
Expression Signature of lncRNAs and mRNAs in Sevoflurane-Induced Mouse Brain Injury: Implication of Involvement of Wide Molecular Networks and Pathways



Abstract
1. Introduction
2. Results
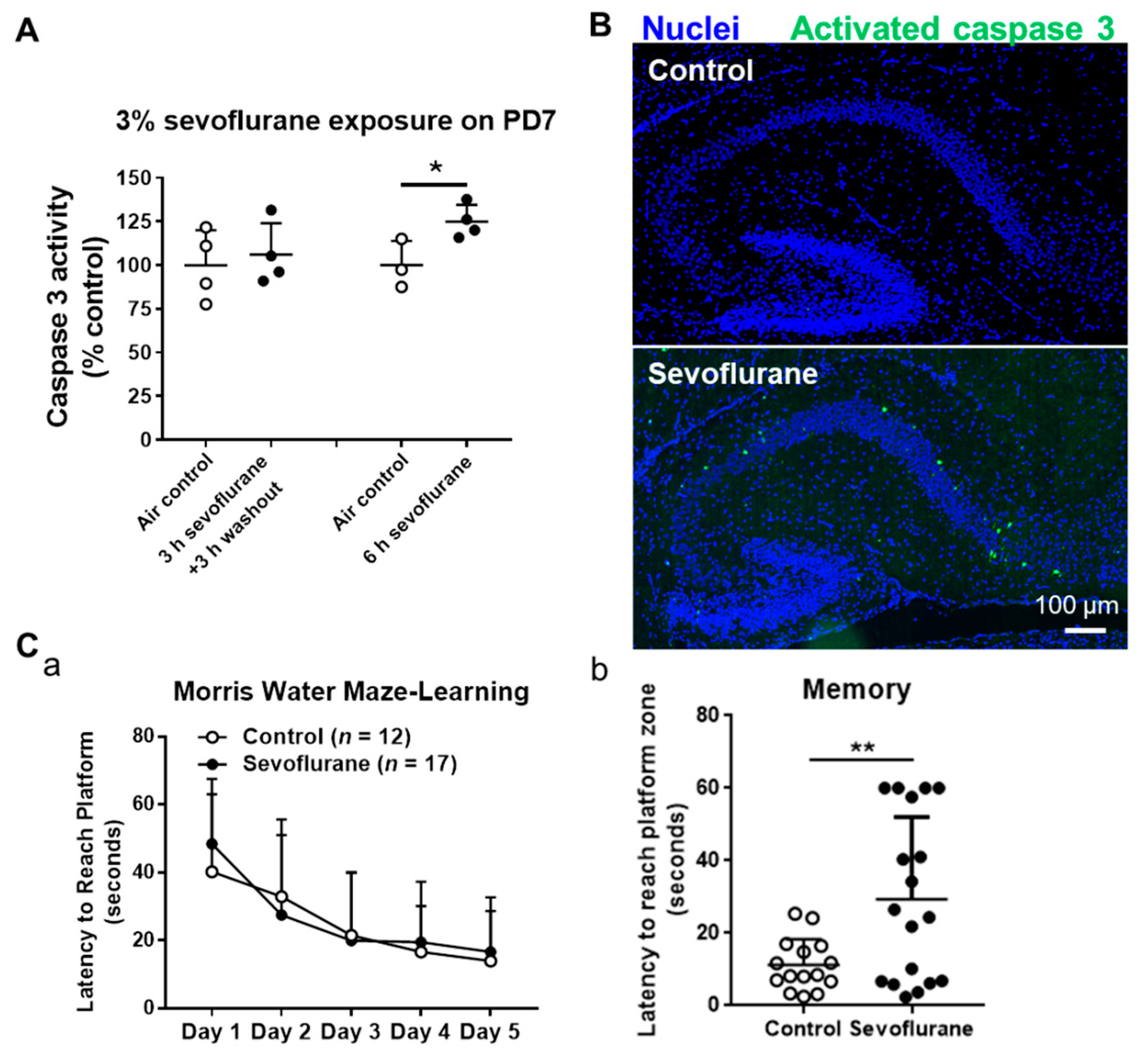
2.1. Sevoflurane Induces Neuroapoptosis in the Neonatal Mouse Hippocampi in a Time-Dependent Manner
2.2. Exposure of Sevoflurane to PD7 Mice Leads to Impaired Memory Capacity in PD60 Adult Mice
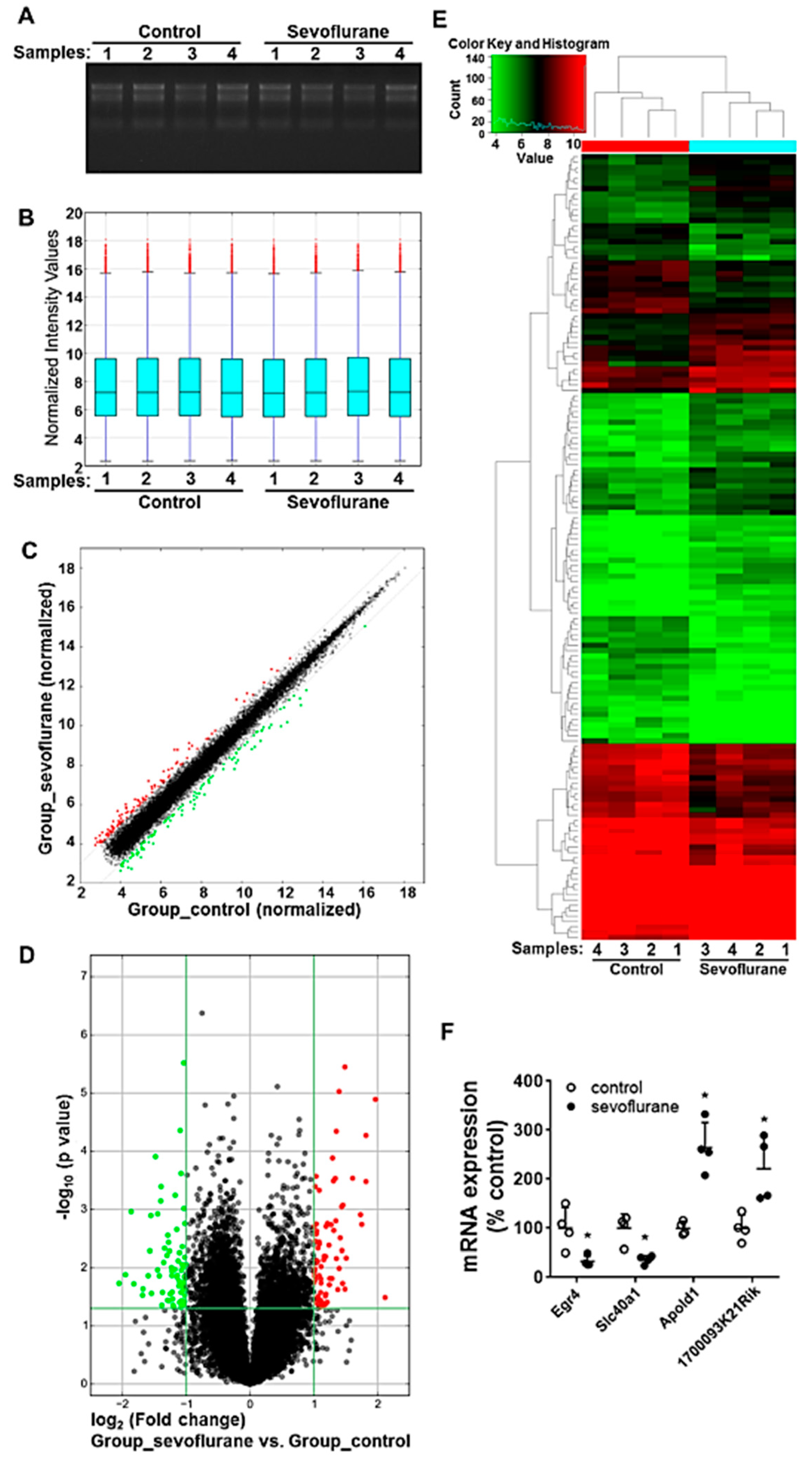
2.3. Sevoflurane Exposure on PD7 Induces the Alteration of mRNA Profiles in Mouse Hippocampi
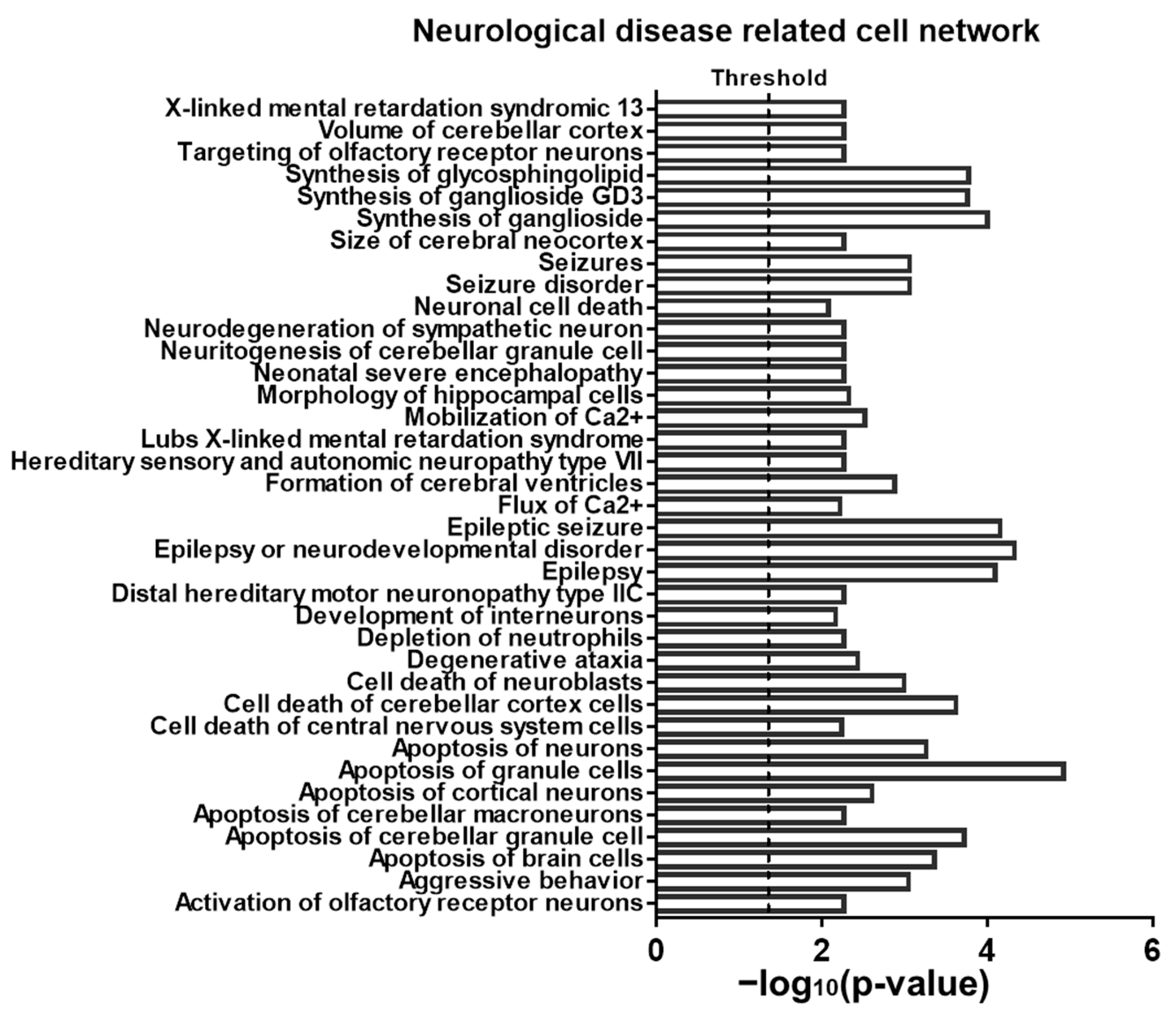
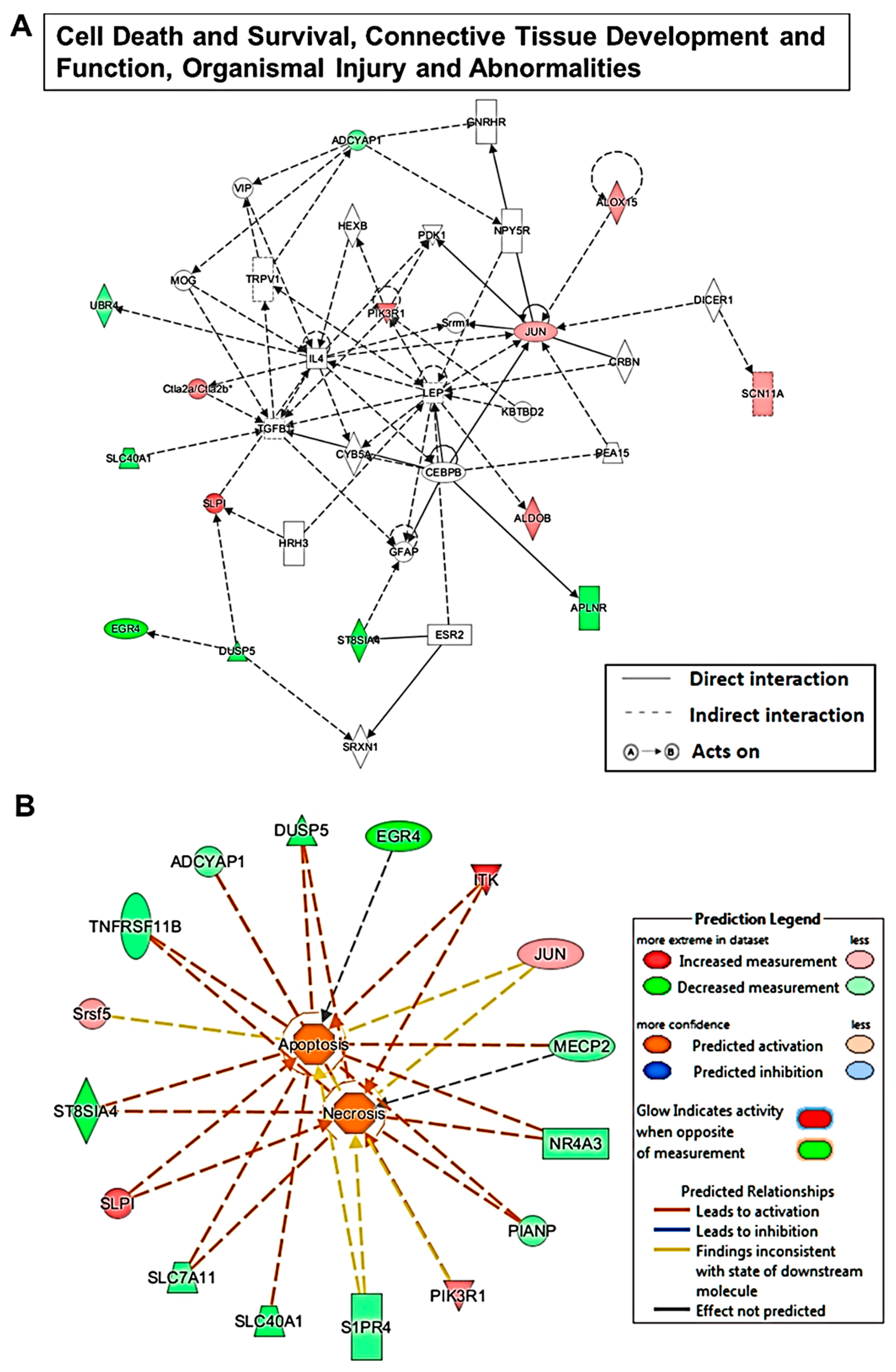
2.4. Sevoflurane Exposure-Induced Dysregulated mRNA Are Potentially Involved in Neurotoxicity-Related Signalling
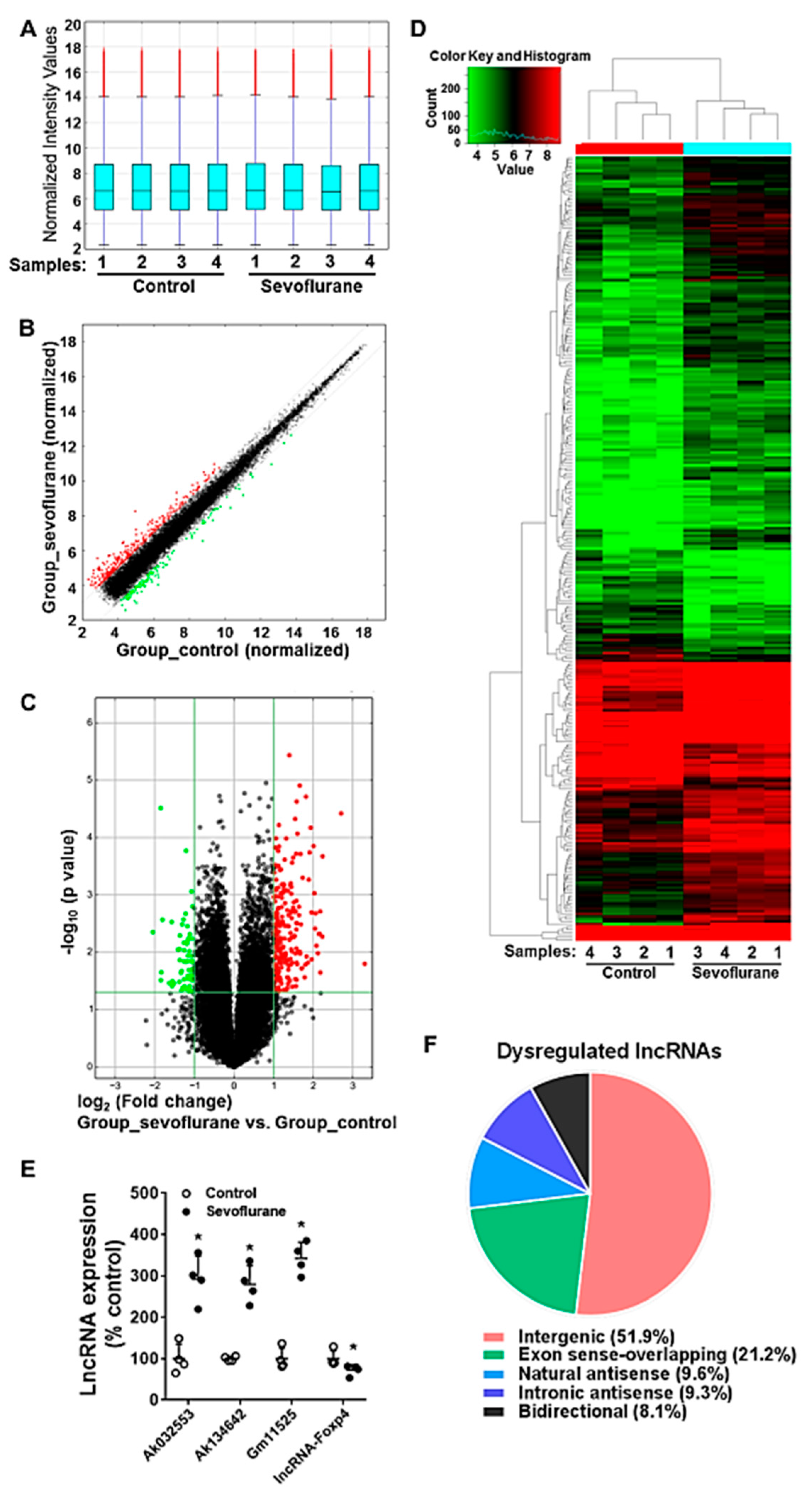
2.5. Sevoflurane Induces the Alteration of lncRNA Expression in PD7 Mouse Hippocampi
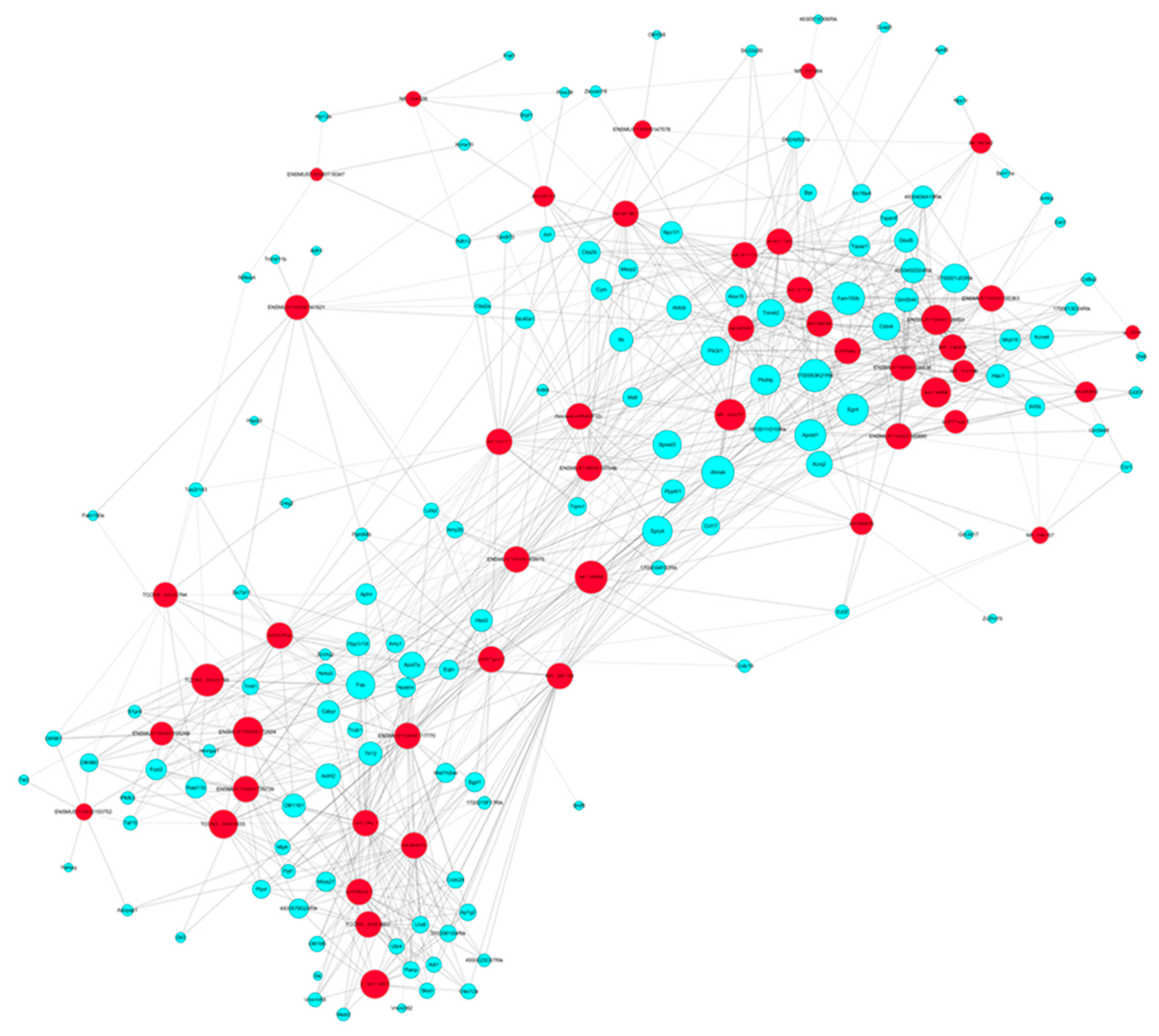
2.6. Sevoflurane Exposure Results in the Dysregulated lncRNAs and Their Corresponding co-Expressed mRNAs in Mouse Hippocampi
2.7. The Predicted Signaling Networks Formed between Sevoflurane Exposure-Induced Altered lncRNAs and mRNAs
3. Discussion
4. Materials and Methods
4.1. Animal Studies
4.2. Anesthetic Exposure
4.3. Caspase 3 Acitivity Assay
4.4. Immunofluorescence Staining
4.5. Cognitive Function Assay
4.6. RNA Extraction
4.7. Microarray Assay
4.8. RT-qPCR
4.9. IPA of Dysregulated Mrnas and Related Pathways
4.10. Networks Analysis of Dysregulated lncRNAs-Related Pathways and lncRNA-mRNA Interactions
4.11. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AIDN | anesthetics-induced developmental neurotoxicity |
| CA | cornus ammonis |
| CNC | coding and non-coding gene co-expression |
| Ct | cycle threshold |
| CNS | central nervous system |
| ESCs | embryonic stem cells |
| FDR | false-discovery rate |
| GABA | γ-aminobutyric acid |
| IPA | ingenuity pathway analysis |
| lncRNAs | long non-coding RNAs |
| mRNAs | messenger RNAs |
| LTD | long-term depression |
| NSC | neural stem cells |
| PCC | Pearson correlation coefficient |
| PD | postnatal day |
| RT-qPCR | reverse transcription-quantitative polymerase chain reaction |
| SD | standard deviation |
| SD (rat) | Sprague-Dawley (rat) |
| WNT5A-AS | wingless-type mouse mammary tumor virus integration site family member 5a-antisense |
References
- Tymofiyeva, O.; Hess, C.P.; Xu, D.; Barkovich, A.J. Structural MRI connectome in development: Challenges of the changing brain. Br. J. Radiol. 2014, 87. [Google Scholar] [CrossRef] [PubMed]
- Andropoulos, D.B. Effect of Anesthesia on the Developing Brain: Infant and Fetus. Fetal Diagn. Ther. 2018, 43, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hudson, A.E.; Hemmings, H.C. Are anaesthetics toxic to the brain? Br. J. Anaesth. 2011, 107, 30–37. [Google Scholar] [CrossRef]
- Sanders, R.D.; Hassell, J.; Davidson, A.J.; Robertson, N.J.; Ma, D. Impact of anaesthetics and surgery on neurodevelopment: An update. Br. J. Anaesth. 2013, 110, i53–i72. [Google Scholar] [CrossRef] [PubMed]
