
Genetic Bias, Diversity Indices, Physiochemical Properties and CDR3 Motifs Divide Auto-Reactive from Allo-Reactive T-Cell Repertoires


 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Antigen-Specific T-Cells Show Different Frequencies between Auto-Reactive and Allo-Reactive Settings
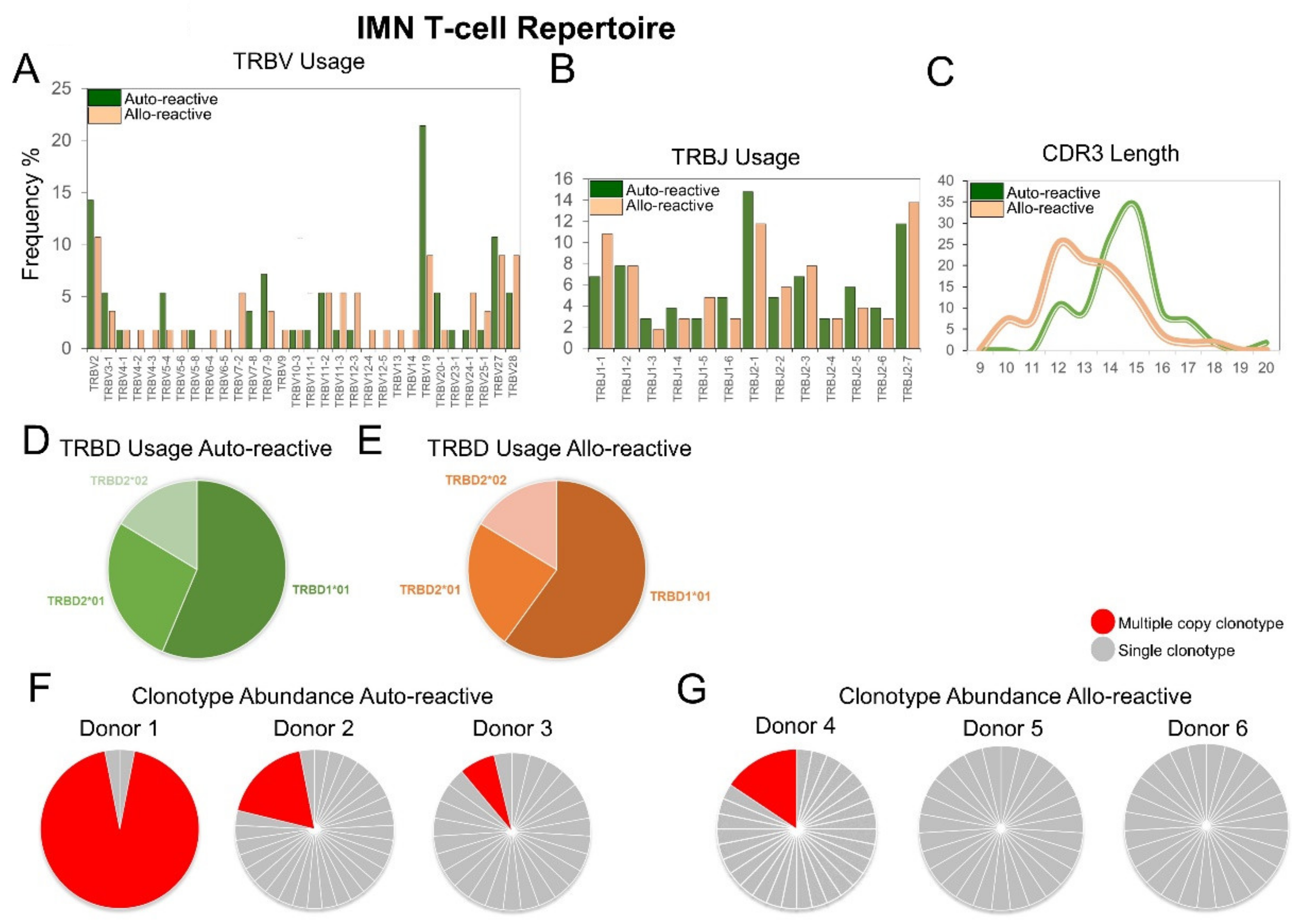
2.2. The IMN Allo-Reactive T-Cell Repertoire Exhibits Expanded TRBV Gene Usage and Unconventional CDR3 Loop Length
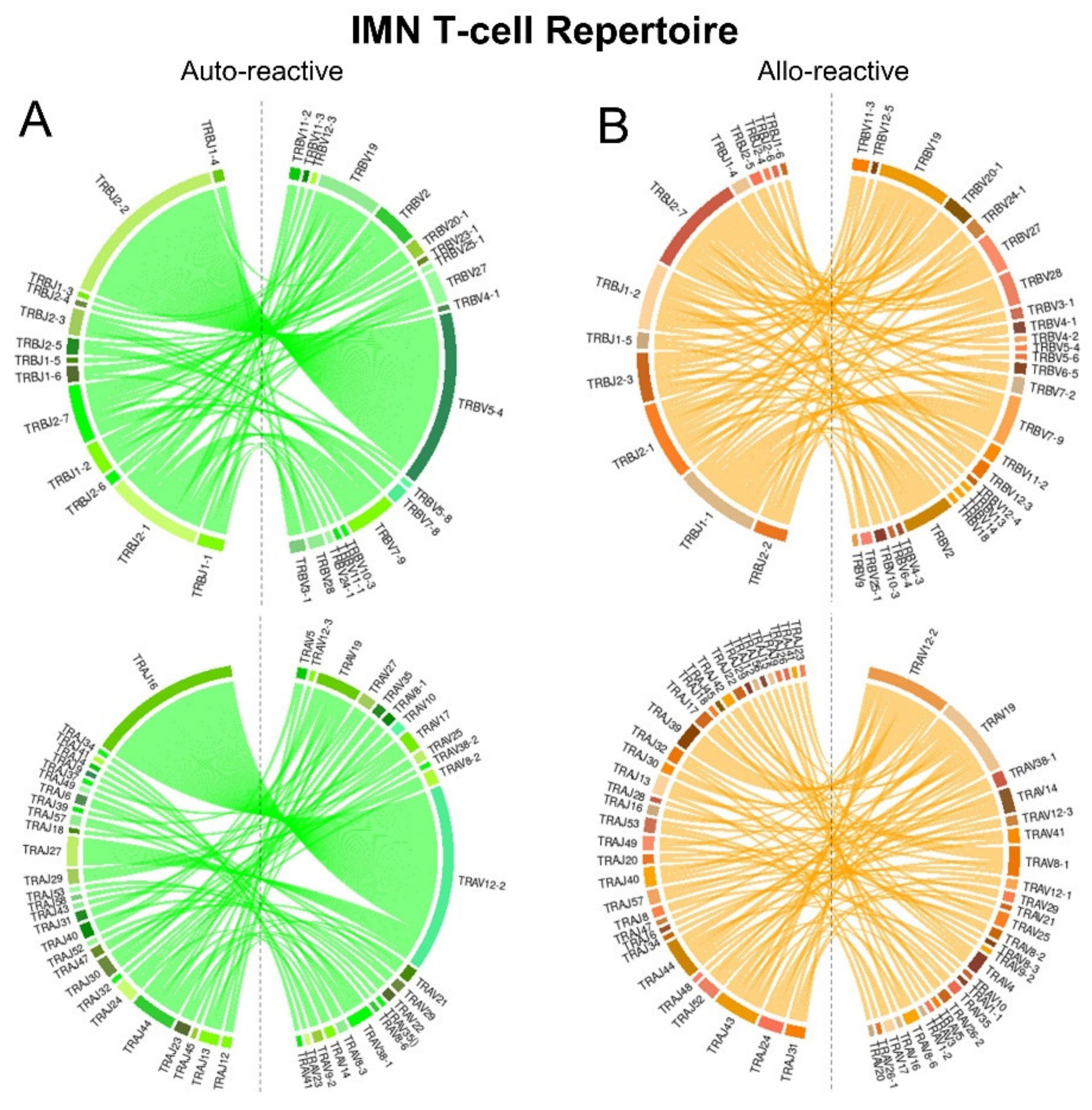
2.3. IMN-Specific T-Cell Repertoires Exhibit Divergent TCR Gene Pairing
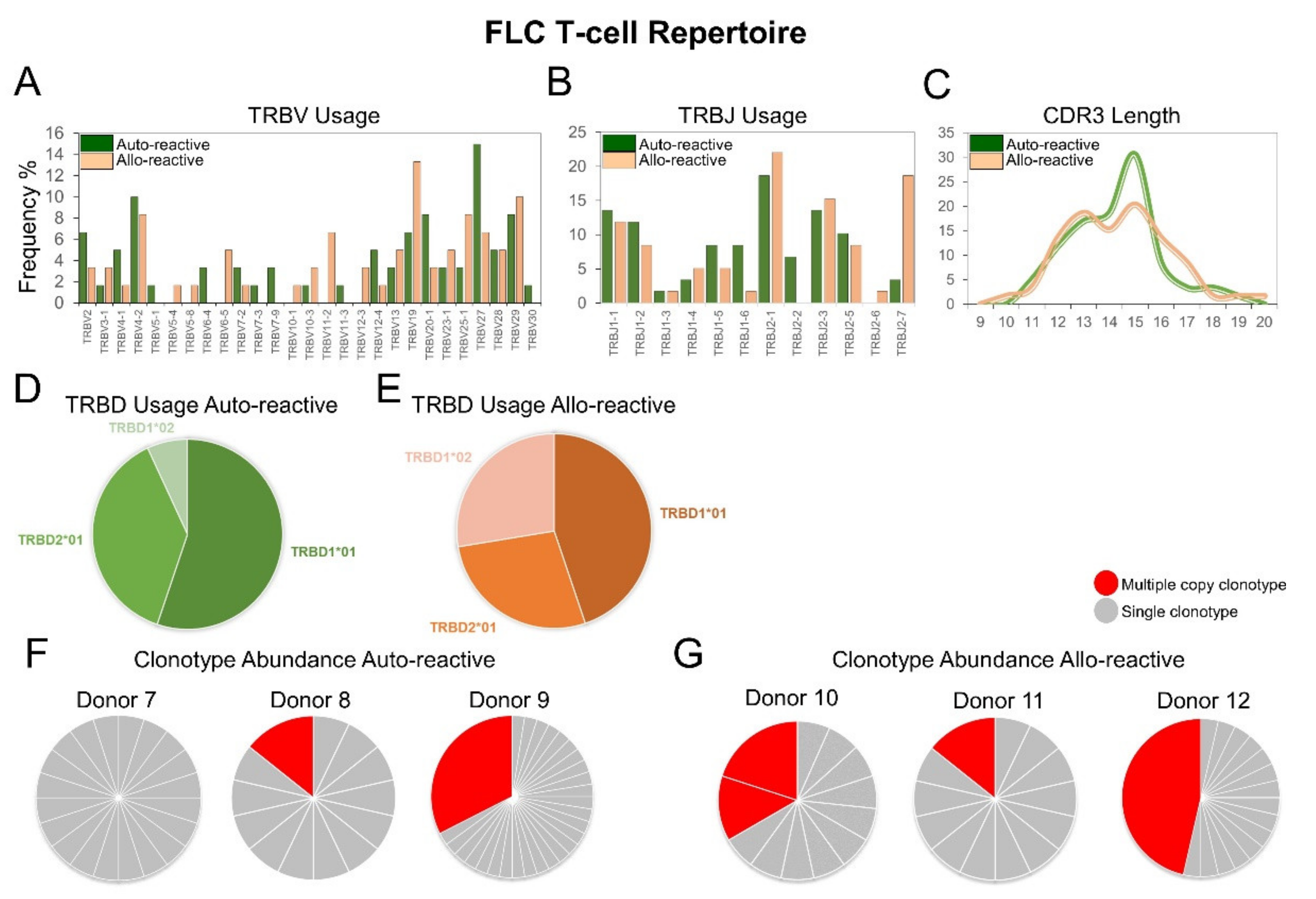
2.4. The FLC-Specific Allo-Reactive T-Cell Repertoire Exhibits Divergent TRBV and TRBJ Gene Usage, Unconventional CDR3 Loop Length and Common Multiple Copy Clonotypes
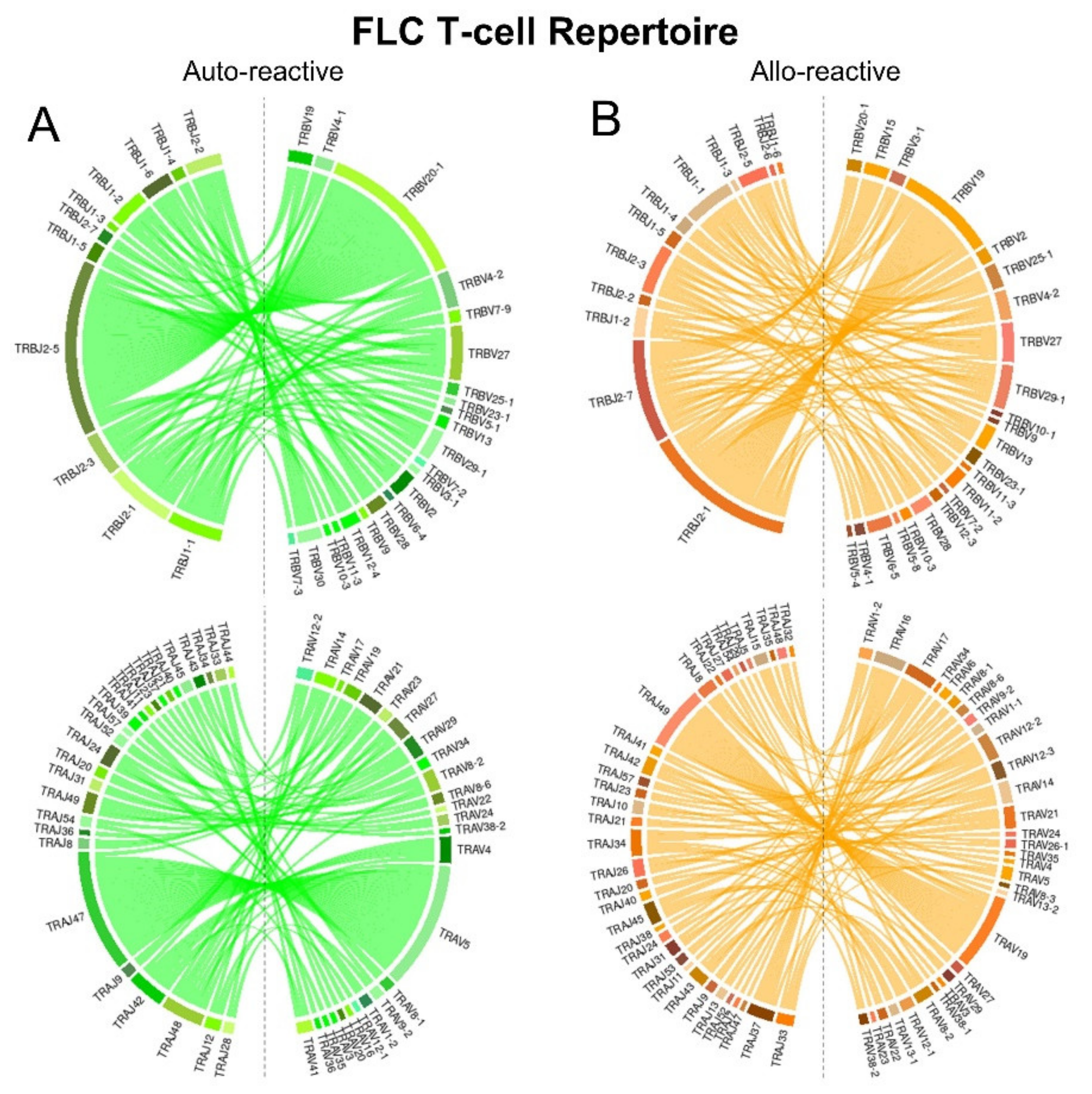
2.5. FLC-Specific T-Cell Repertoires Exhibit Divergent TCR Gene Pairing
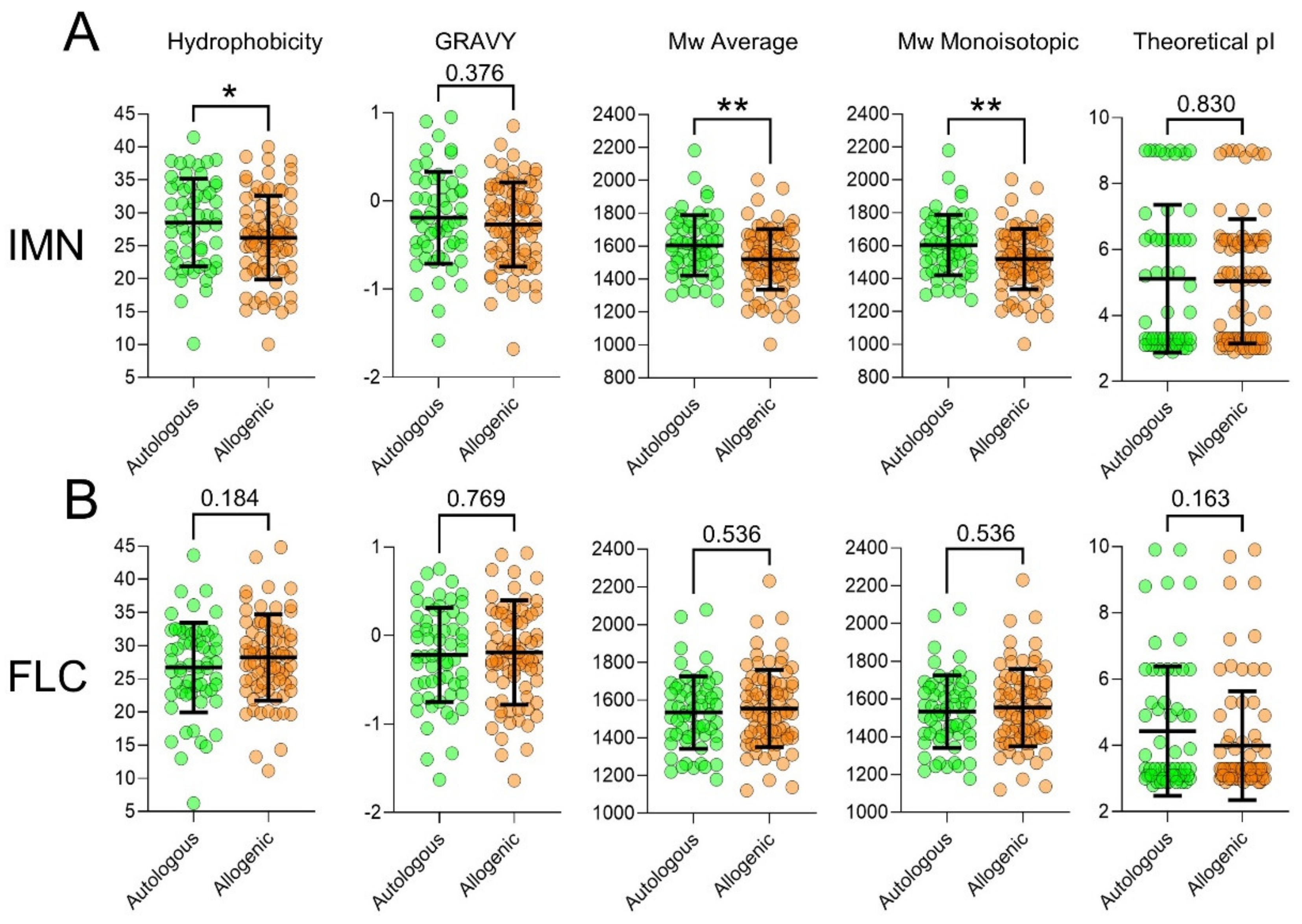
2.6. T-Cells Exhibit Divergent CDR3 Physical-Chemical Properties between Auto-Reactive and Allo-Reactive Repertoires
2.7. T-Cells Exhibit Divergent CDR3 Diversity Metrics between Auto-Reactive and Allo-Reactive Repertoires
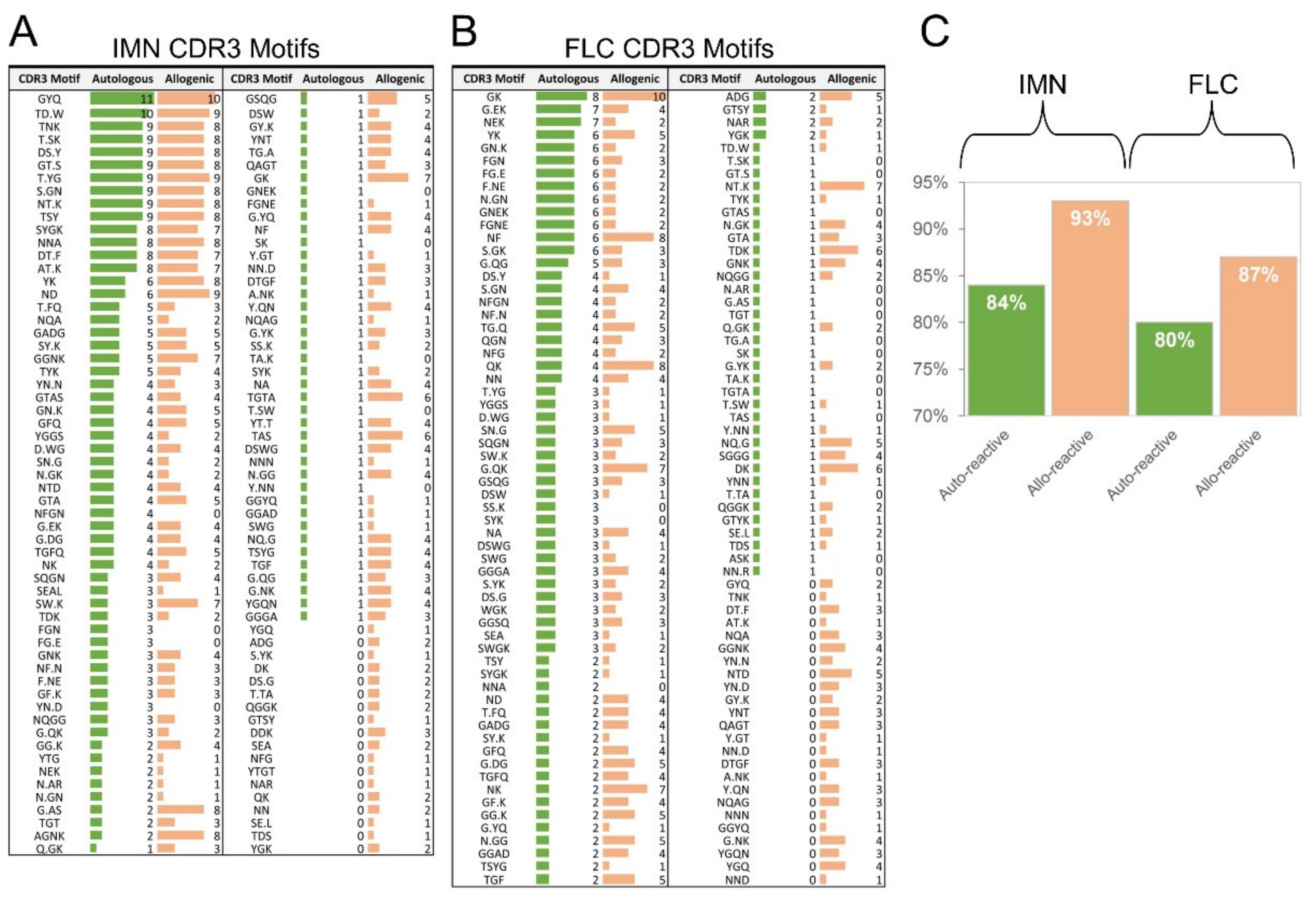
2.8. T-Cells Exhibit Divergent CDR3 Motifs between Auto-Reactive and Allo-Reactive Repertoires
3. Discussion
4. Materials and Methods
4.1. Sample Preparation and Processing
4.2. Flow Cytometry and Single-Cell Sorting
4.3. Single-Cell TCRαβ Amplification and Annotation
4.4. Diversity Analysis

4.5. Physical-Chemical Properties
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TCR | T-cell receptor |
| MHC | Major histocompatibility complex |
| HLA | Human leukocyte antigen complex |
| p | Peptide within the MHC/HLA binding grove |
| TRBV | T-cell receptor β gene |
| TRBJ | T-cell receptor β joining gene |
| TRBD | T-cell receptor β diversity gene |
| TRAV | T-cell receptor α region gene |
| TRAJ | T-cell receptor α joining gene |
| CDR | Complementarity determining region |
References
- Felix, N.J.; Allen, P.M. Specificity of T-cell alloreactivity. Nat. Rev. Immunol. 2007, 7, 942–953. [Google Scholar] [CrossRef]
- Gkazi, A.S.; Margetts, B.K.; Attenborough, T.; Mhaldien, L.; Standing, J.F.; Oakes, T.; Heather, J.M.; Booth, J.; Pasquet, M.; Chiesa, R.; et al. Clinical T Cell Receptor Repertoire Deep Sequencing and Analysis: An Application to Monitor Immune Reconstitution Following Cord Blood Transplantation. Front. Immunol. 2018, 9, 2547. [Google Scholar] [CrossRef]
- Klarenbeek, P.L.; Remmerswaal, E.B.; ten Berge, I.J.; Doorenspleet, M.E.; van Schaik, B.D.; Esveldt, R.E.; Koch, S.D.; ten Brinke, A.; van Kampen, A.H.; Bemelman, F.J.; et al. Deep sequencing of antiviral T-cell responses to HCMV and EBV in humans reveals a stable repertoire that is maintained for many years. PLoS Pathog. 2012, 8, e1002889. [Google Scholar] [CrossRef] [PubMed]
