
microRNA-mRNA Profile of Skeletal Muscle Differentiation and Relevance to Congenital Myotonic Dystrophy

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Differential Expression of mRNA and miRNA during Myoblast Differentiation
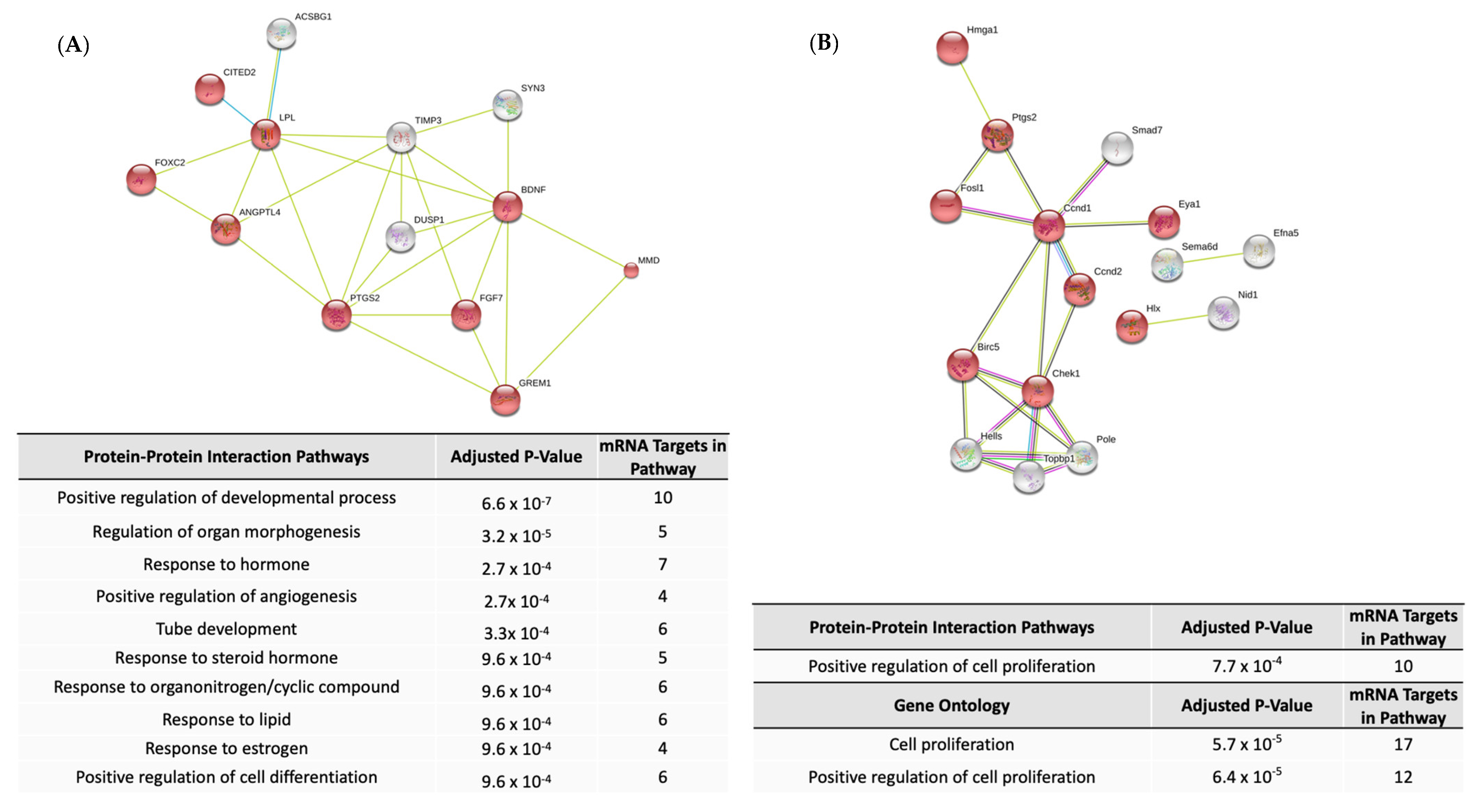
2.2. Pathway Enrichment for Differentially Expressed mRNAs
2.3. Pathway Enrichment for Predicted mRNA Targets of Differentially Expressed miRNAs
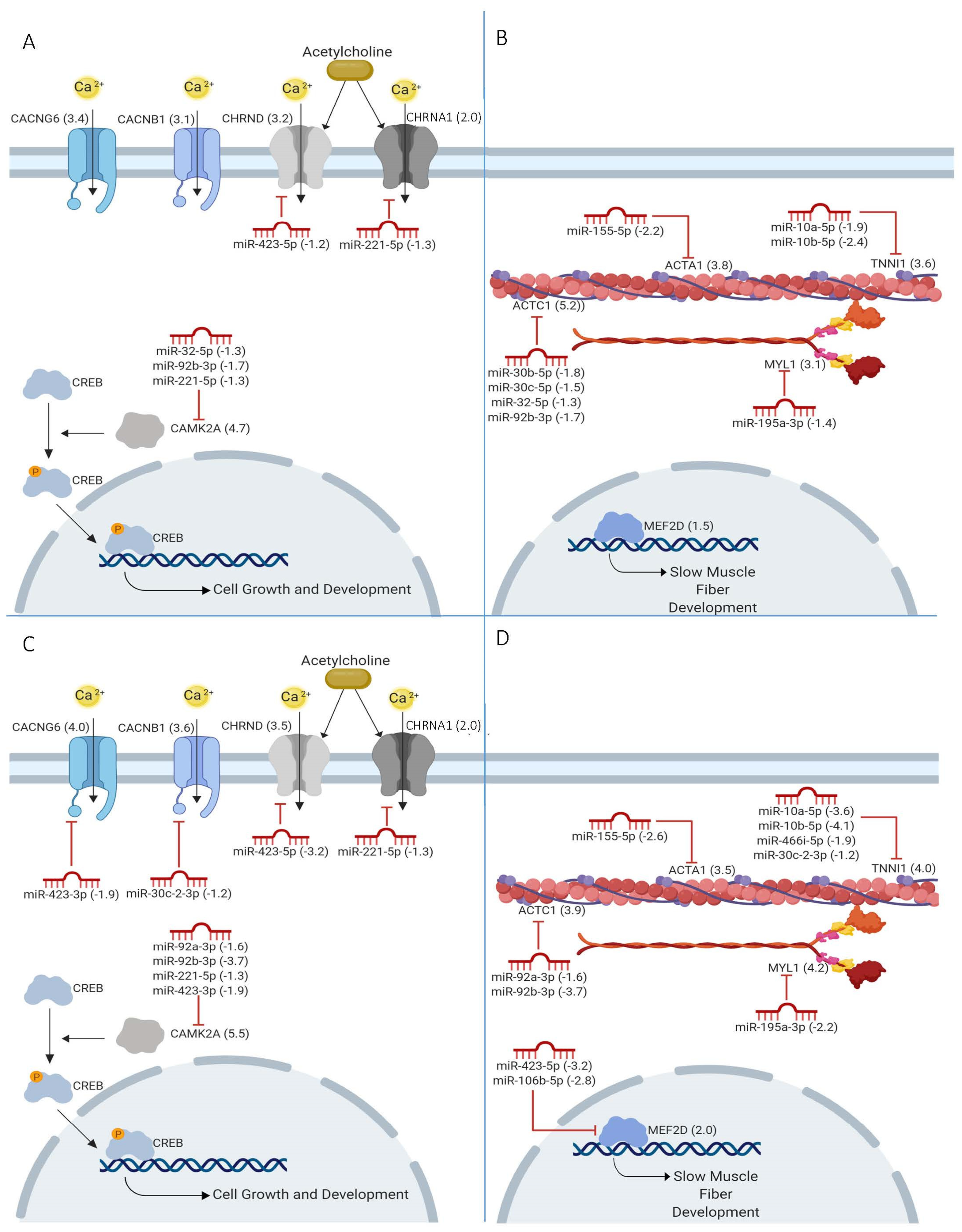
2.4. miRNAs in Calcium Signaling
2.5. Sarcomere Genes Targeted by miRNAs
2.6. myomiR Target Functions
2.7. Loss of miRNA Differential Expression in CMD
2.8. Chromosome X miRNAs in CMD
3. Discussion
3.1. Calcium Signaling and Sarcomere Function
3.2. miRNA Dysregulation in CMD
3.3. MyomiRs and Glucuronyltransferases
4. Materials and Methods
4.1. Cell Culture
4.2. RNA Extraction and Sequencing
4.3. mRNA Expression Analysis
4.4. miRNA-mRNA Correlation Analysis
4.5. Functional Enrichment
4.6. miRNA Expression during CMD Myoblast Differentiation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CMD | congenital myotonic dystrophy |
| mRNA | messenger RNA |
| miRNA | microRNA |
References
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [Green Version]
- Pasquinelli, A.E. MicroRNAs and their targets: Recognition, regulation and an emerging reciprocal relationship. Nat. Rev. Genet. 2012, 13, 271–282. [Google Scholar] [CrossRef]
- Kim, J.; Kang, Y.; Kojima, Y.; Lighthouse, J.K.; Hu, X.; Aldred, M.A.; McLean, D.L.; Park, H.; Comhair, S.A.; Greif, D.M.; et al. An endothelial apelin-FGF link mediated by miR-424 and miR-503 is disrupted in pulmonary arterial hypertension. Nat. Med. 2013, 19, 74–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Cook-Mills, J.; Morgan, G.; Sredni, S.T.; Pachman, L.M. Increased expression of vascular cell adhesion molecule 1 in muscle biopsy samples from juvenile dermatomyositis patients with short duration of untreated disease is regulated by miR-126. Arthritis Rheum. 2012, 64, 3809–3817. [Google Scholar] [CrossRef] [Green Version]
- Karolina, D.S.; Armugam, A.; Tavintharan, S.; Wong, M.T.K.; Lim, S.C.; Sum, C.F.; Jeyaseelan, K. MicroRNA 144 Impairs Insulin Signaling by Inhibiting the Expression of Insulin Receptor Substrate 1 in Type 2 Diabetes Mellitus. PLoS ONE 2011, 6, e22839. [Google Scholar] [CrossRef]
- Chen, J.-F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.-Z. The Role of MicroRNA-1 and MicroRNA-133 in Skeletal Muscle Proliferation and Differentiation. Nat. Genet. 2005, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ransom, J.F.; Li, A.; Vedantham, V.; Von Drehle, M.; Muth, A.N.; Tsuchihashi, T.; McManus, M.T.; Schwartz, R.J.; Srivastava, D. Dysregulation of Cardiogenesis, Cardiac Conduction, and Cell Cycle in Mice Lacking miRNA-1-2. Cell 2007, 129, 303–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horak, M.; Novak, J.; Bienertova-Vasku, J. Muscle-specific microRNAs in skeletal muscle development. Dev. Biol. 2016, 410, 1–13. [Google Scholar] [CrossRef]