- Mellon, R.D.; Simone, A.F.; Rappaport, B.A. Use of Anesthetic Agents in Neonates and Young Children. Anesth. Analg. 2007, 104, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, B.A.; Suresh, S.; Hertz, S.; Evers, A.S.; Orser, B.A. Anesthetic Neurotoxicity—Clinical Implications of Animal Models. N. Engl. J. Med. 2015, 372, 796–797. [Google Scholar] [CrossRef]
- Bosnjak, Z.J.; Logan, S.; Liu, Y.A.; Bai, X.W. Recent Insights into Molecular Mechanisms of Propofol-Induced Developmental Neurotoxicity: Implications for the Protective Strategies. Anesth. Analg. 2016, 123, 1286–1296. [Google Scholar] [CrossRef]
- Steen, P.A.; Michenfelder, J.D. Neurotoxicity of Anesthetics. Anesthesiology 1979, 50, 437–453. [Google Scholar] [CrossRef]
- Cattano, D.; Young, C.; Straiko, M.M.W.; Olney, J.W. Subanesthetic Doses of Propofol Induce Neuroapoptosis in the Infant Mouse Brain. Anesth. Analg. 2008, 106, 1712–1714. [Google Scholar] [CrossRef]
- Jevtovic-Todorovic, V.; Hartman, R.E.; Izumi, Y.; Benshoff, N.D.; Dikranian, K.; Zorumski, C.F.; Olney, J.W.; Wozniak, D.F. Early exposure to common anesthetic agents causes widespread neurodegeneration in the developing rat brain and persistent learning deficits. J. Neurosci. 2003, 23, 876–882. [Google Scholar] [CrossRef]
- Stratmann, G.; Sall, J.W.; May, L.D.V.; Bell, J.S.; Magnusson, K.R.; Rau, V.; Visrodia, K.H.; Alvi, R.S.; Ku, B.; Lee, M.T.; et al. Isoflurane Differentially Affects Neurogenesis and Long-term Neurocognitive Function in 60-day-old and 7-day-old Rats. Anesthesiology 2009, 110, 834–848. [Google Scholar] [CrossRef] [PubMed]
- Lunardi, N.; Ori, C.; Erisir, A.; Jevtovic-Todorovic, V. General Anesthesia Causes Long-Lasting Disturbances in the Ultrastructural Properties of Developing Synapses in Young Rats. Neurotox. Res. 2009, 17, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Stachnik, J. Inhaled anesthetic agents. Am. J. Health Pharm. 2006, 63, 623–634. [Google Scholar] [CrossRef]
- Satomoto, M.; Satoh, Y.; Terui, K.; Miyao, H.; Takishima, K.; Ito, M.; Imaki, J. Neonatal Exposure to Sevoflurane Induces Abnormal Social Behaviors and Deficits in Fear Conditioning in Mice. Anesthesiology 2009, 110, 628–637. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, L.; Wang, C.; Tang, X.; Chen, Y.; Wang, X.; Su, L.; Hu, N.; Xie, K.; Yu, Y.; et al. Sevoflurane-induced learning deficits and spine loss via nectin-1/corticotrophin-releasing hormone receptor type 1 signaling. Brain Res. 2019, 1710, 188–198. [Google Scholar] [CrossRef]
- Fang, F.; Xue, Z.; Cang, J. Sevoflurane exposure in 7-day-old rats affects neurogenesis, neurodegeneration and neurocognitive function. Neurosci. Bull. 2012, 28, 499–508. [Google Scholar] [CrossRef]
- Shen, X.; Liu, Y.; Xu, S.; Zhao, Q.; Guo, X.; Shen, R.; Wang, F. Early life exposure to sevoflurane impairs adulthood spatial memory in the rat. NeuroToxicology 2013, 39, 45–56. [Google Scholar] [CrossRef]