- van Heijst, J.W.; Ceberio, I.; Lipuma, L.B.; Samilo, D.W.; Wasilewski, G.D.; Gonzales, A.M.; Nieves, J.L.; van den Brink, M.R.; Perales, M.A.; Pamer, E.G. Quantitative assessment of T cell repertoire recovery after hematopoietic stem cell transplantation. Nat. Med. 2013, 19, 372–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.L.; McIntosh, C.M.; Williams, J.B.; Wang, Y.; Hollinger, M.K.; Isaad, N.J.; Moon, J.J.; Gajewski, T.F.; Chong, A.S.; Alegre, M.L. Distinct Graft-Specific TCR Avidity Profiles during Acute Rejection and Tolerance. Cell Rep. 2018, 24, 2112–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolich-Zugich, J. High specificity, not degeneracy, allows T cell alloresponses. Nat. Immunol. 2007, 8, 335–337. [Google Scholar] [CrossRef]
- Archbold, J.K.; Macdonald, W.A.; Miles, J.J.; Brennan, R.M.; Kjer-Nielsen, L.; McCluskey, J.; Burrows, S.R.; Rossjohn, J. Alloreactivity between disparate cognate and allogeneic pMHC-I complexes is the result of highly focused, peptide-dependent structural mimicry. J. Biol. Chem. 2006, 281, 34324–34332. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.C.; Miles, J.J.; Neller, M.A.; Gostick, E.; Price, D.A.; Purcell, A.W.; McCluskey, J.; Burrows, S.R.; Rossjohn, J.; Gras, S. Highly divergent T-cell receptor binding modes underlie specific recognition of a bulged viral peptide bound to a human leukocyte antigen class I molecule. J. Biol. Chem. 2013, 288, 15442–15454. [Google Scholar] [CrossRef] [Green Version]
- Rossjohn, J.; Gras, S.; Miles, J.J.; Turner, S.J.; Godfrey, D.I.; McCluskey, J. T cell antigen receptor recognition of antigen-presenting molecules. Annu. Rev. Immunol. 2015, 33, 169–200. [Google Scholar] [CrossRef]
- Miles, J.J.; McCluskey, J.; Rossjohn, J.; Gras, S. Understanding the complexity and malleability of T-cell recognition. Immunol. Cell Biol. 2015, 93, 433–441. [Google Scholar] [CrossRef]
- Chapman, J.R.; Webster, A.C.; Wong, G. Cancer in the transplant recipient. Cold Spring Harb. Perspect. Med. 2013, 3. [Google Scholar] [CrossRef] [Green Version]
- Benichou, G.; Gonzalez, B.; Marino, J.; Ayasoufi, K.; Valujskikh, A. Role of Memory T Cells in Allograft Rejection and Tolerance. Front. Immunol. 2017, 8, 170. [Google Scholar] [CrossRef] [Green Version]
- Ingulli, E. Mechanism of cellular rejection in transplantation. Pediatr. Nephrol. 2010, 25, 61–74. [Google Scholar] [CrossRef] [Green Version]
- DeWolf, S.; Sykes, M. Alloimmune T cells in transplantation. J. Clin. Investig. 2017, 127, 2473–2481. [Google Scholar] [CrossRef] [Green Version]
