
The Role of Lipids in Legionella-Host Interaction


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Characteristics of Legionella Phospholipids
2.1. Phospholipids of Legionella spp.
2.2. Fatty Acid Composition of Legionella spp.
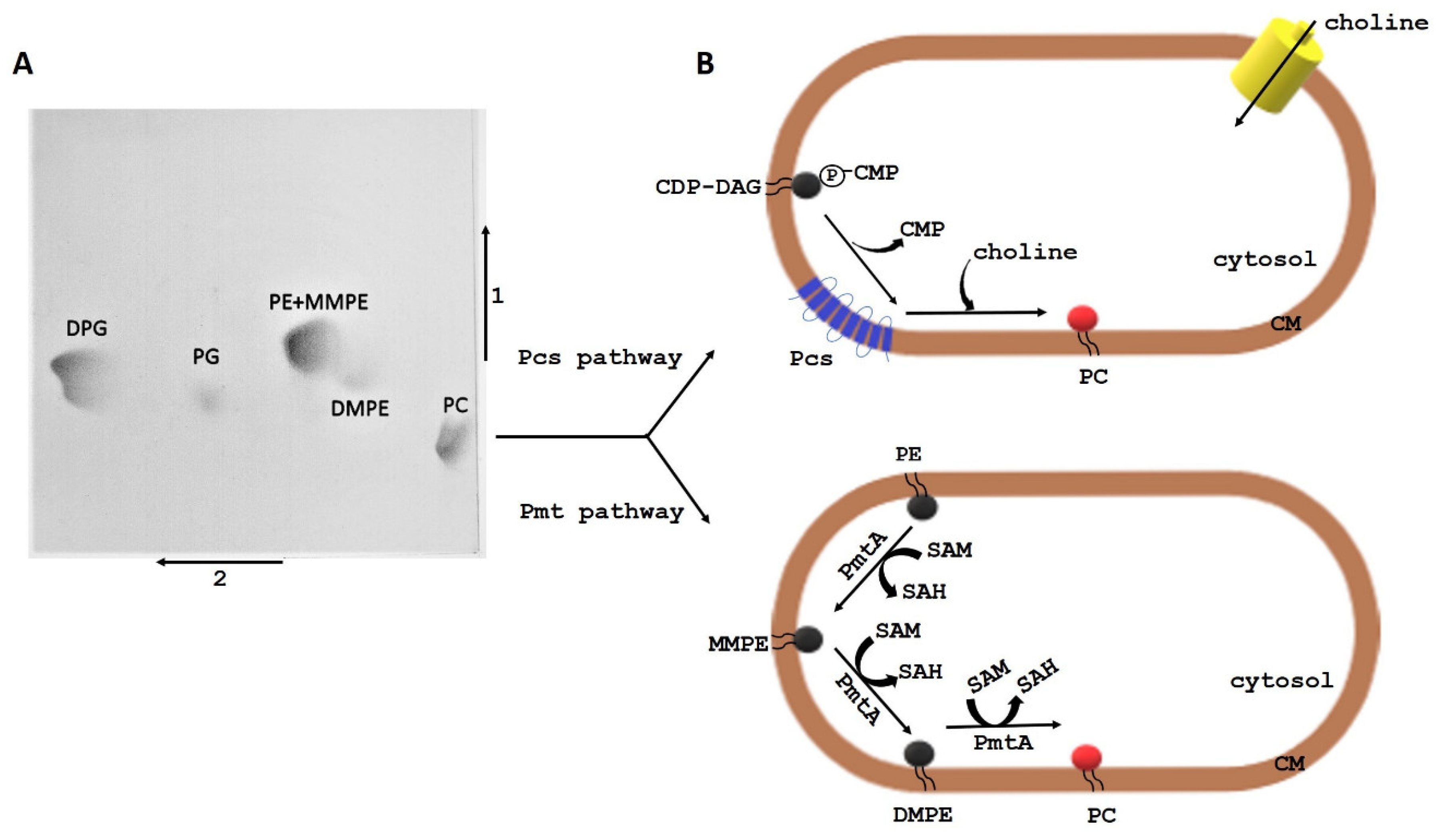
2.3. PC Synthesis Pathways
2.4. Genetic Diversity of Legionella pcs and pmtA Genes
2.5. Utilisation of Exogenous Choline for PC Synthesis by Legionella spp.
2.6. Biological Importance of Legionella PC
3. Structure and Significance of Legionella Lipopolysaccharide
3.1. Chemical Structure of Legionella LPS
3.2. Biological Significance of Legionella LPS
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fields, B.S.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ disease: 25 years of investigation. Clin. Microbiol. Rev. 2002, 15, 506–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaro, F.; Wang, W.; Gilbert, J.A.; Anderson, O.R.; Shuman, H.A. Diverse protist grazers select for virulence-related traits in Legionella. ISME J. 2015, 9, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Euzeby, J.P. List of Prokaryotic Names with Standing in Nomenclature Genus Legionella. Available online: https://www.bacterio.net/legionella.html (accessed on 27 January 2021).
- O’Connor, T.J.; Adepoju, Y.; Boyd, D.; Isberg, R.R. Minimization of the Legionella pneumophila genome reveals chromosomal regions involved in host range expansion. Proc. Natl. Acad. Sci. USA 2011, 108, 14733–14740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaheen, M.; Ashbolt, N.J. Differential bacterial predation by free-living amoebae may result in blooms of Legionella in drinking water systems. Microorganisms 2021, 15, 174. [Google Scholar] [CrossRef] [PubMed]
- Brieland, J.; McClain, M.; LeGendre, M.; Engleberg, C. Intrapulmonary Hartmannella vermiformis: A potential niche for Legionella pneumophila replication in a murine model of legionellosis. Infect. Immun. 1997, 65, 4892–4896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Personnic, N.; Striednig, B.; Lezan, E.; Manske, C.; Welin, A.; Schmidt, A.; Hilbi, H. Quorum sensing modulates the formation of virulent Legionella persisters within infected cells. Nat. Commun. 2019, 10, 5216. [Google Scholar] [CrossRef] [Green Version]
- Brieland, J.; McClain, M.; Heath, L.; Chrisp, C.; Huffnagle, G.; LeGendre, M.; Hurley, M.; Fantone, J.C.; Engleberg, C. Coinoculation with Hartmanella vermiformis enhances replicative Legionella pneumophila lung infection in a murine model of Legionnaires’ disease. Infect. Immun. 1996, 64, 2449–2456. [Google Scholar] [CrossRef] [Green Version]
- Al-Quadan, T.; Price, C.T.; Kwaik, Y.A. Exploitation of evolutionarily conserved amoeba and mammalian processes by Legionella. Trends Microbiol. 2012, 20, 299–306. [Google Scholar] [CrossRef] [Green Version]
- De Giglio, O.; Fasano, F.; Diella, G.; Lopuzzo, M.; Napoli, C.; Apollonio, F.; Brigida, S.; Calia, C.; Campanale, C.; Marzella, A.; et al. Legionella and legionellosis in touristic-recreational facilities: Influence of climate factors and geostatistical analysis in Southern Italy (2001–2017). Environ. Res. 2019, 178, 108721. [Google Scholar] [CrossRef]