- Diaz, L.K.; Gaynor, J.W.; Koh, S.J.; Ittenbach, R.F.; Gerdes, M.; Bernbaum, J.C.; Zackai, E.H.; Clancy, R.R.; Rehman, M.A.; Pennington, J.W.; et al. Increasing cumulative exposure to volatile anesthetic agents is associated with poorer neurodevelopmental outcomes in children with hypoplastic left heart syndrome. J. Thorac. Cardiovasc. Surg. 2016, 152, 482–489. [Google Scholar] [CrossRef]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [CrossRef]
- Riva, P.; Ratti, A.; Venturin, M. The Long Non-Coding RNAs in Neurodegenerative Diseases: Novel Mechanisms of Pathogenesis. Curr. Alzheimer Res. 2016, 13, 1219–1231. [Google Scholar] [CrossRef]
- Zhang, L.; Yan, J.; Liu, Q.D.; Xie, Z.C.; Jiang, H. LncRNA Rik-203 contributes to anesthesia neurotoxicity via microRNA-101a-3p and GSK-3 beta-mediated neural differentiation. Sci. Rep. 2019, 9, 1–12. [Google Scholar]
- Lerman, J.; Sikich, N.; Kleinman, S.; Yentis, S. The Pharmacology of Sevoflurane in Infants and Children. Anesthesiology 1994, 80, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Stefan De Hert, A.M. Sevoflurane. F1000Research 2015, 4, 1–8. [Google Scholar]
- Carmel, J.B.; Eyoung, W.; Hart, R.P. Flipping the transcriptional switch from myelin inhibition to axon growth in the CNS. Front. Mol. Neurosci. 2015, 8, 34. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Loh, S.H.Y.; Francescut, L.; Lingor, P.; Bähr, M.; Nicotera, P. Identification of new kinase clusters required for neurite outgrowth and retraction by a loss-of-function RNA interference screen. Cell Death Differ. 2007, 15, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Bai, J.; Zhang, X.; Lv, Y.; Gong, Y.; Liu, L.; Zhao, H.; Yu, F.; Ping, Y.; Zhang, G.; et al. A comprehensive overview of lncRNA annotation resources. Brief. Bioinform. 2016, 18, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, M.; Pilpel, Y. Genome-wide natural antisense transcription: Coupling its regulation to its different regulatory mechanisms. EMBO Rep. 2006, 7, 1216–1222. [Google Scholar] [CrossRef]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 1–13. [Google Scholar] [CrossRef]
- Han, X.D.; Li, M.; Zhang, X.G.; Xue, Z.G.; Cang, J. Single sevoflurane exposure increases methyl-CpG island binding protein 2 phosphorylation in the hippocampus of developing mice. Mol. Med. Rep. 2015, 11, 226–230. [Google Scholar] [CrossRef]
- Tang, X.; Zhao, Y.; Zhou, Z.; Yan, J.; Zhou, B.; Chi, X.; Luo, A.; Li, S.Y. Resveratrol Mitigates Sevoflurane-Induced Neurotoxicity by the SIRT1-Dependent Regulation of BDNF Expression in Developing Mice. Oxidative Med. Cell. Longev. 2020, 2020, 1–18. [Google Scholar] [CrossRef]
- Ge, X.; Zhang, Y.; Zuo, Y.; Israr, M.; Li, B.; Yu, P.; Gao, G.; Chang, Y.-Z.; Shi, Z. Transcriptomic analysis reveals the molecular mechanism of Alzheimer-related neuropathology induced by sevoflurane in mice. J. Cell. Biochem. 2019, 120, 17555–17565. [Google Scholar] [CrossRef]