- Watkins, T.S.; Miles, J.J. The human T cell receptor repertoire in health and disease and potential for omics integration. Immunol. Cell Biol. 2020. [Google Scholar] [CrossRef]
- Kjer-Nielsen, L.; Clements, C.S.; Purcell, A.W.; Brooks, A.G.; Whisstock, J.C.; Burrows, S.R.; McCluskey, J.; Rossjohn, J. A structural basis for the selection of dominant alphabeta T cell receptors in antiviral immunity. Immunity 2003, 18, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, W.A.; Chen, Z.; Gras, S.; Archbold, J.K.; Tynan, F.E.; Clements, C.S.; Bharadwaj, M.; Kjer-Nielsen, L.; Saunders, P.M.; Wilce, M.C.; et al. T cell allorecognition via molecular mimicry. Immunity 2009, 31, 897–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gras, S.; Burrows, S.R.; Kjer-Nielsen, L.; Clements, C.S.; Liu, Y.C.; Sullivan, L.C.; Bell, M.J.; Brooks, A.G.; Purcell, A.W.; McCluskey, J.; et al. The shaping of T cell receptor recognition by self-tolerance. Immunity 2009, 30, 193–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speir, J.A.; Garcia, K.C.; Brunmark, A.; Degano, M.; Peterson, P.A.; Teyton, L.; Wilson, I.A. Structural basis of 2C TCR allorecognition of H-2Ld peptide complexes. Immunity 1998, 8, 553–562. [Google Scholar] [CrossRef] [Green Version]
- Reiser, J.B.; Darnault, C.; Guimezanes, A.; Gregoire, C.; Mosser, T.; Schmitt-Verhulst, A.M.; Fontecilla-Camps, J.C.; Malissen, B.; Housset, D.; Mazza, G. Crystal structure of a T cell receptor bound to an allogeneic MHC molecule. Nat. Immunol. 2000, 1, 291–297. [Google Scholar] [CrossRef]
- Colf, L.A.; Bankovich, A.J.; Hanick, N.A.; Bowerman, N.A.; Jones, L.L.; Kranz, D.M.; Garcia, K.C. How a single T cell receptor recognizes both self and foreign MHC. Cell 2007, 129, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.L.; Colf, L.A.; Stone, J.D.; Garcia, K.C.; Kranz, D.M. Distinct CDR3 conformations in TCRs determine the level of cross-reactivity for diverse antigens, but not the docking orientation. J. Immunol. 2008, 181, 6255–6264. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Singh, N.K.; Spear, T.T.; Hellman, L.M.; Piepenbrink, K.H.; McMahan, R.H.; Rosen, H.R.; Vander Kooi, C.W.; Nishimura, M.I.; Baker, B.M. How an alloreactive T-cell receptor achieves peptide and MHC specificity. Proc. Natl. Acad. Sci. USA 2017, 114, E4792–E4801. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Huseby, E.; Scott-Browne, J.; Rubtsova, K.; Pinilla, C.; Crawford, F.; Marrack, P.; Dai, S.; Kappler, J.W. A Single T Cell Receptor Bound to Major Histocompatibility Complex Class I and Class II Glycoproteins Reveals Switchable TCR Conformers. Immunity 2011, 35, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rist, M.; Smith, C.; Bell, M.J.; Burrows, S.R.; Khanna, R. Cross-recognition of HLA DR4 alloantigen by virus-specific CD8+ T cells: A new paradigm for self-/nonself-recognition. Blood 2009, 114, 2244–2253. [Google Scholar] [CrossRef] [PubMed]
