, M.H.; Yendrek, C.R.; Drag, D.; Locke, A.M.; Rios Acosta, L.; Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Ort, D.R. Heat waves imposed during early pod development in soybean (Glycine max) cause significant yield loss despite a rapid recovery from oxidative stress. Glob. Chang. Biol. 2015, 21, 3114–3125. [Google Scholar] [CrossRef] [PubMed]
- Petkova, V.; Nikolova, V.; Kalapchieva, S.H.; Stoeva, V.; Topalova, E.; Angelova, S. Physiological response and pollen viability of Pisum sativum genotypes under high temperature influence. Acta Hortic. 2009, 830, 665–671. [Google Scholar] [CrossRef]
- Liu, Y.; Li, J.; Zhu, Y.; Jones, A.; Rose, R.J.; Song, Y. Heat stress in legume seed setting: Effects, causes, and future prospects. Front. Plant. Sci. 2019, 10. [Google Scholar] [CrossRef]
- Slattery, R.A.; Ort, D.R. Carbon assimilation in crops at high temperatures. Plant. Cell Environ. 2019, 42, 2750–2758. [Google Scholar] [CrossRef]
- Ruiz-Vera, U.M.; Siebers, M.; Gray, S.B.; Drag, D.W.; Rosenthal, D.M.; Kimball, B.A.; Ort, D.R.; Bernacchi, C.J. Global warming can negate the expected CO2 stimulation in photosynthesis and productivity for soybean grown in the midwestern United States. Plant. Physiol. 2013, 162, 410–423. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.M.; Nayyar, H. Identification of high-temperature tolerant lentil (Lens culinaris Medik.) genotypes through leaf and pollen traits. Front. Plant Sci. 2017, 8, 1–27. [Google Scholar] [CrossRef]
- Kaushal, N.; Awasthi, R.; Gupta, K.; Gaur, P.; Siddique, K.H.M.; Nayyar, H. Heat-stress-induced reproductive failures in chickpea (Cicer arietinum) are associated with impaired sucrose metabolism in leaves and anthers. Funct. Plant. Biol. 2013, 40, 1334–1349. [Google Scholar] [CrossRef] [PubMed]
- Nagatoshi, Y.; Fujita, Y. Accelerating soybean breeding in a CO2-supplemented growth chamber. Plant Cell Physiol. 2019, 60, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, A.; Schmid, M. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef] [PubMed]
- Ausín, I.; Alonso-Blanco, C.; Martínez-Zapater, J.M. Environmental regulation of flowering. Int. J. Dev. Biol. 2005, 49, 689–705. [Google Scholar] [CrossRef] [PubMed]
- Fernández, V.; Takahashi, Y.; Le Gourrierec, J.; Coupland, G. Photoperiodic and thermosensory pathways interact through CONSTANS to promote flowering at high temperature under short days. Plant J. 2016. [Google Scholar] [CrossRef]
- Craufurd, P.Q.; Wheeler, T.R. Climate change and the flowering time of annual crops. J. Exp. Bot. 2009, 60, 2529–2539. [Google Scholar] [CrossRef]
- Amano, T.; Smithers, R.J.; Sparks, T.H.; Sutherland, W.J. A 250-year index of first flowering dates and its response to temperature changes. Proc. R. Soc. B Biol. Sci. 2010, 277, 2451–2457. [Google Scholar] [CrossRef]
- Miller-Rushing, A.J.; Primack, R.B. Global warming and flowering times in Thoreau’s Concord: A community perspective. Ecology 2008, 89, 332–341. [Google Scholar] [CrossRef]
- Devasirvatham, V.; Gaur, P.M.; Mallikarjuna, N.; Tokachichu, R.N.; Trethowan, R.M.; Tan, D.K.Y. Effect of high temperature on the reproductive development of chickpea genotypes under controlled environments. Funct. Plant Biol. 2012, 39, 1009–1018. [Google Scholar] [CrossRef]
- Wu, F.; Hanzawa, Y. Photoperiodic control of flowering in plants. Trends Plant Sci. 2014, 121–138. [Google Scholar] [CrossRef]
- Weller, J.L.; Ortega, R. Genetic control of flowering time in legumes. Front. Plant Sci. 2015, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, M.; Dwiyanti, M.S.; Watanabe, S.; Yamada, T.; Hase, Y.; Kanazawa, A.; Sayama, T.; Ishimoto, M.; Liu, B.; et al. A soybean deletion mutant that moderates the repression of flowering by cool temperatures. Front. Plant Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Kimura, S. Plant temperature sensors. Sensors (Switzerland) 2018, 18, 4365. [Google Scholar] [CrossRef] [PubMed]
- Márquez, A.J.; Stougaard, J.; Udvardi, M.; Parniske, M.; Spaink, H.; Saalbach, G.; Webb, J.; Chiurazzi, M.; Márquez, A.J. (Eds.) RNA isolation using phase extraction and LiCl precipitation. In Lotus Japonicus Handbook; Springer: Berlin/Heidelberg, Germany, 2005; pp. 123–124. ISBN 978-1-4020-3735-1. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
No, D.H.; Baek, D.; Lee, S.H.; Cheong, M.S.; Chun, H.J.; Park, M.S.; Cho, H.M.; Jin, B.J.; Lim, L.H.; Lee, Y.B.; et al. High-Temperature Conditions Promote Soybean Flowering through the Transcriptional Reprograming of Flowering Genes in the Photoperiod Pathway. Int. J. Mol. Sci. 2021, 22, 1314. https://doi.org/10.3390/ijms22031314
No DH, Baek D, Lee SH, Cheong MS, Chun HJ, Park MS, Cho HM, Jin BJ, Lim LH, Lee YB, et al. High-Temperature Conditions Promote Soybean Flowering through the Transcriptional Reprograming of Flowering Genes in the Photoperiod Pathway. International Journal of Molecular Sciences. 2021; 22(3):1314. https://doi.org/10.3390/ijms22031314
Chicago/Turabian StyleNo, Dong Hyeon, Dongwon Baek, Su Hyeon Lee, Mi Sun Cheong, Hyun Jin Chun, Mi Suk Park, Hyun Min Cho, Byung Jun Jin, Lack Hyeon Lim, Yong Bok Lee, and et al. 2021. "High-Temperature Conditions Promote Soybean Flowering through the Transcriptional Reprograming of Flowering Genes in the Photoperiod Pathway" International Journal of Molecular Sciences 22, no. 3: 1314. https://doi.org/10.3390/ijms22031314
APA StyleNo, D. H., Baek, D., Lee, S. H., Cheong, M. S., Chun, H. J., Park, M. S., Cho, H. M., Jin, B. J., Lim, L. H., Lee, Y. B., Shim, S. I., Chung, J.-I., & Kim, M. C. (2021). High-Temperature Conditions Promote Soybean Flowering through the Transcriptional Reprograming of Flowering Genes in the Photoperiod Pathway. International Journal of Molecular Sciences, 22(3), 1314. https://doi.org/10.3390/ijms22031314