- De Giglio, O.; Napoli, C.; Apollonio, F.; Brigida, S.; Marzella, A.; Diella, G.; Calia, C.; Scrascia, M.; Pacifico, C.; Pazzani, C.; et al. Occurrence of Legionella in groundwater used for sprinkler irrigation in Southern Italy. Environ. Res. 2019, 170, 215–221. [Google Scholar] [CrossRef]
- Correia, A.M.; Ferreira, J.S.; Borges, V.; Nunes, A.; Gomes, B.; Capucho, R.; Gonçalves, J.; Antunes, D.M.; Almeida, S.; Mendes, A.; et al. Probable person-to-person transmission of Legionnaires’ disease. N. Engl. J. Med. 2016, 374, 497–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, B.A.; Burillo, A.; Bouza, E. Legionnaires’ disease. Lancet 2016, 387, 376–385. [Google Scholar] [CrossRef]
- Portlock, T.J.; Tyson, J.Y.; Dantu, S.C.; Rehman, S.; White, R.C.; McIntire, I.E.; Sewell, L.; Richardson, K.; Shaw, R.; Pandini, A.; et al. Structure, dynamics and cellular insight into novel substrates of the Legionella pneumophila type II secretion system. Front. Mol. Biosci. 2020, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Bärlocher, K.; Welin, A.; Hilbi, H. Formation of the Legionella replicative compartment at the crossroads of retrograde trafficking. Front. Cell. Infect. Microbiol. 2017, 7, 482. [Google Scholar] [CrossRef] [PubMed]
- Shevchuk, O.; Jäger, J.; Steinert, M. Virulence properties of the Legionella pneumophila cell envelope. Front. Microbiol. 2011, 25, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caroff, M.; Novikov, A. LPS structure, function, and heterogeneity. In Endotoxin Detection and Control in Pharma, Limulus, and Mammalian Systems; Williams, K.L., Ed.; Springer Nature: Berlin/Heidelberg, Germany, 2019; pp. 53–93. [Google Scholar] [CrossRef]
- Bohdanowich, M.; Grinstein, S. Role of phospholipids in endocytosis, phagocytosis, and macropinocytosis. Physiol. Rev. 2013, 93, 69–106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef]
- Geiger, O. Biogenesis of Fatty Acids, Lipids and Membranes; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–886. [Google Scholar] [CrossRef]
- Finnerty, W.R.; Makula, R.A.; Feeley, J.C. Cellular lipids of the Legionnaires’ disease bacterium. Ann. Intern. Med. 1979, 90, 631–634. [Google Scholar] [CrossRef]
- Palusinska-Szysz, M.; Kalityński, R.; Russa, R.; Dawidowicz, A.L.; Drożański, W.J. Cellular envelope phospholipids from Legionella lytica. FEMS Microbiol. Lett. 2008, 283, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Palusinska-Szysz, M.; Janczarek, M.; Kalityński, R.; Dawidowicz, A.L.; Russa, R. Legionella bozemanae synthesizes phosphatidylcholine from exogenous choline. Microbiol. Res. 2011, 166, 87–98. [Google Scholar] [CrossRef]
- Palusinska-Szysz, M.; Szuster-Ciesielska, A.; Kania, M.; Janczarek, M.; Chmiel, E.; Danikiewicz, W. Legionella dumoffii utilizes exogenous choline for phosphatidylcholine synthesis. Int. J. Mol. Sci. 2014, 15, 8256–8279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palusinska-Szysz, M.; Szuster-Ciesielska, A.; Janczarek, M.; Wdowiak-Wróbel, S.; Schiller, J.; Reszczyńska, E.; Gruszecki, W.I.; Fuchs, B. Genetic diversity of Legionella pcs and pmtA genes and the effect of utilization of choline by Legionella spp. on induction of proinflammatory cytokines. Pathog. Dis. 2019, 77, ftz065. [Google Scholar] [CrossRef] [PubMed]
