
Adipokines, Myokines, and Hepatokines: Crosstalk and Metabolic Repercussions


 ,
,
Abstract
:
1. Introduction
2. Discussion
2.1. Adipokines
2.2. Myokines
2.3. Hepatokines
2.4. Adipokines, Myokines and Hepatokines: Crosstalk and Metabolic Repercussions
3. Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huber, B.C.; Steffen, J.; Schlichtiger, J.; Brunner, S. Altered nutrition behavior during COVID-19 pandemic lockdown in young adults. Eur. J. Nutr. 2020. [Google Scholar] [CrossRef]
- Sofra, X.; Badami, S.J.H. Adverse effects of sedentary lifestyles: Inflammation, and high-glucose induced oxidative stress—A double blind randomized clinical trial on diabetic and prediabetic patients. Health 2020, 12, 1029. [Google Scholar] [CrossRef]
- Sandhu, R.; Chandora, C.J.T.U. Cure of obesity in homeopathy: An approach to control a global epidemic. Tantian Univ. J. Homoeopath. Med. Sci. 2020, 1, 17–22. [Google Scholar]
- Alloubani, A.; Nimer, R.; Samara, R. Relationship between hyperlipidemia, cardiovascular disease and stroke: A systematic review. Curr. Cardiol. Rev. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bergamini, M.; Iora, P.H.; Rocha, T.A.H.; Tchuisseu, Y.P.; Dutra, A.C.; Scheidt, J.; Nihei, O.K.; de Barros Carvalho, M.D.; Staton, C.A.; Vissoci, J.R.N.; et al. Mapping risk of ischemic heart disease using machine learning in a Brazilian state. PLoS ONE 2020, 15, e0243558. [Google Scholar] [CrossRef] [PubMed]
- Précoma, D.B.; Oliveira, G.M.M.; Simão, A.F.; Dutra, O.P.; Coelho, O.R.; Izar, M.C.O. Atualização da diretriz de prevenção cardiovascular da sociedade brasileira de cardiologia. Arq. Bras. Cardiol. 2019, 113, 787–891. [Google Scholar] [PubMed]
- Jia, T.; Wang, C.; Han, Z.; Wang, X.; Ding, M.; Wang, Q. Experimental rodent models of cardiovascular diseases. Front. Cardiovasc. Med. 2020, 7, 588075. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Flato, U.A.P.; Tofano, R.J.; Goulart, R.A.; Guiguer, E.L.; Detregiachi, C.R.P.; Buchaim, D.V.; Araújo, A.C.; Buchaim, R.L.; Reina, F.T.R.; et al. Physical exercise and myokines: Relationships with sarcopenia and cardiovascular complications. Int. J. Mol. Sci. 2020, 21, 3607. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.Y.; Lim, K.K.; Omar, M.A.; Mohd Yusoff, M.F.; Sooryanarayana, R.; Ahmad, N.A.; Abd Razak, M.A.; Baharudin Shaharuddin, A.; Mahmud, N.A.; Mahmud, M.A.F.; et al. Prevalence and factors associated with physical inactivity among older adults in Malaysia: A cross-sectional study. Geriatr. Gerontol. Int. 2020, 20 (Suppl. 2), 49–56. [Google Scholar] [CrossRef] [PubMed]
- Faramia, J.; Ostinelli, G.; Drolet-Labelle, V.; Picard, F.; Tchernof, A. Metabolic adaptations after bariatric surgery: Adipokines, myokines and hepatokines. Curr. Opin. Pharmacol. 2020, 52, 67–74. [Google Scholar] [CrossRef]
- Gonzalez-Gil, A.M.; Elizondo-Montemayor, L. The role of exercise in the interplay between myokines, hepatokines, osteokines, adipokines, and modulation of inflammation for energy substrate redistribution and fat mass loss: A review. Nutrients 2020, 12, 1899. [Google Scholar] [CrossRef] [PubMed]
- Frithioff-Bøjsøe, C.; Lund, M.A.; Lausten-Thomsen, U.; Hedley, P.L.; Pedersen, O.; Christiansen, M.; Baker, J.L.; Hansen, T.; Holm, J.C.J.P.D. Leptin, adiponectin, and their ratio as markers of insulin resistance and cardiometabolic risk in childhood obesity. Pediatr. Diab. 2020, 21, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Kwon, Y.; Joo, J.Y.; Kim, D.G.; Yoon, J.H. Secretomics to discover regulators in diseases. Int. J. Mol. Sci. 2019, 20, 3893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, K.J.; Lee, D.S.; Kim, W.K.; Han, B.S.; Lee, S.C.; Bae, K.H. Metabolic adaptation in obesity and type II diabetes: Myokines, adipokines and hepatokines. Int. J. Mol. Sci. 2016, 18, 8. [Google Scholar] [CrossRef] [Green Version]
- Musovic, S.; Shrestha, M.M.; Komai, A.M.; Olofsson, C.S. Resistin is co-secreted with adiponectin in white mouse adipocytes. Biochem. Biophys. Res. Commun. 2020, 534, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Duerrschmid, C.; He, Y.; Wang, C.; Li, C.; Bournat, J.C.; Romere, C.; Saha, P.K.; Lee, M.E.; Phillips, K.J.; Jain, M.; et al. Asprosin is a centrally acting orexigenic hormone. Nat. Med. 2017, 23, 1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romere, C.; Duerrschmid, C.; Bournat, J.; Constable, P.; Jain, M.; Xia, F.; Saha, P.K.; Del Solar, M.; Zhu, B.; York, B.; et al. Asprosin, a fasting-induced glucogenic protein hormone. Cell 2016, 165, 566–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Fan, J.; Su, Q.; Yang, Z. Cytokines and abnormal glucose and lipid metabolism. Front. Endocrinol. 2019, 10, 703. [Google Scholar] [CrossRef]
- Watanabe, T.; Watanabe-Kominato, K.; Takahashi, Y.; Kojima, M.; Watanabe, R. Adipose tissue-derived omentin-1 function and regulation. Compr. Physiol. 2017, 7, 765–781. [Google Scholar] [CrossRef]