- Liu, N.; Williams, A.H.; Maxeiner, J.M.; Bezprozvannaya, S.; Shelton, J.M.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. microRNA-206 promotes skeletal muscle regeneration and delays progression of Duchenne muscular dystrophy in mice. J. Clin. Investig. 2012, 122, 2054–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rooij, E.; Quiat, D.; Johnson, B.A.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Kelm, R.J.; Olson, E.N. A Family of microRNAs Encoded by Myosin Genes Governs Myosin Expression and Muscle Performance. Dev. Cell 2009, 17, 662–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, B.K.; Gagan, J.; Dutta, A. miR-206 and -486 Induce Myoblast Differentiation by Downregulating Pax7. Mol. Cell. Biol. 2010, 31, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Li, X.; Chen, C.; Li, Y.; Zhao, L.; Jing, Y.; Liu, W.; Wang, X.; Zhang, Y.; Xia, H.; et al. Attenuation of p38-Mediated miR-1/133 Expression Facilitates Myoblast Proliferation during the Early Stage of Muscle Regeneration. PLoS ONE 2012, 7, e41478. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-F.; Tao, Y.; Li, J.; Deng, Z.; Yan, Z.; Xiao, X.; Wang, D.-Z. microRNA-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7. J. Cell Biol. 2010, 190, 867–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Niu, L.-L.; Wei, W.; Zhang, W.-Y.; Li, X.-Y.; Cao, J.-H.; Zhao, S.-H. A feedback circuit between miR-133 and the ERK1/2 pathway involving an exquisite mechanism for regulating myoblast proliferation and differentiation. Cell Death Dis. 2013, 4, e934. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, I.; Eran, A.; Nishino, I.; Moggio, M.; Lamperti, C.; Amato, A.A.; Lidov, H.G.; Kang, P.B.; North, K.N.; Mitrani-Rosenbaum, S.; et al. Distinctive patterns of microRNA expression in primary muscular disorders. Proc. Natl. Acad. Sci. USA 2007, 104, 17016–17021. [Google Scholar] [CrossRef] [Green Version]
- Alexander, M.S.; Kawahara, G.; Motohashi, N.; Casar, J.C.; Eisenberg, I.; Myers, J.A.; Gasperini, M.J.; Estrella, E.A.; Kho, A.T.; Mitsuhashi, S.; et al. MicroRNA-199a is induced in dystrophic muscle and affects WNT signaling, cell proliferation, and myogenic differentiation. Cell Death Differ. 2013, 20, 1194–1208. [Google Scholar] [CrossRef]
- Perbellini, R.; Greco, S.; Sarra-Ferraris, G.; Cardani, R.; Capogrossi, M.C.; Meola, G.; Martelli, F. Dysregulation and cellular mislocalization of specific miRNAs in myotonic dystrophy type 1. Neuromuscul. Disord. 2011, 21, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Gambardella, S.; Rinaldi, F.; Lepore, S.M.; Viola, A.; Loro, E.; Angelini, C.; Vergani, L.; Novelli, G.; Botta, A. Overexpression of microRNA-206 in the skeletal muscle from myotonic dystrophy type 1 patients. J. Transl. Med. 2010, 8, 48. [Google Scholar] [CrossRef]