- Bi, C.; Cai, Q.; Shan, Y.; Yang, F.; Sun, S.; Wu, X.; Liu, H. Sevoflurane induces neurotoxicity in the developing rat hippocampus by upregulating connexin 43 via the JNK/c-Jun/AP-1 pathway. Biomed. Pharmacother. 2018, 108, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xue, Z.; Sun, A. Subclinical concentration of sevoflurane potentiates neuronal apoptosis in the developing C57BL/6 mouse brain. Neurosci. Lett. 2008, 447, 109–114. [Google Scholar] [CrossRef]
- Yu, D.; Li, L.; Yuan, W. Neonatal anesthetic neurotoxicity: Insight into the molecular mechanisms of long-term neurocognitive deficits. Biomed. Pharmacother. 2017, 87, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Wang, X.; Youmans, D.T.; Cech, T.R. How do lncRNAs regulate transcription? Sci. Adv. 2017, 3, eaao2110. [Google Scholar] [CrossRef] [PubMed]
- Logan, S.; Jiang, C.; Yan, Y.; Inagaki, Y.; Arzua, T.; Bai, X. Propofol Alters Long Non-Coding RNA Profiles in the Neonatal Mouse Hippocampus: Implication of Novel Mechanisms in Anesthetic-Induced Developmental Neurotoxicity. Cell. Physiol. Biochem. 2018, 49, 2496–2510. [Google Scholar] [CrossRef] [PubMed]
- Hannila, S.S. Secretory Leukocyte Protease Inhibitor (SLPI): Emerging Roles in CNS Trauma and Repair. Neuroscientist 2015, 21, 630–636. [Google Scholar] [CrossRef]
- Lu, G.; Xu, H.; Zhao, W.; Zhang, J.; Rao, D.; Xu, S. Upregulation of long noncoding RNA Gadd45a is associated with sevoflurane-induced neurotoxicity in rat neural stem cells. NeuroReport 2018, 29, 605–614. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, X.; Lu, D.; Yang, X.; Zhou, Z.; Chen, X.; Chen, Y.; He, W.; Feng, X. Aberrantly expressed long noncoding RNAs are involved in sevoflurane-induced developing hippocampal neuronal apoptosis: A microarray related study. Metab. Brain Dis. 2016, 31, 1031–1040. [Google Scholar] [CrossRef]
- Zhao, Y.; Ai, Y. Overexpression of lncRNA Gm15621 alleviates apoptosis and inflammation response resulting from sevoflurane treatment through inhibiting miR-133a/Sox4. J. Cell. Physiol. 2020, 235, 957–965. [Google Scholar] [CrossRef]
- Liao, Q.; Liu, C.; Yuan, X.; Kang, S.; Miao, R.; Xiao, H.; Zhao, G.; Luo, H.; Bu, D.; Zhao, H.; et al. Large-scale prediction of long non-coding RNA functions in a coding-non-coding gene co-expression network. Nucleic Acids Res. 2011, 39, 3864–3878. [Google Scholar] [CrossRef]
- Yan, Y.; Qiao, S.; Kikuchi, C.; Zaja, I.; Logan, S.; Jiang, C.; Arzua, T.; Bai, X. Propofol Induces Apoptosis of Neurons but Not Astrocytes, Oligodendrocytes, or Neural Stem Cells in the Neonatal Mouse Hippocampus. Brain Sci. 2017, 7, 130. [Google Scholar] [CrossRef]
- Twaroski, D.M.; Yan, Y.; Olson, J.M.; Bosnjak, Z.J.; Bai, X. Down-regulation of microRNA-21 is involved in the propofol-induced neurotoxicity observed in human stem cell-derived neurons. Anesthesiology 2014, 121, 786–800. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]






| lncRNA Symbol | lncRNA Strand | lncRNA | mRNA Symbol | mRNA Strand | Nearby Coding mRNA | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| p-Value 1 | Fold Change | Regulation | FDR | p-Value | Fold Change | Regulation | FDR | ||||
