
Local Protein Translation and RNA Processing of Synaptic Proteins in Autism Spectrum Disorder


{kind=link}
Abstract
1. Introduction
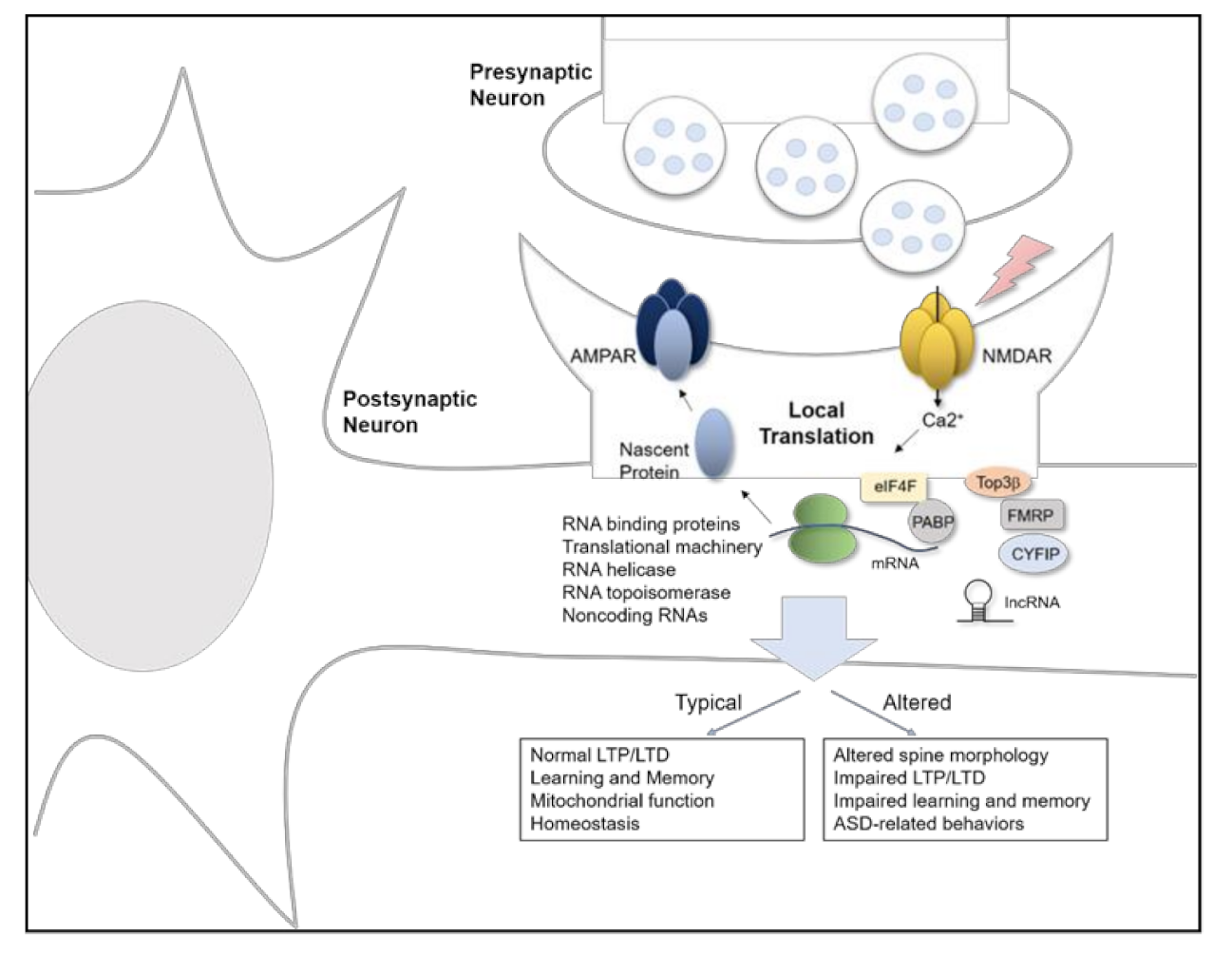
2. Local Translation of Synaptic Proteins in Synaptic Plasticity
3. Imaging Methods for Translation or Nascent Protein Research
4. Local Translation and ASD
4.1. Regulating RBPs and Translational Machineries in ASD
4.2. Mitochondrial RNA Translation in ASD
4.3. mRNA Modification in ASD
4.4. Noncoding RNA in Related to ASD
5. Potentials Links between RNA Metabolism, Protein Translation, and Immune Dysfunction in ASD
6. Potential RNA-Mediated Therapeutics in ASD
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Satterstrom, F.K.; Kosmicki, J.A.; Wang, J.; Breen, M.S.; de Rubeis, S.; An, J.Y.; Peng, M.; Collins, R.; Grove, J.; Klei, L.; et al. Large-Scale Exome Sequencing Study Implicates Both Developmental and Functional Changes in the Neurobiology of Autism. Cell 2020, 180, 568–584.e23. [Google Scholar] [CrossRef]
- Kelleher, R.J.; Bear, M.F. The Autistic Neuron: Troubled Translation? Cell 2008, 135, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Thelen, M.P.; Kye, M.J. The Role of Rna Binding Proteins for Local Mrna Translation: Implications in Neurological Disorders. Front. Mol. Biosci. 2019, 6, 161. [Google Scholar] [CrossRef]
- Livneh, I.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G.; Dominissini, D. The M 6 a Epitranscriptome: Transcriptome Plasticity in Brain Development and Function. Nat. Rev. Neurosci. 2020, 21, 36–51. [Google Scholar] [CrossRef]
- Gonatopoulos-Pournatzis, T.; Niibori, R.; Salter, E.W.; Weatheritt, R.J.; Tsang, B.; Farhangmehr, S.; Liang, X.; Braunschweig, U.; Roth, J.; Zhang, S.; et al. Autism-Misregulated Eif4g Microexons Control Synaptic Translation and Higher Order Cognitive Functions. Mol. Cell 2020, 77, 1176–1192.e16. [Google Scholar] [CrossRef] [PubMed]
- Liu-Yesucevitz, L.; Bassell, G.J.; Gitler, A.D.; Hart, A.C.; Klann, E.; Richter, J.D.; Warren, S.T.; Wolozin, B. Local Rna Translation at the Synapse and in Disease. J. Neurosci. 2011, 31, 16086–16093. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, X.; Ju, W.; Flory, M.; Zhong, J.; Jiang, S.; Wang, P.; Dong, X.; Tao, X.; Chen, Q.; et al. Genome-Wide Differential Expression of Synaptic Long Noncoding Rnas in Autism Spectrum Disorder. Transl. Psychiatry 2015, 5, e660. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Yoon, B.C.; Holt, C.E. Axonal Mrna Localization and Local Protein Synthesis in Nervous System Assembly, Maintenance and Repair. Nat. Rev. Neurosci. 2012, 13, 308–324. [Google Scholar] [CrossRef] [PubMed]
