
Diphlorethohydroxycarmalol Isolated from Ishige okamurae Exerts Vasodilatory Effects via Calcium Signaling and PI3K/Akt/eNOS Pathway



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Cytotoxicity of DPHC and Time-Dependent NO Production in EA.hy926 Cells
2.2. DPHC Promoted Phosphorylation in the PI3K/Akt/eNOS Pathway in EA.hy926 Cells
2.3. Regulatory Effects of DPHC on the [Ca2+]cytol and the [Ca2+]ER Levels
2.4. DPHC Modulated [Ca2+] Levels by Activating AchR and VEGFR2
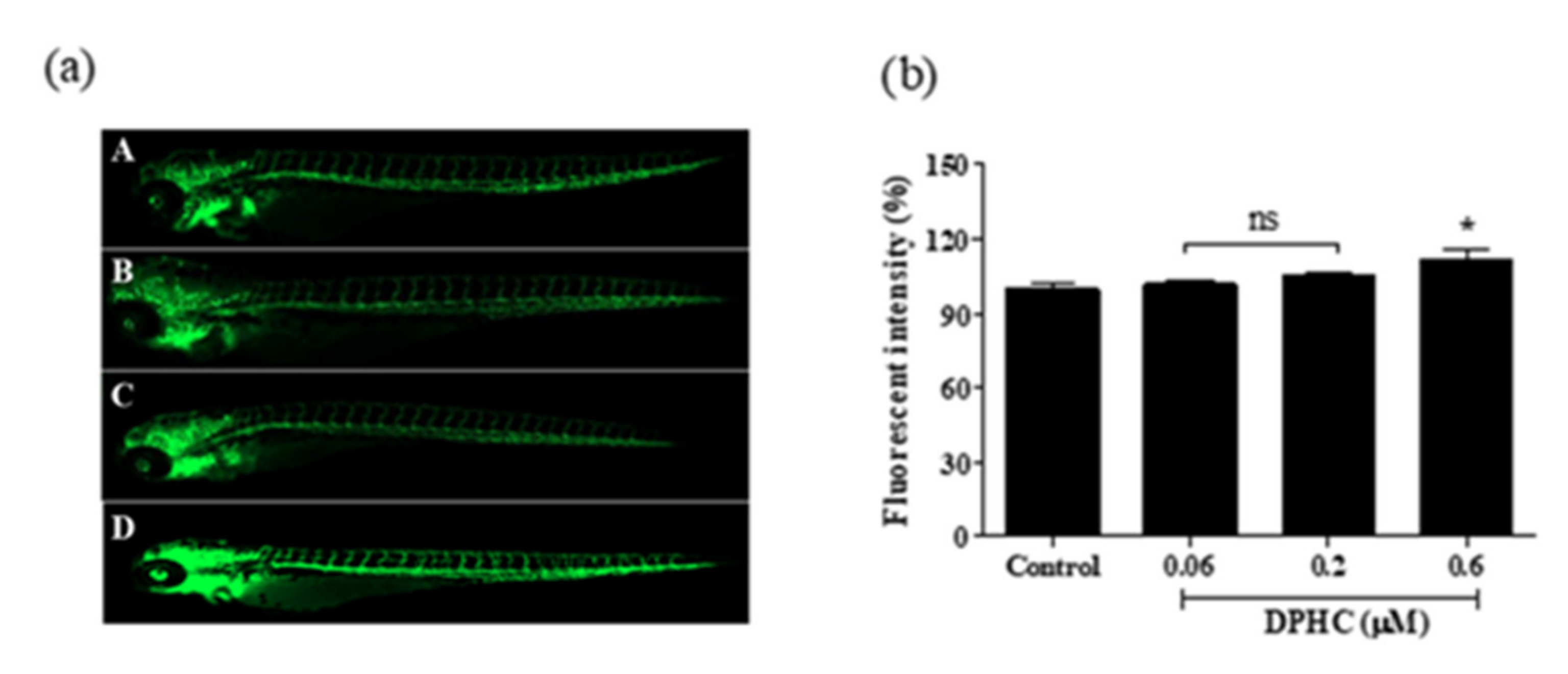
2.5. DPHC Enhanced Vasodilation in the Tg(flk:EGFP) Transgenic Zebrafish
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. DPHC Isolation
4.3. Cell Culture and Cell Viability Analysis
4.4. Quantification of the Intracellular NO Production
4.5. Quantification of the Intracellular and Extracellular Calcium Levels
4.6. Western Blot Analysis
4.7. Maintenance and Fluorescence Intensity Assessment in the Tg(flk:EGFP) Transgenic Zebrafish
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gómez-Guzmán, M.; Toral, M.; Romero, M.; Jiménez, R.; Galindo, P.; Sánchez, M.; Zarzuelo, M.J.; Olivares, M.; Gálvez, J.; Duarte, J. Antihypertensive effects of probiotics lactobacillus strains in spontaneously hypertensive rats. Mol. Nutr. Food Res. 2015, 59, 2326–2336. [Google Scholar] [CrossRef] [PubMed]
- Carrizzo, A.; Ambrosio, M.; Damato, A.; Madonna, M.; Storto, M.; Capocci, L.; Campiglia, P.; Sommella, E.; Trimarco, V.; Rozza, F.; et al. Morus alba extract modulates blood pressure homeostasis through enos signaling. Mol. Nutr. Food Res. 2016, 60, 2304–2311. [Google Scholar] [CrossRef] [PubMed]
- Robles-Vera, I.; Toral, M.; de la Visitación, N.; Sánchez, M.; Romero, M.; Olivares, M.; Jiménez, R.; Duarte, J. The probiotic lactobacillus fermentum prevents dysbiosis and vascular oxidative stress in rats with hypertension induced by chronic nitric oxide blockade. Mol. Nutr. Food Res. 2018, 62, 1800298. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Dai, Y.; Yan, S.; Shi, Y.; Li, J.; Liu, J.; Cha, L.; Mu, J. Resveratrol lowers blood pressure in spontaneously hypertensive rats via calcium-dependent endothelial no production. Clin. Exp. Hypertens. 2016, 38, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W.S. Vascular nitric oxide: Beyond enos. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef]
- Leung, W.K.; Gao, L.; Siu, P.M.; Lai, C.W. Diabetic nephropathy and endothelial dysfunction: Current and future therapies, and emerging of vascular imaging for preclinical renal-kinetic study. Life Sci. 2016, 166, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Nagpure, B.V.; Bian, J.S. Interaction of hydrogen sulfide with nitric oxide in the cardiovascular system. Oxid. Med. Cell Longev. 2016, 2016, 6904327. [Google Scholar] [CrossRef] [PubMed]