- Gaudet, P.; Livstone, M.S.; Lewis, S.E.; Thomas, P.D. Phylogenetic-based propagation of functional annotations within the Gene Ontology consortium. Briefings Bioinform. 2011, 12, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Lyukmanova, E.N.; Shenkarev, Z.O.; Shulepko, M.A.; Mineev, K.S.; D’Hoedt, D.; Kasheverov, I.E.; Filkin, S.Y.; Krivolapova, A.P.; Janickova, H.; Dolezal, V.; et al. NMR Structure and Action on Nicotinic Acetylcholine Receptors of Water-soluble Domain of Human LYNX1. J. Biol. Chem. 2011, 286, 10618–10627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, E.J.; Yau, K.S.; Ong, R.; Slee, J.; McGillivray, G.; Barnett, C.P.; Haliloglu, G.; Talim, B.; Akcoren, Z.; Kariminejad, A.; et al. Next generation sequencing in a large cohort of patients presenting with neuromuscular disease before or at birth. Orphanet J. Rare Dis. 2015, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cetin, H.; Epstein, M.; Liu, W.W.; Maxwell, S.; Cruz, P.M.R.; Cossins, J.; Vincent, A.; Webster, R.; Biggin, P.C.; Beeson, D. Muscle acetylcholine receptor conversion into chloride conductance at positive potentials by a single mutation. Proc. Natl. Acad. Sci. USA 2019, 116, 21228–21235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ow, J.R.; Palanichamy, M.K.; Rao, V.K.; Choi, M.H.; Bharathy, N.; Taneja, R. G9a inhibits {MEF2C} activity to control sarcomere assembly. Sci. Rep. 2016, 6, 34163. [Google Scholar] [CrossRef] [Green Version]
- Potthoff, M.J.; Olson, E.N. MEF2: A central regulator of diverse developmental programs. Development 2007, 134, 4131–4140. [Google Scholar] [CrossRef] [Green Version]
- Liang, T.; Zhou, B.; Shi, L.; Wang, H.; Chu, Q.; Xu, F.; Li, Y.; Chen, R.; Shen, C.; Schinckel, A.P. IncRNA AK017368 promotes proliferation and suppresses differentiation of myoblasts in skeletal muscle development by attenuating the function of miR-30c. FASEB J. 2018, 32, 377–389. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Li, G.; Yi, Z.; Nie, Q.; Zhang, X. E2F1-miR-20a-5p/20b-5p auto-regulatory feedback loop involved in myoblast proliferation and differentiation. Sci. Rep. 2016, 6, 27904. [Google Scholar] [CrossRef] [Green Version]
- Selbach, M.; Schwanhäusser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nat. Cell Biol. 2008, 455, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Penzkofer, D.; Bonauer, A.; Fischer, A.; Tups, A.; Brandes, R.P.; Zeiher, A.M.; Dimmeler, S. Phenotypic Characterization of miR-92a−/− Mice Reveals an Important Function of miR-92a in Skeletal Development. PLoS ONE 2014, 9, e101153. [Google Scholar] [CrossRef]
- Seok, H.Y.; Tatsuguchi, M.; Callis, T.E.; He, A.; Pu, W.T.; Wang, D.-Z. miR-155 Inhibits Expression of the MEF2A Protein to Repress Skeletal Muscle Differentiation. J. Biol. Chem. 2011, 286, 35339–35346. [Google Scholar] [CrossRef] [Green Version]
- Ge, G.; Yang, D.; Tan, Y.; Chen, Y.; Jiang, D.; Jiang, A.; Li, Q.; Liu, Y.; Zhong, Z.; Li, X.; et al. miR-10b-5p Regulates C2C12 Myoblasts Proliferation and Differentiation. Biosci. Biotechnol. Biochem. 2019, 83, 291–299. [Google Scholar] [CrossRef]