- Hafner, A.S.; Donlin-Asp, P.G.; Leitch, B.; Herzog, E.; Schuman, E.M. Local Protein Synthesis Is a Ubiquitous Feature of Neuronal Pre- and Postsynaptic Compartments. Science 2019, 364, eaau3644. [Google Scholar] [CrossRef]
- Younts, T.J.; Monday, H.R.; Dudok, B.; Klein, M.E.; Jordan, B.A.; Katona, I.; Castillo, P.E. Presynaptic Protein Synthesis Is Required for Long-Term Plasticity of Gaba Release. Neuron 2016, 92, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Mofatteh, M. Mrna Localization and Local Translation in Neurons. AIMS Neurosci. 2020, 7, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Rangaraju, V.; Dieck, S.T.; Schuman, E.M. Local Translation in Neuronal Compartments: How Local Is Local? EMBO Rep. 2017, 18, 693–711. [Google Scholar] [CrossRef]
- Martin, K.C.; Kosik, K.S. Synaptic Tagging—Who’s It? Nat. Rev. Neurosci. 2002, 3, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Simbriger, K.; Amorim, I.S.; Chalkiadaki, K.; Lach, G.; Jafarnejad, S.M.; Khoutorsky, A.; Gkogkas, C.G. Monitoring Translation in Synaptic Fractions Using a Ribosome Profiling Strategy. J. Neurosci. Methods 2020, 329, 108456. [Google Scholar] [CrossRef]
- Tang, A.H.; Chen, H.; Li, T.P.; Metzbower, S.R.; MacGillavry, H.D.; Blanpied, T.A. A Trans-Synaptic Nanocolumn Aligns Neurotransmitter Release to Receptors. Nature 2016, 536, 210–214. [Google Scholar] [CrossRef]
- Chen, H.; Tang, A.H.; Blanpied, T.A. Subsynaptic Spatial Organization as a Regulator of Synaptic Strength and Plasticity. Curr. Opin. Neurobiol. 2018, 51, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Fonkeu, Y.; Kraynyukova, N.; Hafner, A.S.; Kochen, L.; Sartori, F.; Schuman, E.M.; Tchumatchenko, T. How Mrna Localization and Protein Synthesis Sites Influence Dendritic Protein Distribution and Dynamics. Neuron 2019, 103, 1109–1122.e7. [Google Scholar] [CrossRef] [PubMed]
- Choquet, D. Linking Nanoscale Dynamics of Ampa Receptor Organization to Plasticity of Excitatory Synapses and Learning. J. Neurosci. 2018, 38, 9318–9329. [Google Scholar] [CrossRef]