| Gm16287 | − | 0.00024 | 2.4 | up | 0.099 | Ubr4 | + | 0.042 | 2.1 | down | 0.32 |
| Ak048274 | + | 0.014 | 2.4 | up | 0.22 | Igfbp5 | - | 0.011 | 2.3 | down | 0.24 |
| Ak081961 | + | 0.000038 | 6.5 | up | 0.057 | Igfbp5 | - | 0.011 | 2.3 | down | 0.24 |
| Ak137249 | − | 0.0019 | 2.9 | up | 0.14 | Mall | - | 0.00013 | 2.5 | up | 0.078 |
| Ak138713 | − | 0.00093 | 2.3 | up | 0.12 | Xlr5b | + | 0.027 | 2.2 | up | 0.28 |
| A730032A03Rik | + | 0.039 | 2.9 | down | 0.28 | Slpi | - | 0.0018 | 3.4 | up | 0.17 |
| Diseases or Functions Annotation | p-Value | mRNAs | Highly Correlated Co-Expressed lncRNAs |
|---|---|---|---|
| Apoptosis of granule cells | 0.00000606 | ADCYAP1, FAS, FRAT1, JUN, MECP2 | ENSMUST00000153752, uc012fts.1, AK084576, TCONS_00020793, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc007guo.1, uc012fts.1, ENSMUST00000155268, uc009mzl.1, mouselincRNA0733-, TCONS_00000533, ENSMUST00000117770, AK084576, AK032934, NR_024326, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Epileptic seizure | 0.000033 | ADCYAP1, CCR1, DUSP5, EGR4, HAS3, HSPB3, JUN, NR4A3 | ENSMUST00000153752, uc012fts.1,AK084576, ENSMUST00000145890, AK045892, NR_046157, AK131720, AK167175, ENSMUST00000134436, NR_024257, NR_036459, AK131720, uc009alp.2, ENSMUST00000126693, AK045957, AK086925, AV471140, ENSMUST00000155363, AK036595, AK014666, AK167175, ENSMUST00000145890, ENSMUST00000137546, AK045892, uc007uqa.1, NR_045190, NR_024257, ENSMUST00000169975, AK143771, ENSMUST00000137546, ENSMUST00000172524, uc007guo.1, uc012fts.1, mouselincRNA0733-, ENSMUST00000117770, AK084576, NR_045190, ENSMUST00000169975, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc012fts.1, uc009mzl.1, TCONS_00000533 |
| Epilepsy or neurodevelopmental disorder | 0.0000338 | ADCYAP1, AHNAK, BRPF1, CCR1, DUSP5, EGR4, GLI3, HAS3, HSPB3, JUN, MECP2, NR4A3, SCN11A | ENSMUST00000153752, uc012fts.1, AK084576, ENSMUST00000134436, NR_024257, NR_036459, AK131720, uc009alp.2, ENSMUST00000169975, AK045957, AK086925, AK143771, AK036595, AK167175, ENSMUST00000145890, ENSMUST00000137546, AK138866, NR_102396, uc007guo.1, mouselincRNA0733-, AK084576, NR_045190, AK081961, AK030101, NR_024326, ENSMUST00000145890, AK045892, NR_046157, AK131720, AK167175, ENSMUST00000134436, NR_024257, NR_036459, AK131720, uc009alp.2, ENSMUST00000126693, AK045957, AK086925, AV471140, ENSMUST00000155363, AK036595, AK014666, AK167175, ENSMUST00000145890, ENSMUST00000137546, AK045892, uc007uqa.1, NR_045190, ENSMUST00000135739, uc007guo.1, NR_024257, ENSMUST00000169975, AK143771, ENSMUST00000137546, ENSMUST00000172524, uc007guo.1, uc012fts.1, mouselincRNA0733-, ENSMUST00000117770, AK084576, NR_045190, ENSMUST00000169975, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc012fts.1, uc009mzl.1, TCONS_00000533, AK149342 |