- Rist, M.J.; Theodossis, A.; Croft, N.P.; Neller, M.A.; Welland, A.; Chen, Z.; Sullivan, L.C.; Burrows, J.M.; Miles, J.J.; Brennan, R.M.; et al. HLA peptide length preferences control CD8+ T cell responses. J. Immunol. 2013, 191, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Rist, M.J.; Hibbert, K.M.; Croft, N.P.; Smith, C.; Neller, M.A.; Burrows, J.M.; Miles, J.J.; Purcell, A.W.; Rossjohn, J.; Gras, S.; et al. T Cell Cross-Reactivity between a Highly Immunogenic EBV Epitope and a Self-Peptide Naturally Presented by HLA-B*18:01+ Cells. J Immunol 2015, 194, 4668–4675. [Google Scholar] [CrossRef] [Green Version]
- Cole, D.K.; Miles, K.M.; Madura, F.; Holland, C.J.; Schauenburg, A.J.; Godkin, A.J.; Bulek, A.M.; Fuller, A.; Akpovwa, H.J.; Pymm, P.G.; et al. T-cell receptor (TCR)-peptide specificity overrides affinity-enhancing TCR-major histocompatibility complex interactions. J. Biol. Chem. 2014, 289, 628–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, P.M.; Faure, O.; Graff-Dubois, S.; Gross, D.A.; Cornet, S.; Chouaib, S.; Miconnet, I.; Lemonnier, F.A.; Kosmatopoulos, K. EphA2 as target of anticancer immunotherapy: Identification of HLA-A*0201-restricted epitopes. Cancer Res. 2003, 63, 8476–8480. [Google Scholar]
- Marrari, A.; Iero, M.; Pilla, L.; Villa, S.; Salvioni, R.; Valdagni, R.; Parmiani, G.; Rivoltini, L. Vaccination therapy in prostate cancer. Cancer Immunol. Immunother. 2007, 56, 429–445. [Google Scholar] [CrossRef]
- Novellino, L.; Castelli, C.; Parmiani, G. A listing of human tumor antigens recognized by T cells: March 2004 update. Cancer Immunol. Immunother. 2005, 54, 187–207. [Google Scholar] [CrossRef]
- McNeel, D.G.; Olsen, B.M. Prostate Cancer Vaccine. U.S. Patent 9,433,668,B2, 2016. [Google Scholar]
- Wang, L.G.; Johnson, E.M.; Kinoshita, Y.; Babb, J.S.; Buckley, M.T.; Liebes, L.F.; Melamed, J.; Liu, X.M.; Kurek, R.; Ossowski, L.; et al. Androgen receptor overexpression in prostate cancer linked to Pur alpha loss from a novel repressor complex. Cancer Res. 2008, 68, 2678–2688. [Google Scholar] [CrossRef] [Green Version]
- Giovannelli, P.; Di Donato, M.; Galasso, G.; Di Zazzo, E.; Bilancio, A.; Migliaccio, A. The Androgen Receptor in Breast Cancer. Front. Endocrinol. (Lausanne) 2018, 9, 492. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Li, X.X.; Yang, Y.; Zhang, Y.; Wang, H.Y.; Zheng, X.F.S. Significance and mechanism of androgen receptor overexpression and androgen receptor/mechanistic target of rapamycin cross-talk in hepatocellular carcinoma. Hepatology 2018, 67, 2271–2286. [Google Scholar] [CrossRef]