- Cuevas-Ramos, D.; Mehta, R.; Aguilar-Salinas, C.A. Fibroblast growth factor 21 and browning of white adipose tissue. Front. Physiol. 2019, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Pan, Y.; Yang, G.; Cui, Z.; Yu, W.; Liu, H.; Bai, B. Sfrp5/Wnt5a and leptin/adiponectin levels in the serum and the periarterial adipose tissue of patients with peripheral arterial occlusive disease. Clin. Biochem. 2020, 87, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Cakirca, G.; Turgut, F.H. Serum matrix metalloproteinase-9, tissue inhibitor of metalloproteinase-1 and matrix metalloproteinase-9/ neutrophil gelatinase-associated lipocalin complex levels in patients with early-stage diabetic nephropathy. Iran. J. Kidney Dis. 2018, 12, 299. [Google Scholar]
- Recinella, L.; Orlando, G.; Ferrante, C.; Chiavaroli, A.; Brunetti, L.; Leone, S. Adipokines: New potential therapeutic target for obesity and metabolic, rheumatic, and cardiovascular diseases. Front. Physiol. 2020, 11, 578966. [Google Scholar] [CrossRef] [PubMed]
- Pazgan-Simon, M.; Zuwała-Jagiełło, J.; Kukla, M.; Grzebyk, E.; Simon, K. Serum concentrations of selected adipokines in virus-related liver cirrhosis and hepatocellular carcinoma. Clin. Exp. Hepatol. 2020, 6, 235–242. [Google Scholar] [CrossRef]
- Zieger, K.; Weiner, J.; Krause, K.; Schwarz, M.; Kohn, M.; Stumvoll, M.; Blüher, M.; Heiker, J.T. Vaspin suppresses cytokine-induced inflammation in 3T3-L1 adipocytes via inhibition of NFκB pathway. Mol. Cell. Endocrinol. 2018, 460, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Horak, M.; Kuruczova, D.; Zlamal, F.; Tomandl, J.; Bienertova-Vasku, J. Follistatin-like 1 is downregulated in morbidly and super obese central-European population. Dis. Mark. 2018, 2018, 4140815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Choi, M.J.; Kim, S.W.; Yun, J.W. Secreted protein acidic and rich in cysteine (SPARC) regulates thermogenesis in white and brown adipocytes. Mol. Cell. Endocrinol. 2020, 506, 110757. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Seldin, M.M.; Little, H.C.; Choy, N.; Klonisch, T.; Wong, G.W. C1q/TNF-related protein 6 (CTRP6) links obesity to adipose tissue inflammation and insulin resistance. J. Biol. Chem. 2017, 292, 14836–14850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarzour, A.; Kim, H.W.; Weintraub, N.L. Understanding obesity-related cardiovascular disease: It’s all about balance. Circulation 2018, 138, 64–66. [Google Scholar] [CrossRef]
- Barchetta, I.; Cimini, F.A.; Capoccia, D.; De Gioannis, R.; Porzia, A.; Mainiero, F.; Di Martino, M.; Bertoccini, L.; De Bernardinis, M.; Leonetti, F.; et al. WISP1 is a marker of systemic and adipose tissue inflammation in dysmetabolic subjects with or without type 2 diabetes. J. Endocr. Soc. 2017, 1, 660–670. [Google Scholar] [CrossRef] [Green Version]
- Abella, V.; Scotece, M.; Conde, J.; Pino, J.; Gonzalez-Gay, M.A.; Gómez-Reino, J.J.; Mera, A.; Lago, F.; Gómez, R.; Gualillo, O. Leptin in the interplay of inflammation, metabolism and immune system disorders. Nat Rev. Rheumatol. 2017, 13, 100–109. [Google Scholar] [CrossRef]
- Korolczuk, A.; Bełtowski, J. Progranulin, a new adipokine at the crossroads of metabolic syndrome, diabetes, dyslipidemia and hypertension. Curr. Pharm. Design 2017, 23, 1533–1539. [Google Scholar] [CrossRef]
- Franco-Trepat, E.; Guillán-Fresco, M.; Alonso-Pérez, A.; Jorge-Mora, A.; Francisco, V.; Gualillo, O.; Gómez, R. Visfatin connection: Present and future in osteoarthritis and osteoporosis. J. Clin. Med. 2019, 8, 1178. [Google Scholar] [CrossRef] [Green Version]
- Kumari, B.; Yadav, U.C.S. Adipokine visfatin’s role in pathogenesis of diabesity and related metabolic derangements. Curr. Mol. Med. 2018, 18, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Khadir, A.; Kavalakatt, S.; Madhu, D.; Tiss, A. Fetuin-a expression profile in mouse and human adipose tissue. Lip. Health Dis. 2020, 19, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severo, J.S.; Morais, J.B.S.; Beserra, J.B.; dos Santos, L.R.; de Sousa Melo, S.R.; de Sousa, G.S.; de Matos Neto, E.M.; Henriques, G.S.; do Nascimento Marreiro, D. Role of Zinc in Zinc-α2-Glycoprotein Metabolism in obesity: A review of literature. Biol. Trace Element Res. 2020, 193, 81–88. [Google Scholar] [CrossRef]
- Wang, L.; Liu, M.; Ning, D.; Zhu, H.; Shan, G.; Wang, D.; Ping, B.; Yu, Y.; Yang, H.; Yan, K.; et al. Low serum ZAG levels correlate with determinants of the metabolic syndrome in Chinese subjects. Front. Endocrinol. 2020, 11, 154. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Qiao, Y.; Qi, F.; Shen, Q.; Zhao, R.; Yang, X. Zinc-α2-glycoprotein knockout influenced genes expression profile in adipose tissue and decreased the lipid mobilizing after dexamethasone treatment in mice. Horm. Metab. Res. 2020, 52, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Scheja, L.; Heeren, J. The endocrine function of adipose tissues in health and cardiometabolic disease. Nat. Rev. Endocrinol. 2019, 15, 507–524. [Google Scholar] [CrossRef]