| Epilepsy | 0.000035 | ADCYAP1, CCR1, DUSP5, EGR4, GLI3, HAS3, HSPB3, JUN, MECP2, NR4A3, SCN11A | ENSMUST00000153752, uc012fts.1, AK084576, ENSMUST00000145890, AK045892, NR_046157, AK131720, AK167175, ENSMUST00000134436, NR_024257, NR_036459, AK131720, uc009alp.2, ENSMUST00000126693, AK045957, AK086925, AV471140, ENSMUST00000155363, AK036595, AK014666, AK167175, ENSMUST00000145890, ENSMUST00000137546, AK045892, uc007uqa.1, NR_045190, ENSMUST00000135739, uc007guo.1, NR_024257, ENSMUST00000169975, AK143771, ENSMUST00000137546, ENSMUST00000172524, uc007guo.1, uc012fts.1, mouselincRNA0733-, ENSMUST00000117770, AK084576, NR_045190, ENSMUST00000169975, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc012fts.1, uc009mzl.1, TCONS_00000533, AK149342 |
| Apoptosis of cerebellar granule cell | 0.000134 | FRAT1, JUN, MECP2 | NR_024326, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-,NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Cell death of cerebellar cortex cells | 0.000174 | FAS, FRAT1, JUN, MECP2 | TCONS_00020793, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc007guo.1, uc012fts.1, ENSMUST00000155268, uc009mzl.1, mouselincRNA0733-, TCONS_00000533, ENSMUST00000117770, AK084576, AK032934, NR_024326, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Apoptosis of brain cells | 0.00022 | ADCYAP1, ALOX15, FAS, FRAT1, JUN, MECP2 | ENSMUST00000153752, uc012fts.1, AK084576, AK131720, uc009alp.2, AK045957, AV471140, ENSMUST00000155363, AK036595, AK167175, AK081961, ENSMUST00000137546, mouselincRNA0733-, TCONS_00020793, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc007guo.1, uc012fts.1, ENSMUST00000155268, uc009mzl.1, mouselincRNA0733-, TCONS_00000533, ENSMUST00000117770, AK084576, AK032934, NR_024326, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Seizure disorder | 0.000386 | ADCYAP1, CCR1,DUSP5, EGR4, GLI3, HAS3, HSPB3, JUN, MECP2, NR4A3, SCN11A, ST8SIA4 | ENSMUST00000153752, uc012fts.1, AK084576, ENSMUST00000145890, AK045892, NR_046157, AK131720, AK167175, ENSMUST00000134436, NR_024257, NR_036459, AK131720, uc009alp.2, ENSMUST00000126693, AK045957, AK086925, AV471140, ENSMUST00000155363, AK036595, AK014666, AK167175, ENSMUST00000145890, ENSMUST00000137546, AK045892, uc007uqa.1, NR_045190, ENSMUST00000135739, uc007guo.1, NR_024257, ENSMUST00000169975, AK143771, ENSMUST00000137546, ENSMUST00000172524, uc007guo.1, uc012fts.1, mouselincRNA0733-, ENSMUST00000117770, AK084576, NR_045190, ENSMUST00000169975 AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc012fts.1, uc009mzl.1, TCONS_00000533, AK149342, ENSMUST00000118347, TCONS_00020794 |
| Seizures | 0.000487 | ADCYAP1, CCR1, DUSP5, EGR4, HAS3, HSPB3, JUN, MECP2, NR4A3, SCN11A, ST8SIA4 | ENSMUST00000153752, uc012fts.1, AK084576, ENSMUST00000145890, AK045892, NR_046157, AK131720, AK167175, ENSMUST00000134436, NR_024257, NR_036459, AK131720, uc009alp.2, ENSMUST00000126693, AK045957, AK086925, AV471140, ENSMUST00000155363, AK036595, AK014666, AK167175, ENSMUST00000145890, ENSMUST00000137546, AK045892, uc007uqa.1, NR_045190, NR_024257, ENSMUST00000169975, AK143771, ENSMUST00000137546, ENSMUST00000172524, uc007guo.1, uc012fts.1, mouselincRNA0733-, ENSMUST00000117770, AK084576, NR_045190, ENSMUST00000169975, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc012fts.1, uc009mzl.1, TCONS_00000533, AK149342, ENSMUST00000118347, TCONS_00020794 |