- Weon, J.L.; Potts, P.R. The MAGE protein family and cancer. Curr. Opin. Cell Biol. 2015, 37, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, Y.; Zhang, L. Expression of cancer-testis antigens in esophageal cancer and their progress in immunotherapy. J. Cancer Res. Clin. Oncol. 2019, 145, 281–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fossa, A.; Alsoe, L.; Crameri, R.; Funderud, S.; Gaudernack, G.; Smeland, E.B. Serological cloning of cancer/testis antigens expressed in prostate cancer using cDNA phage surface display. Cancer Immunol. Immunother. 2004, 53, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Neller, M.A.; Ladell, K.; McLaren, J.E.; Matthews, K.K.; Gostick, E.; Pentier, J.M.; Dolton, G.; Schauenburg, A.J.; Koning, D.; Fontaine Costa, A.I.; et al. Naive CD8(+) T-cell precursors display structured TCR repertoires and composite antigen-driven selection dynamics. Immunol. Cell Biol. 2015, 93, 625–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolton, G.; Tungatt, K.; Lloyd, A.; Bianchi, V.; Theaker, S.M.; Trimby, A.; Holland, C.J.; Donia, M.; Godkin, A.J.; Cole, D.K.; et al. More tricks with tetramers: A practical guide to staining T cells with peptide-MHC multimers. Immunology 2015, 146, 11–22. [Google Scholar] [CrossRef]
- Dolton, G.; Lissina, A.; Skowera, A.; Ladell, K.; Tungatt, K.; Jones, E.; Kronenberg-Versteeg, D.; Akpovwa, H.; Pentier, J.M.; Holland, C.J.; et al. Comparison of peptide-major histocompatibility complex tetramers and dextramers for the identification of antigen-specific T cells. Clin. Exp. Immunol. 2014, 177, 47–63. [Google Scholar] [CrossRef]
- Lissina, A.; Ladell, K.; Skowera, A.; Clement, M.; Edwards, E.; Seggewiss, R.; van den Berg, H.A.; Gostick, E.; Gallagher, K.; Jones, E.; et al. Protein kinase inhibitors substantially improve the physical detection of T-cells with peptide-MHC tetramers. J. Immunol. Methods 2009, 340, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glanville, J.; Huang, H.; Nau, A.; Hatton, O.; Wagar, L.E.; Rubelt, F.; Ji, X.; Han, A.; Krams, S.M.; Pettus, C.; et al. Identifying specificity groups in the T cell receptor repertoire. Nature 2017, 547, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Cole, D.K.; Yuan, F.; Rizkallah, P.J.; Miles, J.J.; Gostick, E.; Price, D.A.; Gao, G.F.; Jakobsen, B.K.; Sewell, A.K. Germ line-governed recognition of a cancer epitope by an immunodominant human T-cell receptor. J. Biol. Chem. 2009, 284, 27281–27289. [Google Scholar] [CrossRef] [Green Version]