- Merli, M.; Lattanzi, B.; Aprile, F. Sarcopenic obesity in fatty liver. Curr. Opin. Clin. Nutr. Metab. Care 2019, 22, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Aronis, K.N.; Kountouras, J.; Raptis, D.D.; Vasiloglou, M.F.; Mantzoros, C.S.J.D. Circulating leptin in non-alcoholic fatty liver disease: A systematic review and meta-analysis. Diabetology 2016, 59, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, I.S.; Jebb, S.A.; Langmack, G.; Lawrence, E.; Cheetham, C.H.; Prentice, A.M.; Hughes, I.A.; McCamish, M.A.; O’Rahilly, S.J.N.E. Effects of recombinant leptin therapy in a child with congenital leptin deficiency. N. Engl. J. Med. 1999, 341, 879–884. [Google Scholar] [CrossRef]
- Moon, H.-S.; Matarese, G.; Brennan, A.M.; Chamberland, J.P.; Liu, X.; Fiorenza, C.G.; Mylvaganam, G.H.; Abanni, L.; Carbone, F.; Williams, C.J.J.D. Efficacy of metreleptin in obese patients with type 2 diabetes: Cellular and molecular pathways underlying leptin tolerance. Diabetes 2011, 60, 1647–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buechler, C.; Haberl, E.M.; Rein-Fischboeck, L.; Aslanidis, C. Adipokines in liver cirrhosis. Int. J. Mol. Sci. 2017, 18, 1392. [Google Scholar] [CrossRef] [Green Version]
- Mikolasevic, I.; Pavic, T.; Kanizaj, T.F.; Bender, D.V.; Domislovic, V.; Krznaric, Z. Nonalcoholic fatty liver disease and sarcopenia: Where do we stand? Can. J. Gastroenterol. Hepatol. 2020, 2020, 8859719. [Google Scholar] [CrossRef]
- Fang, H.; Judd, R.L. Adiponectin regulation and function. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2018; Volume 8, pp. 1031–1063. [Google Scholar]
- Choi, S.-K.; Kwon, Y.; Byeon, S.; Haam, C.E.; Lee, Y.-H. AdipoRon, adiponectin receptor agonist, improves vascular function in the mesenteric arteries of type 2 diabetic mice. PLoS ONE 2020, 15, e0230227. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Liu, Z.; Liu, G.; Zhao, S.; Li, C.; Chen, W.; Coban Akdemir, Z.; Lin, J.; Song, X.; Wang, S.; et al. Genetic and molecular mechanism for distinct clinical phenotypes conveyed by allelic truncating mutations implicated in FBN1. Mol. Genet. Genom. Med. 2020, 8, e1023. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.A.; Choi, K.M.J.F.i.P. Newly discovered adipokines: Pathophysiological link between obesity and cardiometabolic disorders. Front. Physiol. 2020, 11, 568800. [Google Scholar] [CrossRef]
- Gozel, N.; Kilinc, F. Investigation of plasma asprosin and saliva levels in newly diagnosed type 2 diabetes mellitus patients treated with metformin. Endokrynol. Polska 2020. [Google Scholar] [CrossRef]
- Wen, M.S.; Wang, C.Y.; Yeh, J.K.; Chen, C.C.; Tsai, M.L.; Ho, M.Y.; Hung, K.C.; Hsieh, I.C. The role of Asprosin in patients with dilated cardiomyopathy. BMC Cardiovasc. Dis. 2020, 20, 402. [Google Scholar] [CrossRef]
- Yuan, M.; Li, W.; Zhu, Y.; Yu, B.; Wu, J. Asprosin: A novel player in metabolic diseases. Front. Endocrinol. 2020, 11, 64. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, T.; Church, C.; Tsintzas, K.; Jones, R.; Breen, L.; Davis, E.T.; Baker, D.J.; Jones, S.W. Vaspin promotes insulin sensitivity of elderly muscle and is upregulated in obesity. J. Endocrinol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K.; Shirai, R.; Yamaguchi, M.; Yamashita, T.; Shibata, K.; Okano, T.; Mori, Y.; Matsuyama, T.A.; Ishibashi-Ueda, H.; Hirano, T.; et al. Anti-atherogenic effects of vaspin on human aortic smooth muscle cell/macrophage responses and hyperlipidemic mouse plaque phenotype. Int. J. Mol. Sci. 2018, 19, 1732. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Li, Y.; Esangbedo, I.C.; Li, G.; Feng, D.; Li, L.; Xu, L.; Han, L.; Li, M.; Li, C.; et al. Circulating osteonectin and adipokine profiles in relation to metabolically healthy obesity in Chinese children: Findings from BCAMS. J. Am. Heart Assoc. 2018, 7, e009169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbalho, S.M.; Prado Neto, E.V.; De Alvares Goulart, R.; Bechara, M.D.; Baisi Chagas, E.F.; Audi, M.; Guissoni Campos, L.M.; Landgraf Guiger, E.; Buchaim, R.L.; Buchaim, D.V.; et al. Myokines: A descriptive review. J. Sports Med. Phys. Fitness 2020, 60, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Mikolajczak, A.; Oswiecimska, J.M.; Swietochowska, E.; Roczniak, W.; Ziora, K.T. Serum FGF21 in girls with anorexia nervosa—Comparison to normal weight and obese female adolescents. Neuro Endocrinol. Lett. 2017, 38, 173–181. [Google Scholar] [PubMed]
- Kwon, J.H.; Moon, K.M.; Min, K.W. Exercise-induced myokines can explain the importance of physical activity in the elderly: An overview. Healthcare 2020, 8, 378. [Google Scholar] [CrossRef]
- Chung, H.S.; Choi, K.M. Organokines in disease. Adv. Clin. Chem. 2020, 94, 261–321. [Google Scholar] [CrossRef]
- Aoi, W.; Hirano, N.; Lassiter, D.G.; Bjornholm, M.; Chibalin, A.V.; Sakuma, K.; Tanimura, Y.; Mizushima, K.; Takagi, T.; Naito, Y.; et al. Secreted protein acidic and rich in cysteine (SPARC) improves glucose tolerance via AMP-activated protein kinase activation. FASEB J. 2019, 33, 10551–10562. [Google Scholar] [CrossRef] [Green Version]