- Buckingham, M.; Bajard, L.; Chang, T.; Daubas, P.; Hadchouel, J.; Meilhac, S.; Montarras, D.; Rocancourt, D.; Relaix, F. The formation of skeletal muscle: From somite to limb. J. Anat. 2003, 202, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, N.J.; Tooney, P.A.; Carroll, A.P.; Tran, N.; Cairns, M.J. Down-regulation of miR-17 family expression in response to retinoic acid induced neuronal differentiation. Cell. Signal. 2009, 21, 1837–1845. [Google Scholar] [CrossRef] [PubMed]
- Ivanovska, I.; Ball, A.S.; Diaz, R.L.; Magnus, J.F.; Kibukawa, M.; Schelter, J.M.; Kobayashi, S.V.; Lim, L.; Burchard, J.; Jackson, A.L.; et al. MicroRNAs in the miR-106b Family Regulate p21/CDKN1A and Promote Cell Cycle Progression. Mol. Cell. Biol. 2008, 28, 2167–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrest, A.R.R.; Kanamori-Katayama, M.; Tomaru, Y.; Lassmann, T.; Ninomiya, N.; Takahashi, Y.; De Hoon, M.J.L.; Kubosaki, A.; Kaiho, A.; Suzuki, M.; et al. Induction of microRNAs, mir-155, mir-222, mir-424 and mir-503, promotes monocytic differentiation through combinatorial regulation. Leukemia 2009, 24, 460–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gramantieri, L.; Fornari, F.; Ferracin, M.; Veronese, A.; Sabbioni, S.; Calin, G.A.; Grazi, G.L.; Croce, C.M.; Bolondi, L.; Negrini, M. MicroRNA-221 Targets Bmf in Hepatocellular Carcinoma and Correlates with Tumor Multifocality. Clin. Cancer Res. 2009, 15, 5073–5081. [Google Scholar] [CrossRef] [Green Version]
- Potthoff, M.J.; Arnold, M.A.; McAnally, J.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. Regulation of Skeletal Muscle Sarcomere Integrity and Postnatal Muscle Function by Mef2c. Mol. Cell. Biol. 2007, 27, 8143–8151. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Lee, Y.S.; Sivaprasad, U.; Malhotra, A.; Dutta, A. Muscle-specific microRNA miR-206 promotes muscle differentiation. J. Cell Biol. 2006, 174, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Zhu, J.; Xia, B.; Cao, H.; Peng, Y.; Li, X.; Yu, T.; Chu, G.; Yang, G.; Shi, X. miR-423-5p inhibits myoblast proliferation and differentiation by targeting Sufu. J. Cell. Biochem. 2018, 119, 7610–7620. [Google Scholar] [CrossRef] [PubMed]
- Poon, E.N.-Y.; Hao, B.; Guan, D.; Li, M.J.; Lu, J.; Yang, Y.; Wu, B.; Wu, S.C.-M.; Webb, S.E.; Liang, Y.; et al. Integrated transcriptomic and regulatory network analyses identify microRNA-200c as a novel repressor of human pluripotent stem cell-derived cardiomyocyte differentiation and maturation. Cardiovasc. Res. 2018, 114, 894–906. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Liu, N.; Shao, Y.; Huang, Y.; Zhu, D. MiR-218 regulated cardiomyocyte differentiation and migration in mouse embryonic stem cells by targeting PDGFRα. J. Cell. Biochem. 2019, 120, 4355–4365. [Google Scholar] [CrossRef] [PubMed]
- Turczyńska, K.M.; Sadegh, M.K.; Hellstrand, P.; Swärd, K.; Albinsson, S. MicroRNAs Are Essential for Stretch-induced Vascular Smooth Muscle Contractile Differentiation via MicroRNA (miR)-145-dependent Expression of L-type Calcium Channels. J. Biol. Chem. 2012, 287, 19199–19206. [Google Scholar] [CrossRef] [Green Version]
- Harada, M.; Luo, X.; Murohara, T.; Yang, B.; Dobrev, D.; Nattel, S. MicroRNA Regulation and Cardiac Calcium Signaling. Circ. Res. 2014, 114, 689–705. [Google Scholar] [CrossRef] [Green Version]