| Apoptosis of cortical neurons | 0.00153 | ALOX15, FAS, JUN, MECP2 | AK131720, uc009alp.2, AK045957, AV471140, ENSMUST00000155363, AK036595, AK167175, AK081961, ENSMUST00000137546, mouselincRNA0733-, TCONS_00020793, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc007guo.1, uc012fts.1, ENSMUST00000155268, uc009mzl.1, mouselincRNA0733-, TCONS_00000533, ENSMUST00000117770, AK084576, AK032934, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733-, NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Hereditary sensory and autonomic neuropathy type VII | 0.00479 | SCN11A | AK149342 |
| Zappella variant Rett syndrome | 0.00479 | MECP2 | NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Blepharophimosis, ptosis, and epicanthus inversus type II with Duane retraction syndrome | 0.00479 | FOXL2 | TCONS_00020793, TCONS_00020794, ENSMUST00000135739, ENSMUST00000172524, uc007guo.1, ENSMUST00000153752, uc012fts.1, ENSMUST00000155268, TCONS_00000533, AK032934 |
| Kallmann syndrome type 17 | 0.00479 | SPRY4 | ENSMUST00000134436, AK131720, uc009alp.2, ENSMUST00000169975, AK045957, AK086925, AK143771, AK036595, ENSMUST00000145890, ENSMUST00000137546, AK138866, uc007guo.1, uc012fts.1, mouselincRNA0733-, ENSMUST00000117770, AK084576, NR_045190 |
| Susceptibility to X-linked autism type 3 | 0.00479 | MECP2 | NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Distal hereditary motor neuronopathy type IIC | 0.00479 | HSPB3 | ENSMUST00000169975 |
| Susceptibility to hypogonadotropic hypogonadism type 17 | 0.00479 | SPRY4 | ENSMUST00000134436, AK131720, uc009alp.2, ENSMUST00000169975, AK045957, AK086925, AK143771, AK036595, ENSMUST00000145890, ENSMUST00000137546, AK138866, uc007guo.1, uc012fts.1, mouselincRNA0733-, ENSMUST00000117770, AK084576, NR_045190 |
| Neurodegeneration of sympathetic neuron | 0.00479 | JUN | AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Hypogonadotropic hypogonadism 17 without anosmia | 0.00479 | SPRY4 | ENSMUST00000134436, AK131720, uc009alp.2, ENSMUST00000169975, AK045957, AK086925, AK143771, AK036595, ENSMUST00000145890, ENSMUST00000137546, AK138866, uc007guo.1, uc012fts.1, mouselincRNA0733-, ENSMUST00000117770, AK084576, NR_045190 |
| X-linked mental retardation syndromic 13 | 0.00479 | MECP2 | NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Neonatal severe encephalopathy | 0.00479 | MECP2 | NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Intellectual developmental disorder with dysmorphic facies and ptosis | 0.00479 | BRPF1 | AK081961, AK030101, NR_024326 |
| Degeneration of cholinergic fibers | 0.00479 | JUN | AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Autosomal recessive intellectual developmental disorder 68 | 0.00479 | TRMT1 | AK143771, TCONS_00020794, uc012fts.1, ENSMUST00000155268, TCONS_00000533, ENSMUST00000117770, AK032934 |