- Madura, F.; Rizkallah, P.J.; Legut, M.; Holland, C.J.; Fuller, A.; Bulek, A.; Schauenburg, A.J.; Trimby, A.; Hopkins, J.R.; Wells, S.A.; et al. TCR-induced alteration of primary MHC peptide anchor residue. Eur. J. Immunol. 2019, 49, 1052–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, J.J.; Douek, D.C.; Price, D.A. Bias in the alphabeta T-cell repertoire: Implications for disease pathogenesis and vaccination. Immunol. Cell Biol. 2011, 89, 375–387. [Google Scholar] [CrossRef]
- Venturi, V.; Price, D.A.; Douek, D.C.; Davenport, M.P. The molecular basis for public T-cell responses? Nat. Rev. Immunol. 2008, 8, 231–238. [Google Scholar] [CrossRef]
- Melenhorst, J.J.; Lay, M.D.; Price, D.A.; Adams, S.D.; Zeilah, J.; Sosa, E.; Hensel, N.F.; Follmann, D.; Douek, D.C.; Davenport, M.P.; et al. Contribution of TCR-beta locus and HLA to the shape of the mature human Vbeta repertoire. J. Immunol. 2008, 180, 6484–6489. [Google Scholar] [CrossRef] [Green Version]
- Min, B. Spontaneous T Cell Proliferation: A Physiologic Process to Create and Maintain Homeostatic Balance and Diversity of the Immune System. Front. Immunol. 2018, 9, 547. [Google Scholar] [CrossRef]
- Stadinski, B.D.; Shekhar, K.; Gomez-Tourino, I.; Jung, J.; Sasaki, K.; Sewell, A.K.; Peakman, M.; Chakraborty, A.K.; Huseby, E.S. Hydrophobic CDR3 residues promote the development of self-reactive T cells. Nat. Immunol. 2016, 17, 946–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, L.A.; Volpi, S.; Frugoni, F.; Janssen, E.; Kim, S.; Sundel, R.P.; Dedeoglu, F.; Lo, M.S.; Hazen, M.M.; Beth Son, M.; et al. Next-Generation Sequencing Reveals Restriction and Clonotypic Expansion of Treg Cells in Juvenile Idiopathic Arthritis. Arthritis Rheumatol. 2016, 68, 1758–1768. [Google Scholar] [CrossRef]
- Rosenthal, K.M.; Edwards, L.J.; Sabatino, J.J., Jr.; Hood, J.D.; Wasserman, H.A.; Zhu, C.; Evavold, B.D. Low 2-dimensional CD4 T cell receptor affinity for myelin sets in motion delayed response kinetics. PLoS ONE 2012, 7, e32562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, J.J.; Bulek, A.M.; Cole, D.K.; Gostick, E.; Schauenburg, A.J.; Dolton, G.; Venturi, V.; Davenport, M.P.; Tan, M.P.; Burrows, S.R.; et al. Genetic and structural basis for selection of a ubiquitous T cell receptor deployed in Epstein-Barr virus infection. PLoS Pathog. 2010, 6, e1001198. [Google Scholar] [CrossRef] [PubMed]