- Bernatova, I. Endothelial dysfunction in experimental models of arterial hypertension: Cause or consequence? Biomed. Res. Int. 2014, 2014, 598271. [Google Scholar] [CrossRef] [PubMed]
- Gheibi, S.; Jeddi, S.; Kashfi, K.; Ghasemi, A. Regulation of vascular tone homeostasis by no and h2s: Implications in hypertension. Biochem. Pharm. 2018, 149, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Fernando, I.P.S.; Ryu, B.; Ahn, G.; Yeo, I.-K.; Jeon, Y.-J. Therapeutic potential of algal natural products against metabolic syndrome: A review of recent developments. Trends Food Sci. Technol. 2020, 97, 286–299. [Google Scholar] [CrossRef]
- Lee, K.M.; Yang, E.C.; Coyer, J.A.; Zuccarello, G.C.; Wang, W.-L.; Choi, C.G.; Boo, S.M. Phylogeography of the seaweed ishige okamurae (phaeophyceae): Evidence for glacial refugia in the northwest pacific region. Mar. Biol. 2012, 159, 1021–1028. [Google Scholar] [CrossRef]
- Fernando, K.; Yang, H.-W.; Jiang, Y.; Jeon, Y.-J.; Ryu, B. Ishige okamurae extract and its constituent ishophloroglucin a attenuated in vitro and in vivo high glucose-induced angiogenesis. Int. J. Mol. Sci. 2019, 20, 5542. [Google Scholar] [CrossRef]
- Fernando, K.H.N.; Yang, H.W.; Jiang, Y.; Jeon, Y.J.; Ryu, B. Diphlorethohydroxycarmalol isolated from ishige okamurae represses high glucose-induced angiogenesis in vitro and in vivo. Mar. Drugs 2018, 16, 375. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, T.S.; de Oliveira, L.M.; de Oliveira, L.P.; Costa, R.M.D.; Tostes, R.C.; Georg, R.C.; Costa, E.A.; Lobato, N.S.; Filgueira, F.P.; Ghedini, P.C. Activation of PI3K/Akt pathway mediated by estrogen receptors accounts for estrone-induced vascular activation of cgmp signaling. Vasc. Pharm. 2018, 110, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Radu, B.M.; Osculati, A.M.M.; Suku, E.; Banciu, A.; Tsenov, G.; Merigo, F.; Di Chio, M.; Banciu, D.D.; Tognoli, C.; Kacer, P.; et al. All muscarinic acetylcholine receptors (M1-M5) are expressed in murine brain microvascular endothelium. Sci. Rep. 2017, 7, 5083. [Google Scholar] [CrossRef] [PubMed]
- McFadzean, I.; Gibson, A. The developing relationship between receptor-operated and store-operated calcium channels in smooth muscle. Br. J. Pharmacol. 2002, 135, 1–13. [Google Scholar] [CrossRef]
- Sun, G.; Liu, K. Developmental toxicity and cardiac effects of butyl benzyl phthalate in zebrafish embryos. Aquat. Toxicol. 2017, 192, 165–170. [Google Scholar] [CrossRef]