- Cooper, C.; Moon, H.Y.; van Praag, H. On the run for hippocampal plasticity. Cold Spring Harbor Perspect. Med. 2018, 8, a029736. [Google Scholar] [CrossRef]
- Gholamrezayi, A.; Mohamadinarab, M.; Rahbarinejad, P.; Fallah, S.; Barez, S.R.; Setayesh, L.; Moradi, N.; Fadaei, R.; Chamani, E.; Tavakoli, T. Characterization of the serum levels of Meteorin-like in patients with inflammatory bowel disease and its association with inflammatory cytokines. Lip. Health Dis. 2020, 19, 230. [Google Scholar] [CrossRef]
- Domin, R.; Dadej, D.; Pytka, M.; Zybek-Kocik, A.; Ruchała, M.; Guzik, P. Effect of various exercise regimens on selected Exercise-induced cytokines in healthy people. Int. J. Environ. Res. Public Health 2021, 18, 1261. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.M. The impact of organokines on insulin resistance, inflammation, and atherosclerosis. Endocrinol. Metab. 2016, 31, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Yang, L.; Yan, S.; Zheng, H.; Liang, L.; Cai, X.; Liao, M. Fetuin A promotes lipotoxicity in β cells through the TLR4 signaling pathway and the role of pioglitazone in anti-lipotoxicity. Mol. Cell. Endocrinol. 2015, 412, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ruth, C.M.; Andrew, J.H.; Morris, A.; Russell, D.B.; Jennifer, C.Y.L.; Burke, M.; Robert, J.A.G.; Bronwyn, A.K.; Michael, J.K.; Mark, A.F.; et al. Fetuin B is a secreted hepatocyte factor linking steatosis to impaired glucose metabolism. Cell Metab. 2015, 22, 1078–1089. [Google Scholar] [CrossRef] [Green Version]
- Meex, R.C.R.; Watt, M.J. Hepatokines: Linking nonalcoholic fatty liver disease and insulin resistance. Nat. Rev. Endocrinol. 2017, 13, 509–520. [Google Scholar] [CrossRef]
- Hashimoto, O.; Funaba, M.; Sekiyama, K.; Doi, S.; Shindo, D.; Satoh, R.; Itoi, H.; Oiwa, H.; Morita, M.; Suzuki, C.; et al. Activin E controls energy homeostasis in both brown and white adipose tissues as a hepatokine. Cell Rep. 2018, 25, 1193–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, M.; Kikuchi, A.; Misu, H.; Igawa, H.; Ashihara, M.; Kushima, Y.; Honda, K.; Suzuki, Y.; Kawabe, Y.; Kaneko, S.; et al. Inhibin βE (INHBE) is a possible insulin resistance-associated hepatokine identified by comprehensive gene expression analysis in human liver biopsy samples. PLoS ONE 2018, 13, e0194798. [Google Scholar] [CrossRef] [Green Version]
- Lebensztejn, D.M.; Flisiak-Jackiewicz, M.; Białokoz-Kalinowska, I.; Bobrus-Chociej, A.; Kowalska, I. Hepatokines and non-alcoholic fatty liver disease. Acta Biochim. Polonica 2016, 63, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Simó, R.; Sáez-López, C.; Barbosa-Desongles, A.; Hernández, C.; Selva, D.M. Novel insights in SHBG regulation and clinical implications. Trends Endocrinol. Metab. 2015, 26, 376–383. [Google Scholar] [CrossRef]
- Pohl, R.; Haberl, E.M.; Rein-Fischboeck, L.; Zimny, S.; Neumann, M.; Aslanidis, C.; Schacherer, D.; Krautbauer, S.; Eisinger, K.; Weiss, T.S.; et al. Hepatic chemerin mRNA expression is reduced in human nonalcoholic steatohepatitis. Eur. J. Clin. Investig. 2017, 47, 7–18. [Google Scholar] [CrossRef]
- Misu, H.; Takayama, H.; Saito, Y.; Mita, Y.; Kikuchi, A.; Ishii, K.A.; Chikamoto, K.; Kanamori, T.; Tajima, N.; Lan, F.; et al. Deficiency of the hepatokine selenoprotein P increases responsiveness to exercise in mice through upregulation of reactive oxygen species and AMP-activated protein kinase in muscle. Nat. Med. 2017, 23, 508–516. [Google Scholar] [CrossRef]
- Perakakis, N.; Mougios, V.; Fatouros, I.; Siopi, A.; Draganidis, D.; Peradze, N.; Ghaly, W.; Mantzoros, C.S. Physiology of activins/follistatins: Associations with metabolic and anthropometric variables and response to exercise. J. Clin. Endocrinol. Metab. 2018, 103, 3890–3899. [Google Scholar] [CrossRef] [Green Version]
- Ennequin, G.; Sirvent, P.; Whitham, M. Role of exercise-induced hepatokines in metabolic disorders. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E11–E24. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Feng, M.; Zhang, S.; Dong, Z.; Wang, Y.; Zhang, W.; Liu, C. Angptl8 mediates food-driven resetting of hepatic circadian clock in mice. Nat. Commun. 2019, 10, 3518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.T.; Ou, H.Y.; Hung, H.C.; Su, Y.C.; Lu, F.H.; Wu, J.S.; Yang, Y.C.; Wu, C.L.; Chang, C.J. A novel hepatokine, HFREP1, plays a crucial role in the development of insulin resistance and type 2 diabetes. Diabetologia 2016, 59, 1732–1742. [Google Scholar] [CrossRef] [Green Version]
- Mouchiroud, M.; Camiré, É.; Aldow, M.; Caron, A.; Jubinville, É.; Turcotte, L.; Kaci, I.; Beaulieu, M.-J.; Roy, C.; Labbé, S.M.; et al. The hepatokine Tsukushi is released in response to NAFLD and impacts cholesterol homeostasis. JCI Insight 2019, 4, e129492. [Google Scholar] [CrossRef] [Green Version]
- Iroz, A.; Couty, J.-P.; Postic, C. Hepatokines: Unlocking the multi-organ network in metabolic diseases. Diabetologia 2015, 58, 1699–1703. [Google Scholar] [CrossRef] [PubMed]