- Hindahl, M.S.; Iglewski, B.H. Isolation and characterization of the Legionella pneumophila outer membrane. J. Bacteriol. 1984, 159, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, M.A.; Moss, C.W. Cellular fatty acid compositions and isoprenoid quinine contents of 23 Legionella species. J. Clin. Microbiol. 1989, 27, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Palusinska-Szysz, M.; Zdybicka-Barabas, A.; Reszczyńska, E.; Luchowski, R.; Kania, M.; Gisch, N.; Waldow, F.; Mak, P.; Danikiewicz, W.; Gruszecki, W.I.; et al. The lipid composition of Legionella dumoffii membrane modulates the interaction with Galleria mellonella apolipophorin III. Biochim. Biophys. Acta 2016, 1861, 617–629. [Google Scholar] [CrossRef]
- Verdon, J.; Labanowski, J.; Sahar, T.; Ferreira, T.; Lacombe, C.; Buchrieser, C.; Berjeaud, J.M.; Hechard, Y. Fatty acid composition modulates sensitivity of Legionella pneumophila to warnericin RK, an antimicrobial peptide. Biochim. Biophys. Acta 2011, 1808, 1146–1153. [Google Scholar] [CrossRef]
- López-Lara, I.M.; Geiger, O. Novel pathway for phosphatidylcholine biosynthesis in bacteria associated with eukaryotes. J. Biotechnol. 2001, 91, 211–221. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; López-Lara, M.I.; Geiger, O. Biosynthesis of phosphatidylcholine in bacteria. Prog. Lipid Res. 2003, 42, 115–162. [Google Scholar] [CrossRef]
- de Rudder, K.E.E.; Sohlenkamp, C.; Geiger, O. Plant-exudated choline is used for rhizobial membrane lipid biosynthesis by phosphatidylcholine synthase. J. Biol. Chem. 1999, 274, 20011–20016. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Morales, F.; Schobert, M.; Lopez-Lara, I.M.; Geiger, O. Pathways for phosphatidylcholine biosynthesis in bacteria. Microbiology 2003, 149, 3461–3471. [Google Scholar] [CrossRef]
- Gomez-Valero, L.; Rusniok, C.; Rolando, M.; Neou, M.; Dervins-Ravault, D.; Demirtas, J.; Rouy, Z.; Moore, R.J.; Chen, H.; Petty, N.K.; et al. Comparative analyses of Legionella species identifies genetic features of strains causing Legionnaires’ disease. Genome Biol. 2014, 15, 505. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.G.; McMaster, C.R. Scanning alanine mutagenesis of the CDP-alcohol phosphotransferase motif of Saccharomyces cerevisiae cholinephosphotransferase. J. Biol. Chem. 1998, 273, 13482–13487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Rudder, K.E.; López-Lara, I.M.; Geiger, O. Inactivation of the gene for phospholipid N-methyltransferase in Sinorhizobium meliloti: Phosphatidylcholine is required for normal growth. Mol. Microbiol. 2000, 37, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Arondel, V.; Benning, C.; Somerville, C. Isolation and functional expression in Escherichia coli of a gene encoding phosphatidylethanolamine methyltransferase (EC 2.1.1.17) from Rhodobacter sphaeroides. J. Biol. Chem. 1993, 268, 16002–16008. [Google Scholar] [CrossRef]
- Janczarek, M.; Palusinska-Szysz, M. PCR method for the rapid detection and discrimination of Legionella spp. based on the amplification of pcs, pmtA, and 16S rRNA genes. J. Appl. Genet. 2016, 57, 251–261. [Google Scholar] [CrossRef]