- Tomczak, K.K.; Marinescu, V.D.; Ramoni, M.F.; Sanoudou, D.; Montanaro, F.; Han, M.; Kunkel, L.M.; Kohane, I.S.; Beggs, A.H. Expression profiling and identification of novel genes involved in myogenic differentiation. FASEB J. 2003, 18, 1–23. [Google Scholar] [CrossRef] [Green Version]
- McKinsey, T.A.; Zhang, C.-L.; Lu, J.; Olson, E.N. Signal-dependent nuclear export of a histone deacetylase regulates muscle differentiation. Nat. Cell Biol. 2000, 408, 106–111. [Google Scholar] [CrossRef]
- Kuklin, E.A.; Alkins, S.; Bakthavachalu, B.; Genco, M.C.; Sudhakaran, I.; Raghavan, K.V.; Ramaswami, M.; Griffith, L.C. The Long 3’UTR mRNA of CaMKII Is Essential for Translation-Dependent Plasticity of Spontaneous Release in Drosophila melanogaster. J. Neurosci. 2017, 37, 10554–10566. [Google Scholar] [CrossRef] [Green Version]
- Allen, D.L.; Leinwand, L.A. Intracellular Calcium and Myosin Isoform Transitions. J. Biol. Chem. 2002, 277, 45323–45330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, X.; Brown, L.D.; Liu, Y.; Schneider, M.F. Roles of the calcineurin and CaMK signaling pathways in fast-to-slow fiber type transformation of cultured adult mouse skeletal muscle fibers. Physiol. Genom. 2007, 30, 300–312. [Google Scholar] [CrossRef] [Green Version]
- Chin, E.R.; Olson, E.N.; Richardson, J.A.; Yang, Q.; Humphries, C.; Shelton, J.M.; Wu, H.; Zhu, W.; Bassel-Duby, R.; Williams, R.S. A calcineurin-dependent transcriptional pathway controls skeletal muscle fiber type. Genes Dev. 1998, 12, 2499–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, A.L.; Murgia, M.; Pallafacchina, G.; Calabria, E.; Coniglio, P.; Lømo, T.; Schiaffino, S. Calcineurin controls nerve activity-dependent specification of slow skeletal muscle fibers but not muscle growth. Proc. Natl. Acad. Sci. USA 2001, 98, 13108–13113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Roves, P.M.; Huss, J.; Holloszy, J.O. Role of calcineurin in exercise-induced mitochondrial biogenesis. Am. J. Physiol. Metab. 2006, 290, E1172–E1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.Q.; Garcia-Roves, P.M.; Barbosa, T.D.C.; Zierath, J.R. Constitutively active calcineurin in skeletal muscle increases endurance performance and mitochondrial respiratory capacity. Am. J. Physiol. Metab. 2010, 298, E8–E16. [Google Scholar] [CrossRef] [Green Version]
- Long, Y.C.; Glund, S.; Garcia-Roves, P.M.; Zierath, J.R. Calcineurin Regulates Skeletal Muscle Metabolism via Coordinated Changes in Gene Expression. J. Biol. Chem. 2007, 282, 1607–1614. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, C.; Xu, A.; Narayanan, N. Comparison of the effects of the membraneassociated Ca2+/calmodulin-dependent protein kinase on Ca2+-ATPase function in cardiac and slow-twitch skeletal muscle sarcoplasmic reticulum. Mol. Cell. Biochem. 1995, 142, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Aurora, A.B.; Mahmoud, A.I.; Luo, X.; Johnson, B.A.; Van Rooij, E.; Matsuzaki, S.; Humphries, K.M.; Hill, J.A.; Bassel-Duby, R.; Sadek, H.A.; et al. MicroRNA-214 protects the mouse heart from ischemic injury by controlling Ca2+ overload and cell death. J. Clin. Investig. 2012, 122, 1222–1232. [Google Scholar] [CrossRef]