- Glück, M.; Glück, J.; Wiewióra, M.; Rogala, B.; Piecuch, J. Serum irisin, adropin, and preptin in obese patients 6 months after bariatric surgery. Obesity Surg. 2019, 29, 3334–3341. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.P.; Xu, W.T.; Wang, L.; You, T.; Chan, S.P.; Zhao, X.; Yang, X.J. Serum adropin level in patients with stable coronary artery disease. Heart Lung Circulat. 2015, 24, 975–979. [Google Scholar] [CrossRef]
- Hansen, J.S.; Pedersen, B.K.; Xu, G.; Lehmann, R.; Weigert, C.; Plomgaard, P. Exercise-induced secretion of FGF21 and follistatin are blocked by pancreatic clamp and impaired in type 2 diabetes. J. Clin. Endocrinol. Metab. 2016, 101, 2816–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigert, C.; Hoene, M.; Plomgaard, P. Hepatokines—A novel group of exercise factors. Eur. J. Physiol. 2019, 471, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, B.K. Physical activity and muscle-brain crosstalk. Nat. Rev. Endocrinol. 2019, 15, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Moon, J.; Jo, G.; Lee, J.; Kim, O.Y.; Oh, H.; Lim, H.; Shin, M.-J. The association between genetic variants of angiopoietin-like 3 and risk of diabetes mellitus is modified by dietary factors in Koreans. Sci. Rep. 2019, 9, 766. [Google Scholar] [CrossRef]
- Misu, H. Pathophysiological significance of hepatokine overproduction in type 2 diabetes. Diabetol. Int. 2018, 9, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Chikamoto, K.; Misu, H.; Takayama, H.; Kikuchi, A.; Ishii, K.A.; Lan, F.; Takata, N.; Tajima-Shirasaki, N.; Takeshita, Y.; Tsugane, H.; et al. Rapid response of the steatosis-sensing hepatokine LECT2 during diet-induced weight cycling in mice. Biochem. Biophys. Res. Commun. 2016, 478, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Cody, S.O.; Potthoff, M.J. Hepatokines and metabolism: Deciphering communication from the liver. Mol. Metab. 2021, 44, 101138. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, D.; Fu, J.; Zhao, L.; Li, D.; Sun, H.; Liu, X.; Xu, J.; Tian, T.; Zhang, L.; et al. Tsukushi is a novel prognostic biomarker and correlates with tumor-infiltrating B cells in non-small cell lung cancer. Aging 2021, 12, 4428–4451. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Pan, R.; Cheng, J.; Cui, Q.; Chen, J.; Yuan, Z. Sodium rutin extends lifespan and health span in mice including positive impacts on liver health. Br. J. Pharmacol. 2021. [Google Scholar] [CrossRef]
- Hou, G.; Tang, Y.; Ren, L.; Guan, Y.; Hou, X.; Song, G. The ANGPTL8 rs2278426 (C/T) polymorphism is associated with prediabetes and type 2 diabetes in a han Chinese population in Hebei province. Int. J. Endocrinol. 2020, 2020, 1621239. [Google Scholar] [CrossRef]
- Li, F.; Li, Y.; Duan, Y.; Hu, C.A.; Tang, Y.; Yin, Y. Myokines and adipokines: Involvement in the crosstalk between skeletal muscle and adipose tissue. Cytokine Growth Factor Rev. 2017, 33, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhou, M.; Wang, X.; Li, S.; Yang, D. The role of myokines and adipokines in hypertension and hypertension-related complications. Hypertens. Res. 2019, 42, 1544–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta-Delgado, A.S.; Roffe-Vazquez, D.N.; Gonzalez-Gil, A.M.; Villarreal-Calderón, J.R.; Tamez-Rivera, O.; Rodriguez-Gutierrez, N.A.; Castillo, E.C.; Silva-Platas, C.; Garcia-Rivas, G.; Elizondo-Montemayor, L. Serum irisin levels, endothelial dysfunction, and inflammation in pediatric patients with type 2 diabetes mellitus and metabolic syndrome. J. Diab. Res. 2020, 2020, 1949415. [Google Scholar] [CrossRef]
- Leal, L.G.; Lopes, M.A.; Batista, M.L., Jr. Physical exercise-induced myokines and muscle-adipose tissue crosstalk: A review of current knowledge and the implications for health and metabolic diseases. Front. Physiol. 2018, 9, 1307. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Becerril, S.; Ezquerro, S.; Méndez-Giménez, L.; Frühbeck, G. Crosstalk between adipokines and myokines in fat browning. Acta Physiol. 2017, 219, 362–381. [Google Scholar] [CrossRef] [PubMed]
- Gomarasca, M.; Banfi, G.; Lombardi, G. Chapter four—Myokines: The endocrine coupling of skeletal muscle and bone. In Advances in Clinical Chemistry; Makowski, G.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 94, pp. 155–218. [Google Scholar]
- Kirk, B.; Feehan, J.; Lombardi, G.; Duque, G. Muscle, bone, and fat crosstalk: The biological role of myokines, osteokines, and adipokines. Curr. Osteopor. Rep. 2020, 18, 388–400. [Google Scholar] [CrossRef]
- Batsis, J.A.; Villareal, D.T. Sarcopenic obesity in older adults: Aetiology, epidemiology and treatment strategies. Nat. Rev. Endocrinol. 2018, 14, 513–537. [Google Scholar] [CrossRef] [PubMed]
- Hirschfeld, H.P.; Kinsella, R.; Duque, G. Osteosarcopenia: Where bone, muscle, and fat collide. Osteopor. Int. 2017, 28, 2781–2790. [Google Scholar] [CrossRef]