- Ely, L.K.; Green, K.J.; Beddoe, T.; Clements, C.S.; Miles, J.J.; Bottomley, S.P.; Zernich, D.; Kjer-Nielsen, L.; Purcell, A.W.; McCluskey, J.; et al. Antagonism of antiviral and allogeneic activity of a human public CTL clonotype by a single altered peptide ligand: Implications for allograft rejection. J. Immunol. 2005, 174, 5593–5601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, J.J.; Silins, S.L.; Burrows, S.R. Engineered T cell receptors and their potential in molecular medicine. Curr. Med. Chem. 2006, 13, 2725–2736. [Google Scholar] [CrossRef]
- Boudousquie, C.; Bossi, G.; Hurst, J.M.; Rygiel, K.A.; Jakobsen, B.K.; Hassan, N.J. Polyfunctional response by ImmTAC (IMCgp100) redirected CD8(+) and CD4(+) T cells. Immunology 2017, 152, 425–438. [Google Scholar] [CrossRef] [Green Version]
- Sant, S.; Grzelak, L.; Wang, Z.; Pizzolla, A.; Koutsakos, M.; Crowe, J.; Loudovaris, T.; Mannering, S.I.; Westall, G.P.; Wakim, L.M.; et al. Single-Cell Approach to Influenza-Specific CD8(+) T Cell Receptor Repertoires Across Different Age Groups, Tissues, and Following Influenza Virus Infection. Front. Immunol. 2018, 9, 1453. [Google Scholar] [CrossRef] [Green Version]
- Van de Sandt, C.E.; Clemens, E.B.; Grant, E.J.; Rowntree, L.C.; Sant, S.; Halim, H.; Crowe, J.; Cheng, A.C.; Kotsimbos, T.C.; Richards, M.; et al. Challenging immunodominance of influenza-specific CD8(+) T cell responses restricted by the risk-associated HLA-A*68:01 allomorph. Nat. Commun. 2019, 10, 5579. [Google Scholar] [CrossRef]
- Kedzierska, K.; Venturi, V.; Field, K.; Davenport, M.P.; Turner, S.J.; Doherty, P.C. Early establishment of diverse T cell receptor profiles for influenza-specific CD8(+)CD62L(hi) memory T cells. Proc. Natl. Acad. Sci. U.S.A. 2006, 103, 9184–9189. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.S.; Li, L.; Zhang, Y.P. Defining Individual-Level Genetic Diversity and Similarity Profiles. Sci. Rep. 2020, 10, 5805. [Google Scholar] [CrossRef] [Green Version]
- Trowsdale, J. The MHC, disease and selection. Immunol. Lett. 2011, 137, 1–8. [Google Scholar] [CrossRef]
- Lepletier, A.; Lutzky, V.P.; Mittal, D.; Stannard, K.; Watkins, T.S.; Ratnatunga, C.N.; Smith, C.; McGuire, H.M.; Kemp, R.A.; Mukhopadhyay, P.; et al. The immune checkpoint CD96 defines a distinct lymphocyte phenotype and is highly expressed on tumor-infiltrating T cells. Immunol. Cell Biol. 2019, 97, 152–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.C.; Dash, P.; McCullers, J.A.; Doherty, P.C.; Thomas, P.G. T cell receptor alphabeta diversity inversely correlates with pathogen-specific antibody levels in human cytomegalovirus infection. Sci. Transl. Med. 2012, 4, 128ra142. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Lefranc, M.P.; Miles, J.J.; Alamyar, E.; Giudicelli, V.; Duroux, P.; Freeman, J.D.; Corbin, V.D.; Scheerlinck, J.P.; Frohman, M.A.; et al. IMGT/HighV QUEST paradigm for T cell receptor IMGT clonotype diversity and next generation repertoire immunoprofiling. Nat. Commun. 2013, 4, 2333. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. Circlize Implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A: IMN T-cell repertoire | ||