- Giardoglou, P.; Beis, D. On zebrafish disease models and matters of the heart. Biomedicines 2019, 7, 15. [Google Scholar] [CrossRef]
- Hügel, H.M.; Jackson, N.; May, B.; Zhang, A.L.; Xue, C.C. Polyphenol protection and treatment of hypertension. Phytomedicine 2016, 23, 220–231. [Google Scholar] [CrossRef]
- Khurana, S.; Venkataraman, K.; Hollingsworth, A.; Piche, M.; Tai, T.C. Polyphenols: Benefits to the cardiovascular system in health and in aging. Nutrients 2013, 5, 3779–3827. [Google Scholar] [CrossRef]
- Wijesinghe, W.; Ko, S.-C.; Jeon, Y.-J. Effect of phlorotannins isolated from ecklonia cava on angiotensin i-converting enzyme (ace) inhibitory activity. Nutr. Res. Pract. 2011, 5, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Guzmán, M.; Rodríguez-Nogales, A.; Algieri, F.; Gálvez, J. Potential role of seaweed polyphenols in cardiovascular-associated disorders. Mar. Drugs 2018, 16, 250. [Google Scholar] [CrossRef]
- Ko, S.-C.; Kang, M.C.; Kang, N.; Kim, H.-S.; Lee, S.-H.; Ahn, G.; Jung, W.-K.; Jeon, Y.-J. Effect of angiotensin i-converting enzyme (ace) inhibition and nitric oxide (no) production of 6,6′-bieckol, a marine algal polyphenol and its anti-hypertensive effect in spontaneously hypertensive rats. Process Biochem. 2017, 58, 326–332. [Google Scholar] [CrossRef]
- Kang, M.C.; Ding, Y.; Kim, H.S.; Jeon, Y.J.; Lee, S.H. Inhibition of adipogenesis by diphlorethohydroxycarmalol (dphc) through ampk activation in adipocytes. Mar. Drugs 2019, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- Dudzinski, D.M.; Michel, T. Life history of enos: Partners and pathways. Cardiovasc. Res. 2007, 75, 247–260. [Google Scholar] [CrossRef]
- Long, Y.; Xia, J.-Y.; Chen, S.-W.; Gao, C.-L.; Liang, G.-N.; He, X.-M.; Wu, J.; Jiang, C.-X.; Liu, X.; Huang, W.; et al. Atp2b1 gene silencing increases insulin sensitivity through facilitating akt activation via the ca(2+)/calmodulin signaling pathway and ca(2+)-associated enos activation in endothelial cells. Int. J. Biol. Sci. 2017, 13, 1203–1212. [Google Scholar] [CrossRef]
- Guizoni, D.M.; Vettorazzi, J.F.; Carneiro, E.M.; Davel, A.P. Modulation of endothelium-derived nitric oxide production and activity by taurine and taurine-conjugated bile acids. Nitric Oxide 2020, 94, 48–53. [Google Scholar] [CrossRef]
- Kida, T.; Tsubosaka, Y.; Hori, M.; Ozaki, H.; Murata, T. Bile acid receptor tgr5 agonism induces no production and reduces monocyte adhesion in vascular endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1663–1669. [Google Scholar] [CrossRef]
- Nakajima, T.; Okuda, Y.; Chisaki, K.; Shin, W.-S.; Iwasawa, K.; Morita, T.; Matsumoto, A.; Suzuki, J.-I.; Suzuki, S.; Yamada, N.; et al. Bile acids increase intracellular Ca2+ concentration and nitric oxide production in vascular endothelial cells. Br. J. Pharmacol. 2000, 130, 1457–1467. [Google Scholar] [CrossRef]