- Chen, W.; Wang, L.; You, W.; Shan, T. Myokines mediate the cross talk between skeletal muscle and other organs. J. Cell. Physiol. 2020, 236, 2393–2412. [Google Scholar] [CrossRef]
- Díaz, B.B.; González, D.A.; Gannar, F.; Pérez, M.C.R.; de León, A.C. Myokines, physical activity, insulin resistance and autoimmune diseases. Immunol. Lett. 2018, 203, 1–5. [Google Scholar] [CrossRef]
- Byun, K.; Lee, S. The potential role of irisin in vascular function and atherosclerosis: A review. Int. J. Mol. Sci. 2020, 21, 7184. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Rosa, S.C.; Nayak, N.; Caymo, A.M.; Gordon, J.W. Mechanisms of muscle insulin resistance and the cross-talk with liver and adipose tissue. Physiol. Rep. 2020, 8, e14607. [Google Scholar] [CrossRef] [PubMed]
- Goedeke, L.; Perry, R.J.; Shulman, G.I. Emerging pharmacological targets for the treatment of nonalcoholic fatty liver disease, insulin resistance, and type 2 diabetes. Ann. Rev. Pharmacol. Toxicol. 2019, 59, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Degirolamo, C.; Sabbà, C.; Moschetta, A. Therapeutic potential of the endocrine fibroblast growth factors FGF19, FGF21 and FGF23. Nat. Rev. Drug Disc. 2016, 15, 51–69. [Google Scholar] [CrossRef]
- Klein Hazebroek, M.; Keipert, S. Adapting to the cold: A role for endogenous fibroblast growth factor 21 in thermoregulation? Front. Endocrinol. 2020, 11, 389. [Google Scholar] [CrossRef]
- Lee, C.H.; Lam, K.S. Obesity-induced insulin resistance and macrophage infiltration of the adipose tissue: A vicious cycle. J. Diabetes Investig. 2018, 10, 29–31. [Google Scholar] [CrossRef] [PubMed]
- Soares, P.E.M.; Ferreira, E.T.d.O.; Mafra, G.C.D.S.; Silva, J.G.D.; Santos, R.L.D.; Farias, J.V.C.; Farias, I.C.C. The role of adipokines in the Immunology of obesity. Res. Soc. Dev. 2020, 9, e935975095. [Google Scholar] [CrossRef]
- Ouerghi, N.; Ben Fradj, M.K.; Talbi, E.; Bezrati, I.; Feki, M.; Bouassida, A. Association of selected adipokines with metabolic syndrome and cardio-metabolic risk factors in young males. Cytokine 2020, 133, 155170. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.S.; Rutti, S.; Arous, C.; Clemmesen, J.O.; Secher, N.H.; Drescher, A.; Gonelle-Gispert, C.; Halban, P.A.; Pedersen, B.K.; Weigert, C.; et al. Circulating follistatin is liver-derived and regulated by the glucagon-to-insulin ratio. J. Clin. Endocrinol. Metab. 2016, 101, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Mushala, B.A.S.; Scott, I. Adropin: A hepatokine modulator of vascular function and cardiac fuel metabolism. Am. J. Physiol. Circ. Physiol. 2021, 320, H238–H244. [Google Scholar] [CrossRef]
- Chen, X.; Sun, X.; Shen, T.; Chen, Q.; Chen, S.; Pang, J.; Mi, J.; Tang, Y.; You, Y.; Xu, H.; et al. Lower adropin expression is associated with oxidative stress and severity of nonalcoholic fatty liver disease. Free Radic. Biol. Med. 2020, 160. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Wang, Y.; Shu, P.; Zhou, Q.; Zhu, J.; Zhou, W.; Du, C.; Xu, C.; Liu, X.; Tang, L. Increased serum levels of fetuin B in patients with coronary artery disease. Endocrine 2017, 58, 97–105. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adipokine | Stimulation for Its Increase | Metabolic Action | Reference |
|---|---|---|---|
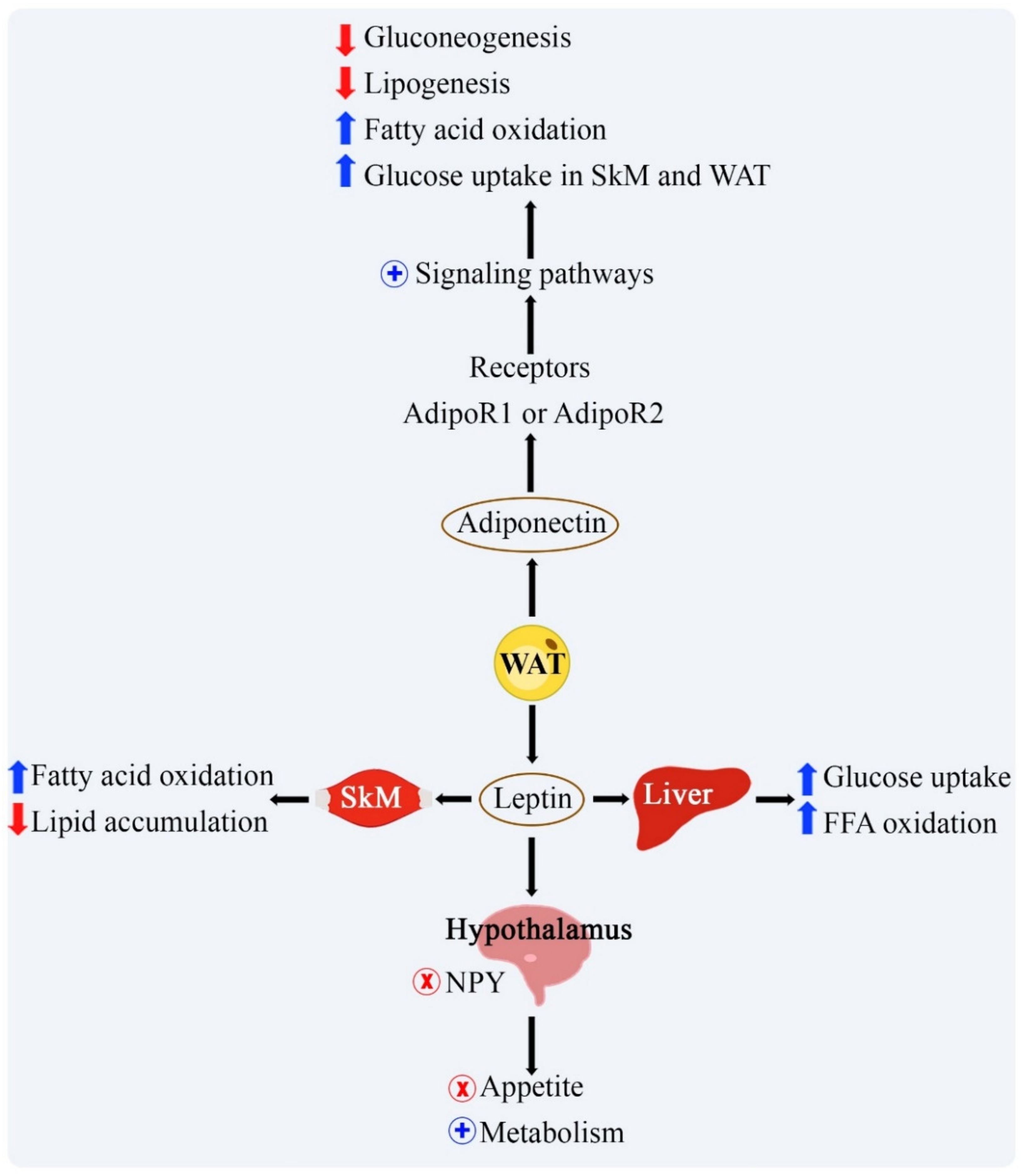
| Leptin | Increase in fat mass. | In the immune system, it acts to increase pro-inflammatory cytokines. In the CNS, it promotes a decrease in food intake and an increase in global energy expenditure. In skeletal muscle, it acts in the absorption and oxidation of glucose and FFA. In the liver it increases the oxidation of fatty acids and reduces the accumulation of lipids. | [11] |
| Adiponectina | Adrenergic beta signaling; increase in FGF21, IL-15, and irisin induced by physical exercise. | In the immune system it has anti-inflammatory actions. In the CNS it promotes an increase in food intake and a reduction in hypothalamic inflammation. In the liver and skeletal muscle, it increases fatty acid oxidation and insulin sensitivity. | [11] |
| Resistin | Increase in fat mass. | Immune system: pro-inflammatory actions. It acts in endothelial dysfunction, CVD and inhibition of insulin signaling through the suppressor of cytokine signaling 3 (SOCS3). | [14,15] |
| IL-6 | Activation of the nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB). | Adipose tissue: proinflammatory action, and acts in inhibiting the expression of insulin receptor substrate 1 (IRS1) and glucose transporter type 4 GLUT4) in adipocytes. | [14] |
| Asprosin | Induced by fasting and produced by white adipose tissue in obese people with DM2. | It increases food consumption and body weight and accelerates the production of liver glucose. | [16,17] |
| Chemerin | Inflammatory and coagulation serine proteases. | It accentuates glucose intolerance and makes insulin signaling difficult. | [18] |
| Omentin | Increase in FGF21 and dexamethasone. | Optimizes the action of insulin and, consequently, the absorption of glucose. It also acts as an anti-atherosclerotic factor. | [19] |