- Conover, G.M.; Martinez-Morales, F.; Heidtman, M.I.; Luo, Z.Q.; Tang, M.; Chen, C.; Geiger, O.; Isberg, R.R. Phosphatidylcholine synthesis required for optimal function of Legionella pneumophila virulence determinants. Cell. Microbiol. 2008, 10, 514–528. [Google Scholar] [CrossRef] [Green Version]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 1, 133–159. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Kang, Y.; Norris, M.H.; Troyer, R.M.; Son, M.S.; Schweizer, H.P.; Dow, S.W.; Hoang, T.T. Blocking phosphatidylcholine utilization in Pseudomonas aeruginosa, via mutagenesis of fatty acid, glycerol and choline degradation pathways, confirms the importance of this nutrient source in vivo. PLoS ONE 2014, 7, e103778. [Google Scholar] [CrossRef] [Green Version]
- Kondakova, T.; D’Heygère, F.; Feuilloley, M.J.; Orange, N.; Heipieper, H.J.; Duclairoir Poc, C. Glycerophospholipid synthesis and functions in Pseudomonas. Chem. Phys. Lipids 2015, 190, 27–42. [Google Scholar] [CrossRef]
- Fozo, E.M.; Rucks, E.A. The making and taking of lipids: The role of bacterial lipid synthesis and the harnessing of host lipids in bacterial pathogenesis. Adv. Microb. Physiol. 2016, 69, 51–155. [Google Scholar] [CrossRef]
- Zhang, Y.; Lewis, R.N.; McElhaney, R.N.; Ryan, R.O. Calorimetric and spectroscopic studies of the interaction of Manduca Sexta apolipophorin III with zwitterionic, anionic, and nonionic lipids. Biochemistry 1993, 32, 3942–3952. [Google Scholar] [CrossRef] [PubMed]
- Palusińska-Szysz, M.; Zdybicka-Barabas, A.; Luchowski, R.; Reszczyńska, E.; Śmiałek, J.; Mak, P.; Gruszecki, W.I.; Cytryńska, C. Choline supplementation sensitizes Legionella dumoffii to Galleria mellonella apolipophorin III. Int. J. Mol. Sci. 2020, 21, 5818. [Google Scholar] [CrossRef] [PubMed]
- Palusińska-Szysz, M.; Zdybicka-Barabas, A.; Pawlikowska-Pawlęga, B.; Mak, P.; Cytryńska, M. Anti-Legionella dumoffii activity of Galleria mellonella defensin and apolipophorin III. Int. J. Mol. Sci. 2012, 13, 17048–17064. [Google Scholar] [CrossRef] [Green Version]
- Richter, E.; Ventz, K.; Harms, M.; Mostertz, J.; Hochgräfe, F. Induction of macrophage function in human THP-1 cells is associated with rewiring of MAPK signaling and activation of MAP3K7 (TAK1) protein kinase. Front. Cell Dev. Biol. 2016, 4, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Campbell, H.A.; King, S.C.; Dowhan, W. Phospholipids as determinants of membrane protein topology. Phosphatidylethanolamine is required for the proper topological organization of the γ-aminobutyric acid permease (GabP) of Escherichia coli. J. Biol. Chem. 2005, 280, 26032–26038. [Google Scholar] [CrossRef] [Green Version]
- Dowhan, W. Molecular basis for membrane phospholipid diversity: Why are there so many lipids? Annu. Rev. Biochem. 1997, 66, 199–232. [Google Scholar] [CrossRef] [Green Version]
- Knirel, Y.A.; Rietschel, E.T.; Marre, R.; Zähringer, U. The structure of the O-specific chain of Legionella pneumophila serogroup 1 lipopolysaccharide. Eur. J. Biochem. 1994, 221, 239–245. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Moll, H.; Zähringer, U. Structural study of a highly O-acetylated core of Legionella pneumophila serogroup 1 lipopolysaccharide. Carbohydr. Res. 1996, 293, 223–234. [Google Scholar] [CrossRef]
- Moll, H.; Knirel, A.Y.; Helbig, H.J.; Zähringer, U. Identification of an ɑ-d-Manp-(1→8)-Kdo disaccharide in the inner core region and the structure of the complete core region of the Legionella pneumophila serogroup 1 lipopolysaccharide. Carbohydr. Res. 1997, 304, 91–95. [Google Scholar] [CrossRef]