- Wang, Q.; Quick, A.P.; Cao, S.; Reynolds, J.; Chiang, D.Y.; Beavers, D.; Li, N.; Wang, G.; Rodney, G.G.; Anderson, M.E.; et al. Oxidized CaMKII (Ca 2+ /Calmodulin-Dependent Protein Kinase II) Is Essential for Ventricular Arrhythmia in a Mouse Model of Duchenne Muscular Dystrophy. Circ. Arrhythm. Electrophysiol. 2018, 11, e005682. [Google Scholar] [CrossRef] [PubMed]
- Kramerova, I.; Ermolova, N.; Eskin, A.; Hevener, A.; Quehenberger, O.; Armando, A.M.; Haller, R.; Romain, N.; Nelson, S.F.; Spencer, M.J. Failure to up-regulate transcription of genes necessary for muscle adaptation underlies limb girdle muscular dystrophy 2A (calpainopathy). Hum. Mol. Genet. 2016, 25, 2194–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epellicena, P.; Eschulman, H. CaMKII inhibitors: From research tools to therapeutic agents. Front. Pharmacol. 2014, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, S.; Lee, M.; Akimoto, T. Conditional Deletion of Dicer in Adult Mice Impairs Skeletal Muscle Regeneration. Int. J. Mol. Sci. 2019, 20, 5686. [Google Scholar] [CrossRef] [Green Version]
- Rau, F.; Freyermuth, F.; Fugier, C.; Villemin, J.-P.; Fischer, M.-C.; Jost, B.; Dembele, D.; Gourdon, G.; Nicole, A.; Duboc, D.; et al. Misregulation of miR-1 processing is associated with heart defects in myotonic dystrophy. Nat. Struct. Mol. Biol. 2011, 18, 840–845. [Google Scholar] [CrossRef]
- Laurent, F.-X.; Sureau, A.; Klein, A.F.; Trouslard, F.; Gasnier, E.; Furling, D.; Marie, J. New function for the RNA helicase p68/DDX5 as a modifier of MBNL1 activity on expanded CUG repeats. Nucleic Acids Res. 2011, 40, 3159–3171. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.Y.; Barefield, D.Y.; Vo, A.H.; Gacita, A.M.; Schuster, E.J.; Wyatt, E.J.; Davis, J.L.; Dong, B.; Sun, C.; Page, P.; et al. Distinct pathological signatures in human cellular models of myotonic dystrophy subtypes. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Kaliman, P.; Llagostera, E. Myotonic dystrophy protein kinase (DMPK) and its role in the pathogenesis of myotonic dystrophy 1. Cell. Signal. 2008, 20, 1935–1941. [Google Scholar] [CrossRef]
- Klinck, R.; Fourrier, A.; Thibault, P.; Toutant, J.; Durand, M.; Lapointe, E.; Caillet-Boudin, M.-L.; Sergeant, N.; Gourdon, G.; Meola, G.; et al. RBFOX1 Cooperates with MBNL1 to Control Splicing in Muscle, Including Events Altered in Myotonic Dystrophy Type 1. PLoS ONE 2014, 9, e107324. [Google Scholar] [CrossRef] [Green Version]
- Kedde, M.; Van Kouwenhove, M.; Zwart, W.; Vrielink, J.A.F.O.; Elkon, R.; Agami, R. A Pumilio-induced RNA structure switch in p27-3′ UTR controls miR-221 and miR-222 accessibility. Nat. Cell Biol. 2010, 12, 1014–1020. [Google Scholar] [CrossRef]