| FGF21 | Exposure to cold and physical exercise. | It acts in the browning of WAT, lipid oxidation and thermogenesis, and stimulates the expression of adiponectin in the bloodstream. | [20] |
| SFRP5 | Induced during the proliferation, differentiation and maturation of pre-adipocytes. | Regulates the expression of pro-inflammatory cytokines by inhibiting the Wingless-type family member 5a signaling (Wnt5a), non-canonical Wnt family. | [14,21] |
| Lipocalin 2 | Low-level systemic inflammation in obese patients with metabolic syndrome. | Regulation of inflammation and the transport of fatty acids and iron. It is associated with CVD, vascular remodeling and instability of atherosclerotic plaques. | [22,23] |
| Vaspin | Increase in fat mass. | Reduces the synthesis of pro-inflammatory cytokines. It improves glucose intolerance and insulin sensitivity and protects the vascular tissues from fatty acid-induced apoptosis. | [24,25] |
| FSTL1 | It is expressed in large quantities by adipose tissue in the state of low-grade chronic inflammation. | Lower levels of FSTL1 are associated with super obesity due to loss of adipogenesis, increased maturated adipocytes, cellular senescence and anti-apoptotic FSLT1 reduction. It has a pro-inflammatory action and possible relationship with overweight and obesity. | [26] |
| Sparc | Secreted by adipocytes, promoted adipose tissue fibrosis and inhibited adipogenesis. | Responsible for modulating the expression of pro-inflammatory cytokines that act on insulin resistance; and inhibits adipogenesis. | [23,27] |
| CTRPs | Expressed in conditions of adiponectin and leptin deficiency and high body mass index. | Regulation of inflammatory processes of adipose tissue. Regulation of glucose and fat metabolism in peripheral tissues and food intake. | [23,28] |
| FAM19A5 | Increase in fat mass. | Inhibits the proliferation and inflammation of vascular smooth muscle cell related to cardiovascular diseases through obesity. | [29] |
| WISP1 | Obesity, adipogenesis and visceral fat abnormalities. | Stimulates the cytokine response in macrophages associated with tissue adipose; induces the proliferation of mesenchymal stem cells, which increases tissue adipose. | [30] |
| Progranulin | Increase in fat mass associated with obesidade visceral, DM2 and dislipidemia. | It has anti-inflammatory properties. Hyper-progranulinemia is associated with insulin resistance and deficient insulin signaling. | [31,32] |
| Nesfatin-1 | Unclear | Induces satiety, which promotes body weight reduction. It can also regulate gastric distension and motility via the melanocortin pathway in the central nucleus of amygdala. | [23] |
| Visfatin | Increase in fat mass. | It produces adipocyte inflammation, insulin resistance and pancreatic beta cell dysfunction. | [33,34] |
| Fetuin-A | Increase in fat mass. | Associated to insulin resistance and inflammation. | [35] |
| ZAG | PPARγ, glucocorticoids, certain β3-adrenergic receptor agonists, thyroid hormones, and growth hormone (GH). | It acts in the acceleration of lipid metabolism, regulating enzymes of lipogenesis and lipolysis and stimulating production of adiponectin and BAT. | [36,37,38] |
| Myokine | Stimulation for Its Increase | Metabolic Action | Reference |
|---|---|---|---|
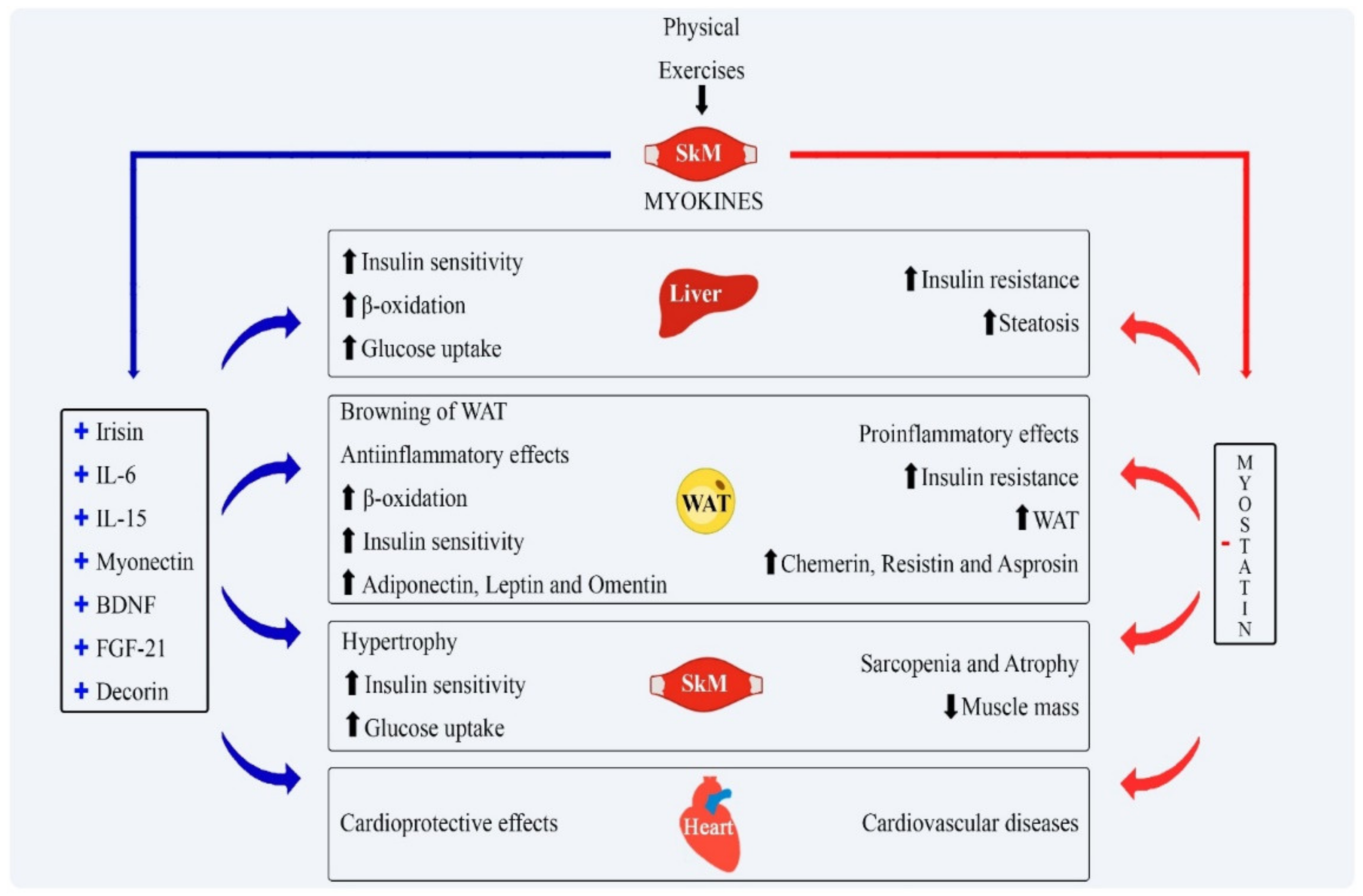
| Irisin | Physical exercise. | Darkening of WAT, increases energy expenditure, improves insulin sensitivity and induces weight loss. | [8] |
| BAIBA | Aerobic exercise | It acts in the Browning of adipose tissue, lipid oxidation and reduces insulin resistance. | [11] |
| Myostatin | Sedentary lifestyle | Induces muscle mass loss associated with insulin resistance and fat accumulation in the liver. Facilitates body fat accumulation. | [45] |
| Follistatin | Expressed in the context of physical activity, especially aerobic, resistance or high intensity training. | Inhibits the actions of myostatin, contributing to hypertrophy of skeletal muscle and reduction in fat mass, with consequent optimization of glucose uptake. | [13] |
| FGF21 | Physical exercise. | Increases insulin sensitivity, reduces plasma glucose and acts on lipolysis. | [57] |
| Apelin | Resistance exercises. | Anti-inflammatory role. It acts in the formation of new vessels and in the control of cardiac muscles and blood pressure. | [58] |
| Myonectin | Resistance exercises. | Increases the uptake of lipids by adipose tissue and liver, decreasing the plasma concentration of FFA. | [20] |
| IL-6 | Physical exercise. | Pro-inflammatory cytokine associated with insulin resistance in obesity. | [59] |
| IL-15 | Released after acute episodes of aerobic exercise. | Anti-inflammatory properties by inhibiting TNF-α expression; contributes to muscle hypertrophy, reduction in visceral adipose tissue and optimizes insulin action. | [14] |
| Sparc | Resistance exercises and muscle hypertrophy. | Inhibition of adipose tissue formation, increased insulin release and optimization of glucose uptake. | [60] |