- Moll, H.; Sonesson, A.; Jantzen, E.; Marre, R.; Zähringer, U. Identification of 27-oxo-octacosanoic acid and heptacosane-1,27-dioic acid in Legionella pneumophila. FEMS Microbiol. Lett. 1992, 76, 1–6. [Google Scholar] [CrossRef]
- Zähringer, U.; Knirel, A.Y.; Lindner, B.; Helbig, J.H.; Sonesson, A.; Marre, R.; Rietschel, E.T. The lipopolysaccharide of Legionella pneumophila (strain Philadelphia 1): Chemical structure and biological significance. Prog. Clin. Biol. Res. 1995, 392, 113–139. [Google Scholar] [PubMed]
- Sonesson, A.; Jantzen, E.; Bryn, K.; Larsson, L.; Eng, J. Chemical composition of a lipopolysaccharide from Legionella pneumophila. Arch. Microbiol. 1989, 153, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Sonesson, A.; Jantzen, E.; Tangen, T.; Zähringer, U. Chemical composition of lipopolysaccharides from Legionella bozemanii and Legionella longbeachae. Arch. Microbiol. 1994, 162, 215–221. [Google Scholar] [CrossRef]
- Sonesson, A.; Jantzen, E.; Bryn, K.; Tangen, T.; Eng, J.; Zähringer, U. Composition of 2,3-dihydroxy fatty acids-containing lipopolysaccharides from Legionella israelensis, Legionella maceachernii and Legionella micdadei. Microbiology 1994, 140, 1261–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palusinska-Szysz, M.; Russa, R. Chemical structure and biological significance of lipopolysaccharide from Legionella. Recent Pat. Anti-Infect. Drug Discov. 2009, 4, 96–107. [Google Scholar] [CrossRef]
- Sonesson, A.; Jantzen, A. The branched-chain octose yersiniose A is a lipopolysaccharide constituent of Legionella micdadei and Legionella maceachernii. J. Microbiol. Methods 1992, 15, 241–248. [Google Scholar] [CrossRef]
- Sonesson, A.; Jantzen, E.; Tangen, T.; Zähringer, U. Chemical characterization of lipopolysaccharides from Legionella feeleii, Legionella hackeliae and Legionella jordanis. Microbiology 1994, 140, 2663–2671. [Google Scholar] [CrossRef] [Green Version]
- Sonesson, A.; Jantzen, E.; Tangen, T.; Zähringer, U. Lipopolysaccharides of Legionella erythra and Legionella oakridgensis. Can. J. Microbiol. 1994, 40, 666–671. [Google Scholar] [CrossRef]
- Ciesielski, C.A.; Blaser, M.J.; Wang, W.L. Serogroup specificity of Legionella pneumophila is related to lipopolysaccharide characteristics. Infect. Immun. 1986, 51, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Helbig, J.H.; Jacobs, E.; Lück, C. Legionella pneumophila urinary antigen subtyping using monoclonal antibodies as a tool for epidemiological investigations. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1673–1677. [Google Scholar] [CrossRef]
- Joly, J.R.; McKinney, R.M.; Tobin, J.O.; Bibb, W.F.; Watkins, I.D.; Ramsay, D. Development of a standardized subgrouping scheme for Legionella pneumophila serogroup 1 using monoclonal antibodies. J. Clin. Microbiol. 1986, 23, 768–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petzold, M.; Thürmer, A.; Menzel, S.; Mouton, J.W.; Heuner, K.; Lück, C. A structural comparison of lipopolysaccharide biosynthesis loci of Legionella pneumophila serogroup 1 strains. BMC Microbiol. 2013, 13, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, H.C.; Knirel, Y.A.; Helbig, H.J.; Zähringer, U.; Mintz, C.S. Molecular cloning and characterization of a locus responsible for O-acetylation of the O polysaccharide of Legionella pneumophila serogroup 1 lipopolysaccharide. J. Bacteriol. 