- Cardinali, B.; Castellani, L.; Fasanaro, P.; Basso, A.; Alemà, S.; Martelli, F.; Falcone, G. Microrna-221 and Microrna-222 Modulate Differentiation and Maturation of Skeletal Muscle Cells. PLoS ONE 2009, 4, e7607. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Shi, Y.; He, H.; Cai, M.; Xiao, W.; Yang, X.; Chen, S.; Jia, X.; Wang, J.; Lai, S. miR-221 modulates skeletal muscle satellite cells proliferation and differentiation. Vitr. Cell. Dev. Biol. Anim. 2017, 54, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Dey, B.K.; Dutta, A. MiR-322/424 and -503 Are Induced during Muscle Differentiation and Promote Cell Cycle Quiescence and Differentiation by Down-Regulation of Cdc25A. Mol. Biol. Cell 2010, 21, 2138–2149. [Google Scholar] [CrossRef] [PubMed]
- Dogan, C.; De Antonio, M.; Hamroun, D.; Varet, H.; Fabbro, M.; Rougier, F.; Amarof, K.; Bes, M.-C.A.; Bedat-Millet, A.-L.; Behin, A.; et al. Gender as a Modifying Factor Influencing Myotonic Dystrophy Type 1 Phenotype Severity and Mortality: A Nationwide Multiple Databases Cross-Sectional Observational Study. PLoS ONE 2016, 11, e0148264. [Google Scholar] [CrossRef] [PubMed]
- Siissalo, S.; Zhang, H.; Stilgenbauer, E.; Kaukonen, A.M.; Hirvonen, J.; Finel, M. The Expression of Most UDP-Glucuronosyltransferases (UGTs) Is Increased Significantly during Caco-2 Cell Differentiation, whereas UGT1A6 Is Highly Expressed Also in Undifferentiated Cells. Drug Metab. Dispos. 2008, 36, 2331–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tukey, R.H.; Strassburg, C.P. Human UDP-Glucuronosyltransferases: Metabolism, Expression, and Disease. Annu. Rev. Pharmacol. Toxicol. 2000, 40, 581–616. [Google Scholar] [CrossRef] [PubMed]
- Pallafacchina, G.; François, S.; Regnault, B.; Czarny, B.; Dive, V.; Cumano, A.; Montarras, D.; Buckingham, M. An adult tissue-specific stem cell in its niche: A gene profiling analysis of in vivo quiescent and activated muscle satellite cells. Stem Cell Res. 2010, 4, 77–91. [Google Scholar] [CrossRef]
- Barresi, R. Dystroglycan: From biosynthesis to pathogenesis of human disease. J. Cell Sci. 2006, 119, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Inamori, K.-I.; Willer, T.; Hara, Y.; Venzke, D.; Anderson, M.E.; Clarke, N.F.; Guicheney, P.; Bönnemann, C.G.; Moore, S.A.; Campbell, K.P. Endogenous Glucuronyltransferase Activity of LARGE or LARGE2 Required for Functional Modification of α-Dystroglycan in Cells and Tissues. J. Biol. Chem. 2014, 289, 28138–28148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.M.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The biochemical basis of microRNA targeting efficacy. Science 2019, 366, eaav1741. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinform. 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.; Beffy, P.; Del Carratore, R.; Falleni, A.; Pretini, V.; D’Aurizio, R.; Botta, A.; Evangelista, M.; Stoccoro, A.; Coppedè, F.; et al. Activation of the interferon type I response rather than autophagy contributes to myogenesis inhibition in congenital DM1 myoblasts. Cell Death Dis. 2018, 9, 1071. [Google Scholar] [CrossRef] [PubMed]