| Diversity Metric | p Value | Log2FC |
| Biodiversity | 0.38 | −0.15 |
| Simpson’s index | 0.41 | −5.45 |
| Dominance index | 0.41 | 0.51 |
| Reciprocal Simpson index | 0.75 | −0.54 |
| Shannon index | 0.39 | 0.56 |
| Menhinick index | 0.37 | 0.57 |
| Buzas and Gibson’s index | 0.35 | 0.34 |
| Equitability index | 0.40 | 0.43 |
| Berger-Parker Dominance index | 0.35 | −2.41 |
| Inverted Berger-Parker Dominance index | 0.17 | 1.63 |
| Margalef Richness index | 0.38 | 0.59 |
| Gini coefficient | 0.34 | −2.46 |
| B: FLC T-cell repertoire | ||
| Diversity Metric | p Value | Log2FC |
| Biodiversity | 0.99 | 0.00 |
| Simpson’s index | 0.73 | −0.71 |
| Dominance index | 0.73 | 0.10 |
| Reciprocal Simpson index | 0.65 | −0.65 |
| Shannon index | 0.90 | 0.05 |
| Menhinick index | 0.81 | 0.10 |
| Buzas and Gibson’s index | 0.90 | 0.06 |
| Equitability index | 0.77 | 0.08 |
| Berger-Parker Dominance index | 0.84 | −0.24 |
| Inverted Berger-Parker Dominance index | 0.56 | 0.81 |
| Margalef Richness index | 0.99 | 0.01 |
| Gini coefficient | 0.92 | −0.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haigh, O.L.; Grant, E.J.; Nguyen, T.H.O.; Kedzierska, K.; Field, M.A.; Miles, J.J. Genetic Bias, Diversity Indices, Physiochemical Properties and CDR3 Motifs Divide Auto-Reactive from Allo-Reactive T-Cell Repertoires. Int. J. Mol. Sci. 2021, 22, 1625. https://doi.org/10.3390/ijms22041625
Haigh OL, Grant EJ, Nguyen THO, Kedzierska K, Field MA, Miles JJ. Genetic Bias, Diversity Indices, Physiochemical Properties and CDR3 Motifs Divide Auto-Reactive from Allo-Reactive T-Cell Repertoires. International Journal of Molecular Sciences. 2021; 22(4):1625. https://doi.org/10.3390/ijms22041625
Chicago/Turabian StyleHaigh, Oscar L., Emma J. Grant, Thi H. O. Nguyen, Katherine Kedzierska, Matt A. Field, and John J. Miles. 2021. "Genetic Bias, Diversity Indices, Physiochemical Properties and CDR3 Motifs Divide Auto-Reactive from Allo-Reactive T-Cell Repertoires" International Journal of Molecular Sciences 22, no. 4: 1625. https://doi.org/10.3390/ijms22041625
APA StyleHaigh, O. L., Grant, E. J., Nguyen, T. H. O., Kedzierska, K., Field, M. A., & Miles, J. J. (2021). Genetic Bias, Diversity Indices, Physiochemical Properties and CDR3 Motifs Divide Auto-Reactive from Allo-Reactive T-Cell Repertoires. International Journal of Molecular Sciences, 22(4), 1625. https://doi.org/10.3390/ijms22041625