1999, 181, 4137–4141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helbig, J.H.; Lück, P.C.; Knirel, Y.A.; Witzleb, W.; Zähringer, U. Molecular characterization of a virulence-associated epitope on the lipopolysaccharide of Legionella pneumophila serogroup 1. Epidemiol. Infect. 1995, 115, 71–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Amemura-Maekawa, J.; Ren, H.; Li, Y.; Sakata, M.; Zhou, H.; Murai, M.; Chang, B.; Ohnishi, M.; Qin, T. Distribution of lag-1 alleles, ORF7, and ORF8 genes of lipopolysaccharide and sequence-based types among Legionella pneumophila serogroup 1 isolates in Japan and China. Front. Cell. Infect. Microbiol. 2019, 9, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palusinska-Szysz, M.; Luchowski, R.; Gruszecki, W.I.; Choma, A.; Szuster-Ciesielska, A.; Lück, C.; Petzold, M.; Sroka-Bartnicka, A.; Kowalczyk, B. The role of Legionella pneumophila serogroup 1 lipopolysaccharide in host-pathogen interaction. Front. Microbiol. 2019, 10, 2890. [Google Scholar] [CrossRef]
- Lück, P.C.; Freier, T.; Steudel, C.; Knirel, Y.A.; Lûneberg, E.; Zähringer, U.; Helbig, J.H. A point mutation in the active site of Legionella pneumophila O-acetyltransferase results in modified lipopolysaccharide but does not influence virulence. Int. J. Med. Microbiol. 2001, 291, 345–352. [Google Scholar] [CrossRef]
- Jäger, J.; Keese, S.; Roessle, M.; Steinert, M.; Schromm, A.B. Fusion of Legionella pneumophila outer membrane vesicles with eukaryotic membrane systems is a mechanism to deliver pathogen factors to host cell membranes. Cell. Microbiol. 2015, 5, 607–620. [Google Scholar] [CrossRef]
- Helbig, J.H.; Fernandez-Moreira, E.; Jacobs, E.; Luck, P.C.; Witt, M. Lipopolysaccharide architecture of Legionella pneumophila grown in broth and host cells. In Legionella: State of the Art 30 Years After its Recognition; Cianciotto, N.P., Ed.; ASM Press: Washington, DC, USA, 2006; pp. 261–264. [Google Scholar]
- Fernandez-Moreira, E.; Helbig, J.H.; Swanson, M.S. Membrane vesicles shed by Legionella pneumophila inhibit fusion of phagosomes with lysosomes. Infect. Immun. 2006, 74, 3285–3295. [Google Scholar] [CrossRef] [Green Version]
- Seeger, E.M.; Thuma, M.; Fernandez-Moreira, E.; Jacobs, E.; Schmitz, M.; Helbig, J.H. Lipopolysaccharide of Legionella pneumophila shed in a liquid culture as a nonvesicular fraction arrests phagosome maturation in amoeba and monocytic host cells. FEMS Microbiol. Lett. 2010, 307, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S. Sialic acid and biology of life: An introduction. In Sialic Acids and Sialoglycoconjugates in the Biology of Life, Health and Disease, 1st ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 1–61. [Google Scholar] [CrossRef]
- Neumeister, B.; Faigle, M.; Sommer, M.; Zähringer, U.; Stelter, F.; Menzel, R.; Schütt, C.; Northoff, H. Low endotoxic potential of Legionella pneumophila lipopolysaccharide due to failure of interaction with the monocyte lipopolysaccharide receptor CD14. Infect. Immun. 1998, 66, 4151–4157. [Google Scholar] [CrossRef] [PubMed]