- Beffy, P.; Del Carratore, R.; Masini, M.; Furling, D.; Puymirat, J.; Masiello, P.; Simili, M. Altered signal transduction pathways and induction of autophagy in human myotonic dystrophy type 1 myoblasts. Int. J. Biochem. Cell Biol. 2010, 42, 1973–1983. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | D2_D0 | D5_D0 | D5_D2 | |||
|---|---|---|---|---|---|---|
| p-Value | z-Score | p-Value | z-Score | p-Value | z-Score | |
| Calcium Signaling | 5.6 × 10−8 | 2.65 | 1.9 × 10−6 | 2.236 | - | - |
| Estrogen-mediated S-phase Entry | 1.3 × 10−5 | −1.00 | 3.7 × 10−4 | - | - | - |
| Nicotine Degradation | 2.3 × 10−5 | −1.34 | - | - | 2.6 × 10−8 | 2.00 |
| Melatonin Degradation | 3.2 × 10−5 | −1.34 | - | - | 3.4 × 10−8 | 2.00 |
| Cell Cycle: G1/S Checkpoint Regulation | 5.6 × 10−5 | 1.00 | 4.8 × 10−4 | - | - | - |
| Thyroid Hormone Metabolism | 8.3 × 10−5 | −2.00 | - | - | 5.2 × 10−9 | 2.00 |
| nNOS Signaling in Skeletal Muscle Cells | 1.1 × 10−4 | - | - | - | - | - |
| Cyclins and Cell Cycle Regulation | 1.4 × 10−4 | −1.34 | 9.8 × 10−4 | -1 | - | - |
| Clathrin-mediated Endocytosis Signaling | - | - | 9.1 × 10−5 | - | - | - |
| Actin Cytoskeleton Signaling | - | - | 2.0 × 10−4 | - | - | - |
| PPAR Signaling | - | - | 2.6 × 10−4 | −0.45 | - | - |
| Protein Kinase A Signaling | - | - | 3.2 × 10−4 | 1.63 | - | - |
| RhoA Signaling | - | - | 5.6 × 10−4 | 1.34 | - | |
| Serotonin Degradation | - | - | - | - | 5.4 × 10−8 | 2.00 |
| Xenobiotic Metabolism Signaling | - | - | - | - | 1.8 × 10−5 | - |
| Functional Category | p-Value | mRNAs Targets |
|---|---|---|
| Muscle structure development | 7.4 × 10−13 | 30 |
| Muscle system process | 1.3 × 10−12 | 25 |
| Circulatory system development | 3.2 × 10−11 | 35 |
| Muscle contraction | 6.7 × 10−11 | 21 |
| Anatomical structure formation involved in morphogenesis | 1.9 × 10−10 | 35 |
| Muscle organ development | 5.1 × 10−9 | 20 |
| Heart development | 5.8 × 10−9 | 23 |
| Striated muscle tissue development | 1.4 × 10−8 | 19 |
| Response to nitrogen compound | 1.9 × 10−8 | 31 |
| Cell differentiation | 1.9 × 10−8 | 68 |
| Cellular developmental process | 1.9 × 10−8 | 70 |
| Muscle tissue development | 1.9 × 10−8 | 19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morton, S.U.; Sefton, C.R.; Zhang, H.; Dai, M.; Turner, D.L.; Uhler, M.D.; Agrawal, P.B. microRNA-mRNA Profile of Skeletal Muscle Differentiation and Relevance to Congenital Myotonic Dystrophy. Int. J. Mol. Sci. 2021, 22, 2692. https://doi.org/10.3390/ijms22052692
Morton SU, Sefton CR, Zhang H, Dai M, Turner DL, Uhler MD, Agrawal PB. microRNA-mRNA Profile of Skeletal Muscle Differentiation and Relevance to Congenital Myotonic Dystrophy. International Journal of Molecular Sciences. 2021; 22(5):2692. https://doi.org/10.3390/ijms22052692
Chicago/Turabian StyleMorton, Sarah U., Christopher R. Sefton, Huanqing Zhang, Manhong Dai, David L. Turner, Michael D. Uhler, and Pankaj B. Agrawal. 2021. "microRNA-mRNA Profile of Skeletal Muscle Differentiation and Relevance to Congenital Myotonic Dystrophy" International Journal of Molecular Sciences 22, no. 5: 2692. https://doi.org/10.3390/ijms22052692