| BDNF | Muscle and brain induced after exercise. | When produced by muscle, it increases sensitivity to insulin. | [61] |
| METRNL | Resistance exercises. | Anti-inflammatory role. Contributes to the browning of WAT and energy expenditure through the oxidation of glucose and FFA. | [62] |
| Decorin | Expressed in response to acute or chronic endurance training | Binds to myostatin by inhibiting its actions. As a consequence, it induces hypertrophy in the skeletal muscle. | [63] |
| Hepatokine | Stimulation for Its Increase | Metabolic Action | Reference |
|---|---|---|---|
| Fetuin A | Related to obesity, especially NAFLD and the increase in VAT. | It causes injury to the pancreas B cells and insulin resistance and works as a predictor of DM2. | [59,65] |
| Fetuin B | Increased in humans with steatosis and is related to insulin resistance. | Promotes insulin resistance and the development of diabetes. | [66] |
| Adropin | Regulated positively with food intake and weight reduction. | Stimulates lipolysis throughout the body, reducing weight gain and hepatic steatosis, optimizing the action of insulin and preventing the progression of atherosclerosis. | [67] |
| Activin E | High with obesity and NAFLD. | Reduces lipolysis and increase fat accumulation in adipocytes. | [68,69] |
| SHBG | Weight reduction and healthy lifestyle. | Transport of sex steroids to its target tissues. The increase in insulin sensitivity, stimulated by SHBG, is not yet fully cleared. | [70,71] |
| Chemerin | Produced in a state of obesity, dyslipidemia, metabolic syndrome and DM2. | Impairment of glucose homeostasis, increases insulin resistance and fat accumulation in the liver. | [14,72] |
| Selenoprotein | Associated with metabolic diseases, insulin resistance and hypoxia. | Attenuates fat loss induced by exercise. In hypoxia, insulin resistance and fat accumulation in adipose tissue increases. | [11,73] |
| Folistatin | It increases when the glucagon-to-insulin ratio rises in situations of aerobic exercise and resistance. | Actions on skeletal muscle hypertrophy, which increases glucose capture, and on the expression of thermogenic genes in murine adipocytes. | [11,74] |
| FGF21 | Aerobic exercises | It increases the sensitivity to insulin, the oxidation of fatty acids in the liver, decreases the production of glucose and the development of hepatic steatosis. | [11] |
| ANGPTL4 | Physical exercise | Stimulates lipolysis and decreases the action of the LPL enzyme on white adipose tissue. Inhibits pancreatic lipase and consequently decreases fat absorption. | [11,75] |
| ANGPTL4 | Food signals | Mediates food-driven resetting of circadian clock in mice liver; associated with regulation of inflammation, lipid metabolism, cancer cell invasion, and hematopoietic stem activity | [76] |
| LECT2 | Associated with metabolic stress | Impairment of insulin signal transduction and increases the appearance of pro-inflammatory cytokines. | [67] |
| Hepassocin | Elevated in pre-diabetes, DM2, and NAFLD. | Participates in the regulation of hepatocyte proliferation and liver regeneration. | [67,77] |
| Tsukushi | In response to NAFLD. | Reduces HDL-c cholesterol; reduced cholesterol efflux capacity, and reduces cholesterol-to–bile acid conversion in the liver. | [78] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira dos Santos, A.R.; de Oliveira Zanuso, B.; Miola, V.F.B.; Barbalho, S.M.; Santos Bueno, P.C.; Flato, U.A.P.; Detregiachi, C.R.P.; Buchaim, D.V.; Buchaim, R.L.; Tofano, R.J.; et al. Adipokines, Myokines, and Hepatokines: Crosstalk and Metabolic Repercussions. Int. J. Mol. Sci. 2021, 22, 2639. https://doi.org/10.3390/ijms22052639
de Oliveira dos Santos AR, de Oliveira Zanuso B, Miola VFB, Barbalho SM, Santos Bueno PC, Flato UAP, Detregiachi CRP, Buchaim DV, Buchaim RL, Tofano RJ, et al. Adipokines, Myokines, and Hepatokines: Crosstalk and Metabolic Repercussions. International Journal of Molecular Sciences. 2021; 22(5):2639. https://doi.org/10.3390/ijms22052639
Chicago/Turabian Stylede Oliveira dos Santos, Ana Rita, Bárbara de Oliveira Zanuso, Vitor Fernando Bordin Miola, Sandra Maria Barbalho, Patrícia C. Santos Bueno, Uri Adrian Prync Flato, Claudia Rucco P. Detregiachi, Daniela Vieira Buchaim, Rogério Leone Buchaim, Ricardo José Tofano, and et al. 2021. "Adipokines, Myokines, and Hepatokines: Crosstalk and Metabolic Repercussions" International Journal of Molecular Sciences 22, no. 5: 2639. https://doi.org/10.3390/ijms22052639
APA Stylede Oliveira dos Santos, A. R., de Oliveira Zanuso, B., Miola, V. F. B., Barbalho, S. M., Santos Bueno, P. C., Flato, U. A. P., Detregiachi, C. R. P., Buchaim, D. V., Buchaim, R. L., Tofano, R. J., Mendes, C. G., Tofano, V. A. C., & dos Santos Haber, J. F. (2021). Adipokines, Myokines, and Hepatokines: Crosstalk and Metabolic Repercussions. International Journal of Molecular Sciences, 22(5), 2639. https://doi.org/10.3390/ijms22052639