| Familial episodic pain syndrome type 3 | 0.00479 | SCN11A | AK149342 |
| Lubs X-linked mental retardation syndrome | 0.00479 | MECP2 | NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Hypothalamic hamartoma | 0.00479 | GLI3 | ENSMUST00000135739, uc007guo.1 |
| Anosmia | 0.00908 | SLC7A11, SPRY4 | ENSMUST00000141521, ENSMUST00000135739, uc007guo.1, ENSMUST00000117770, AK032934, ENSMUST00000134436, AK131720, uc009alp.2, ENSMUST00000169975, AK045957, AK086925, AK143771, AK036595, ENSMUST00000145890, ENSMUST00000137546, AK138866, uc007guo.1, uc012fts.1, mouselincRNA0733-, ENSMUST00000117770, AK084576, NR_045190 |
| Lack of inferior colliculus | 0.00957 | GLI3 | ENSMUST00000135739, uc007guo.1 |
| Apoptosis of somatotrophs | 0.00957 | FAS | TCONS_00020793, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc007guo.1, uc012fts.1, ENSMUST00000155268, uc009mzl.1, mouselincRNA0733-, TCONS_00000533, ENSMUST00000117770, AK084576, AK032934 |
| Neurodegeneration of substantia nigra pars compacta | 0.00957 | SLC7A11 | ENSMUST00000141521, ENSMUST00000135739, uc007guo.1, ENSMUST00000117770, AK032934 |
| Familial Angelman syndrome | 0.00957 | MECP2 | NR_024257, AK131720, AV471140, AK143771, AK167175, AK081961, AK030101, ENSMUST00000137546, mouselincRNA0733- |
| Misrouting of thalamocortical axons | 0.00957 | ST8SIA4 | ENSMUST00000118347, TCONS_00020794 |
| Apoptosis of lactotropes | 0.00957 | FAS | TCONS_00020793, ENSMUST00000169975, AK143771, TCONS_00020794, ENSMUST00000137546, ENSMUST00000135739, ENSMUST00000172524, uc007guo.1, uc012fts.1, ENSMUST00000155268, uc009mzl.1, mouselincRNA0733-, TCONS_00000533, ENSMUST00000117770, AK084576, AK032934 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, C.; Arzua, T.; Yan, Y.; Bai, X. Expression Signature of lncRNAs and mRNAs in Sevoflurane-Induced Mouse Brain Injury: Implication of Involvement of Wide Molecular Networks and Pathways. Int. J. Mol. Sci. 2021, 22, 1389. https://doi.org/10.3390/ijms22031389
Jiang C, Arzua T, Yan Y, Bai X. Expression Signature of lncRNAs and mRNAs in Sevoflurane-Induced Mouse Brain Injury: Implication of Involvement of Wide Molecular Networks and Pathways. International Journal of Molecular Sciences. 2021; 22(3):1389. https://doi.org/10.3390/ijms22031389
Chicago/Turabian StyleJiang, Congshan, Thiago Arzua, Yasheng Yan, and Xiaowen Bai. 2021. "Expression Signature of lncRNAs and mRNAs in Sevoflurane-Induced Mouse Brain Injury: Implication of Involvement of Wide Molecular Networks and Pathways" International Journal of Molecular Sciences 22, no. 3: 1389. https://doi.org/10.3390/ijms22031389
APA StyleJiang, C., Arzua, T., Yan, Y., & Bai, X. (2021). Expression Signature of lncRNAs and mRNAs in Sevoflurane-Induced Mouse Brain Injury: Implication of Involvement of Wide Molecular Networks and Pathways. International Journal of Molecular Sciences, 22(3), 1389. https://doi.org/10.3390/ijms22031389