- Martinotti, S.; Patrone, M.; Balbo, V.; Mazzucco, L.; Ranzato, E. Endothelial response boosted by platelet lysate: The involvement of calcium toolkit. Int. J. Mol. Sci. 2020, 21, 808. [Google Scholar] [CrossRef] [PubMed]
- Bagur, R.; Hajnóczky, G. Intracellular Ca2+ sensing: Its role in calcium homeostasis and signaling. Mol. Cell 2017, 66, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Archundia, M.; Cordomi, A.; Garriga, P.; Perez, J.J. Molecular modeling of the m3 acetylcholine muscarinic receptor and its binding site. J. Biomed. Biotechnol. 2012, 2012, 789741. [Google Scholar] [CrossRef]
- Touyz, R.M.; Alves-Lopes, R.; Rios, F.J.; Camargo, L.L.; Anagnostopoulou, A.; Arner, A.; Montezano, A.C. Vascular smooth muscle contraction in hypertension. Cardiovasc. Res. 2018, 114, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.M.; Nakane, T.; Chiba, S. Muscarinic receptor subtypes mediating vasodilation and vasoconstriction in isolated, perfused simian coronary arteries. J. Cardiovasc. Pharmacol. 1993, 22, 841–846. [Google Scholar] [CrossRef]
- Brown, P.T.; Herbert, P.; Woodruff, R.I. Vitellogenesis in oncopeltus fasciatus: Plc/ip3, dag/pk-c pathway triggered by cam. J. Insect Physiol. 2010, 56, 1300–1305. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, R.; Zhong, H.; Tang, N.; Liu, Y.; Zhao, Y.; Zhang, T.; He, F. Casr participates in the regulation of vascular tension in the mesentery of hypertensive rats via the plc-ip3/ac-v/camp/ras pathway. Mol. Med. Rep. 2019, 20, 4433–4448. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Singhi, E.K.; Arroyo, J.P.; Ikizler, T.A.; Gould, E.R.; Brown, J.; Beckman, J.A.; Harrison, D.G.; Moslehi, J. Mechanisms of vegf (vascular endothelial growth factor) inhibitor–associated hypertension and vascular disease. Hypertension 2018, 71, e1–e8. [Google Scholar] [CrossRef]
- Tao, B.B.; Liu, S.Y.; Zhang, C.C.; Fu, W.; Cai, W.J.; Wang, Y.; Shen, Q.; Wang, M.J.; Chen, Y.; Zhang, L.J.; et al. Vegfr2 functions as an h2s-targeting receptor protein kinase with its novel cys1045-cys1024 disulfide bond serving as a specific molecular switch for hydrogen sulfide actions in vascular endothelial cells. Antioxid. Redox Signal. 2013, 19, 448–464. [Google Scholar] [CrossRef] [PubMed]
- Bencze, M. The role of calcium influx and calcium sensitization in contraction of isolated arteries of normotensive and hypertensive rat. 2017. Available online: https://dspace.cuni.cz/handle/20.500.11956/96364 (accessed on 5 February 2021).