- Girard, R.; Pedron, T.; Uematsu, S.; Balloy, V.; Chignard, M.; Akira, S.; Chaby, R. Lipopolysaccharides from Legionella and Rhizobium stimulate mouse bone marrow granulocytes via Toll-like receptor 2. J. Cell Sci. 2003, 116, 293–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lettinga, K.D.; Florquin, S.; Speelman, P.; van Ketel, R.; van der Poll, T.; Verbon, A. Toll-like receptor 4 is not involved in host defense against pulmonary Legionella pneumophila infection in a mouse model. J. Infect. Dis. 2002, 186, 570–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archer, K.A.; Roy, C.R. MyD88-dependent responses involving toll-like receptor 2 are important for protection and clearance of Legionella pneumophila in a mouse model of Legionnaires’ disease. Infect. Immun. 2006, 74, 3325–3333. [Google Scholar] [CrossRef] [Green Version]
- Hawn, T.R.; Smith, K.D.; Aderem, A.; Skerrett, S.J. Myeloid differentiation primary response gene (88)-and toll-like receptor 2-deficient mice are susceptible to infection with aerosolized Legionella pneumophila. J. Infect. Dis. 2006, 193, 1693–1702. [Google Scholar] [CrossRef] [Green Version]
- Archer, K.A.; Alexopoulou, L.; Flavell, R.A.; Roy, C.R. Multiple MyD88-dependent responses contribute to pulmonary clearance of Legionella pneumophila. Cell. Microbiol. 2009, 11, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Mallama, C.A.; McCoy-Simandle, K.; Cianciotto, N.P. The type II secretion system of Legionella pneumophila dampens the MyD88 and Toll-like receptor 2 signaling pathway in infected human macrophages. Infect. Immun. 2017, 85, e00897-16. [Google Scholar] [CrossRef] [Green Version]
- Lüneberg, E.; Zähringer, U.; Knirel, Y.A.; Steinmann, D.; Hartmann, M.; Steinmetz, I.; Rohde, M.; Köhl, J.; Frosch, M. Phase-variable expression of lipopolysaccharide contributes to the virulence of Legionella pneumophila. J. Exp. Med. 1998, 188, 49–60. [Google Scholar] [CrossRef]
- Lüneberg, E.; Mayer, B.; Daryab, N.; Kooistra, O.; Zähringer, U.; Rohde, M.; Swanson, J.; Frosch, M. Chromosomal insertion and excision of a 30 kb unstable genetic element is responsible for phase variation of lipopolysaccharide and other virulence determinants in Legionella pneumophila. Mol. Microbiol. 2001, 39, 1259–1271. [Google Scholar] [CrossRef]
- Kooistra, O.; Lüneberg, E.; Knirel, Y.A.; Frosch, M.; Zähringer, U. N-Methylation in polylegionaminic acid is associated with the phase-variable epitope of Legionella pneumophila serogroup 1 lipopolysaccharide. Identification of 5-(N,N-dimethylacetimidoyl) amino and 5-acetimidoyl(N-methyl)amino-7-acetamido-3,5,7,9-tetradeoxynon-2-ulosonic acid in the O-chain polysaccharide. Eur. J. Biochem. 2002, 269, 560–572. [Google Scholar] [CrossRef]
- Kooistra, O.; Knirel, Y.A.; Lüneberg, E.; Frosch, M.; Zähringer, U. Phase variation in Legionella pneumophila serogroup 1, subgroup OLDA, strain RC1 influences lipid A structure. In Proceedings of the 5th International Conference on Legionella, Ulm, Germany, 26–29 September 2000; Marre, R., Abu Kwaik, Y., Bartlett, C., Cianciotto, N.P., Fields, B.S., Frosch, M., Hacker, J., Lück, P.C., Eds.; ASM Press: Washington, DC, USA, 2002; pp. 68–73. [Google Scholar] [CrossRef]
- Borges, V.; Nunes, A.; Sampaio, D.A.; Vieira, L.; Machado, J.; Simões, M.J.; Gonçalves, P.; Gomes, J.P. Legionella pneumophila strain associated with the first evidence of person-to-person transmission of Legionnaires’ disease: A unique mosaic genetic backbone. Sci. Rep. 2016, 6, 26261. [Google Scholar] [CrossRef] [PubMed]