- Vimalraj, S.; Pichu, S.; Pankajam, T.; Dharanibalan, K.; Djonov, V.; Chatterjee, S. Nitric oxide regulates intussusceptive-like angiogenesis in wound repair in chicken embryo and transgenic zebrafish models. Nitric Oxide 2019, 82, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Margiotta-Casaluci, L.; Owen, S.F.; Rand-Weaver, M.; Winter, M.J. Testing the translational power of the zebrafish: An interspecies analysis of responses to cardiovascular drugs. Front. Pharm. 2019, 10, 893. [Google Scholar] [CrossRef] [PubMed]
- Wiggenhauser, L.M.; Kroll, J. Vascular damage in obesity and diabetes: Highlighting links between endothelial dysfunction and metabolic disease in zebrafish and man. Curr. Vasc. Pharmacol. 2019, 17, 476–490. [Google Scholar] [CrossRef]
- Fritsche, R.; Schwerte, T.; Pelster, B. Nitric oxide and vascular reactivity in developing zebrafish, danio rerio. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R2200–R2207. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, N.; Childs, S.J. Development of vascular regulation in the zebrafish embryo. Development 2020, 147, dev183061. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Fresta, C.G.; Siegel, J.M.; Wijesinghe, M.B.; Lunte, S.M. Microchip electrophoresis with laser-induced fluorescence detection for the determination of the ratio of nitric oxide to superoxide production in macrophages during inflammation. Anal. Bioanal. Chem. 2017, 409, 4529–4538. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Y.A.; Jiang, Y.; Yang, H.-W.; Hwang, J.; Jeon, Y.-J.; Ryu, B. Diphlorethohydroxycarmalol Isolated from Ishige okamurae Exerts Vasodilatory Effects via Calcium Signaling and PI3K/Akt/eNOS Pathway. Int. J. Mol. Sci. 2021, 22, 1610. https://doi.org/10.3390/ijms22041610
Lu YA, Jiang Y, Yang H-W, Hwang J, Jeon Y-J, Ryu B. Diphlorethohydroxycarmalol Isolated from Ishige okamurae Exerts Vasodilatory Effects via Calcium Signaling and PI3K/Akt/eNOS Pathway. International Journal of Molecular Sciences. 2021; 22(4):1610. https://doi.org/10.3390/ijms22041610
Chicago/Turabian StyleLu, Yu An, Yunfei Jiang, Hye-Won Yang, Jin Hwang, You-Jin Jeon, and Bomi Ryu. 2021. "Diphlorethohydroxycarmalol Isolated from Ishige okamurae Exerts Vasodilatory Effects via Calcium Signaling and PI3K/Akt/eNOS Pathway" International Journal of Molecular Sciences 22, no. 4: 1610. https://doi.org/10.3390/ijms22041610
APA StyleLu, Y. A., Jiang, Y., Yang, H.-W., Hwang, J., Jeon, Y.-J., & Ryu, B. (2021). Diphlorethohydroxycarmalol Isolated from Ishige okamurae Exerts Vasodilatory Effects via Calcium Signaling and PI3K/Akt/eNOS Pathway. International Journal of Molecular Sciences, 22(4), 1610. https://doi.org/10.3390/ijms22041610