- Case, C.L.; Kohler, L.J.; Lima, J.B.; Strowig, T.; de Zoete, M.R.; Flavell, R.A.; Zamboni, D.S.; Roy, C.R. Caspase-11 stimulates rapid flagellin-independent pyroptosis in response to Legionella pneumophila. Proc. Natl. Acad. Sci. USA 2013, 110, 1851–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilla, D.M.; Hagar, J.A.; Haldar, A.K.; Mason, A.K.; Degrandi, D.; Pfeffer, K.; Ernst, R.K.; Yamamoto, M.; Miao, E.A.; Coers, J. Guanylate binding proteins promote caspase-11-dependent pyroptosis in response to cytoplasmic LPS. Proc. Natl. Acad. Sci. USA 2014, 111, 6046–6051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, K.; Ariki, S.; Kojima, T.; Saito, A.; Yamazoe, M.; Nishitani, C.; Shimizu, T.; Takahashi, M.; Mitsuzawa, H.; Yokota, S.; et al. Pulmonary collectins protect macrophages against pore-forming activity of Legionella pneumophila and suppress its intracellular growth. J. Biol. Chem. 2010, 285, 8434–8443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palusinska-Szysz, M.; Zdybicka-Barabas, A.; Cytryńska, M.; Wdowiak-Wróbel, S.; Chmiel, E.; Gruszecki, W.I. Analysis of cell surface alterations in Legionella pneumophila cells treated with human apolipoprotein E. Pathog. Dis. 2015, 73, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viasus, D.; Calatayud, L.; McBrown, M.V.; Ardanuy, C.; Carratalà, J. Urinary antigen testing in community-acquired pneumonia in adults: An update. Expert Rev. Anti-Infect. Ther. 2019, 17, 107–115. [Google Scholar] [CrossRef]
- Welte, T.; Torres, A.; Nathwani, D. Clinical and economic burden of community-acquired pneumonia among adults in Europe. Thorax 2012, 67, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Nisar, M.A.; Ross, K.E.; Brown, M.H.; Bentham, R.; Whiley, H. Legionella pneumophila and protozoan hosts: Implications for the control of hospital and potable water systems. Pathogens 2020, 9, 286. [Google Scholar] [CrossRef]
- Palazzolo, C.; Maffongelli, G.; D’Abramo, A.; Lepore, L.; Mariano, A.; Vulcano, A.; Bartoli, T.A.; Bevilacqua, N.; Giancola, M.L.; Di Rosa, E.; et al. Legionella pneumonia: Increased risk after COVID-19 lockdown? Italy, May to June 2020. Euro Surveill. 2020, 25, 1–3. [Google Scholar] [CrossRef]
- Gomez-Valero, L.; Rusniok, C.; Carson, D.; Mondino, S.; Pérez-Cobas, A.E.; Rolando, M.; Pasricha, S.; Reuter, S.; Demirtas, J.; Crumbach, J.; et al. More than 18,000 effectors in the Legionella genus genome provide multiple, independent combinations for replication in human cells. Proc. Natl. Acad. Sci. USA 2019, 5, 2265–2273. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, R.; Chowdhury, A.R.; Mukherjee, D.; Chakravortty, D. Lipid larceny: Channelizing host lipids for establishing successful pathogenesis by bacteria. Virulence 2021, 12, 195–216. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalczyk, B.; Chmiel, E.; Palusinska-Szysz, M. The Role of Lipids in Legionella-Host Interaction. Int. J. Mol. Sci. 2021, 22, 1487. https://doi.org/10.3390/ijms22031487
Kowalczyk B, Chmiel E, Palusinska-Szysz M. The Role of Lipids in Legionella-Host Interaction. International Journal of Molecular Sciences. 2021; 22(3):1487. https://doi.org/10.3390/ijms22031487
Chicago/Turabian StyleKowalczyk, Bozena, Elzbieta Chmiel, and Marta Palusinska-Szysz. 2021. "The Role of Lipids in Legionella-Host Interaction" International Journal of Molecular Sciences 22, no. 3: 1487. https://doi.org/10.3390/ijms22031487