- Crosby, K.C.; Gookin, S.E.; Garcia, J.D.; Hahm, K.M.; Dell’Acqua, M.L.; Smith, K.R. Nanoscale Subsynaptic Domains Underlie the Organization of the Inhibitory Synapse. Cell Rep. 2019, 26, 3284–3297.e3. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, J.; Bartol, T.M.; Camus, C.; Levet, F.; Menegolla, A.P.; Sejnowski, T.J.; Sibarita, J.B.; Vivaudou, M.; Choquet, D.; Hosy, E. Nanoscale Co-Organization and Coactivation of Ampar, Nmdar, and Mglur at Excitatory Synapses. Proc. Natl. Acad. Sci. USA 2020, 117, 14503–14511. [Google Scholar] [CrossRef]
- Ortiz, R.; Georgieva, M.V.; Gutiérrez, S.; Pedraza, N.; Fernández-Moya, S.M.; Gallego, C. Recruitment of Staufen2 Enhances Dendritic Localization of an Intron-Containing Camkiiα Mrna. Cell Rep. 2017, 20, 13–20. [Google Scholar] [CrossRef]
- Biever, A.; Glock, C.; Tushev, G.; Ciirdaeva, E.; Dalmay, T.; Langer, J.D.; Schuman, E.M. Monosomes Actively Translate Synaptic Mrnas in Neuronal Processes. Science 2020, 367, eaay4991. [Google Scholar] [CrossRef]
- Goodman, C.A.; Hornberger, T.A. Measuring Protein Synthesis with Sunset: A Valid Alternative to Traditional Techniques? Exerc. Sport Sci. Rev. 2013, 41, 107–115. [Google Scholar] [CrossRef]
- Dieck, T.S.; Kochen, L.; Hanus, C.; Heumüller, M.; Bartnik, I.; Nassim-Assir, B.; Merk, K.; Mosler, T.; Garg, S.; Bunse, S.; et al. Direct Visualization of Newly Synthesized Target Proteins in Situ. Nat. Methods 2015, 12, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Seo, J.Y.; Ryu, H.G.; Kim, D.Y.; Lee, K.H.; Kim, K.T. Bdnf-Induced Local Translation of glua1 is Regulated by Hnrnp A2/B1. Sci. Adv. 2020, 6, eabd2163. [Google Scholar] [CrossRef] [PubMed]
- Batista, A.F.R.; Martínez, J.C.; Hengst, U. Intra-Axonal Synthesis of Snap25 Is Required for the Formation of Presynaptic Terminals. Cell Rep. 2017, 20, 3085–3098. [Google Scholar] [CrossRef] [PubMed]
- Enam, S.U.; Zinshteyn, B.; Goldman, D.H.; Cassani, M.; Livingston, N.M.; Seydoux, G.; Green, R. Puromycin Reactivity Does Not Accurately Localize Translation at the Subcellular Level. eLife 2020, 9, e60303. [Google Scholar] [CrossRef] [PubMed]
- David, A.; Bennink, J.R.; Yewdell, J.W. Emetine Optimally Facilitates Nascent Chain Puromycylation and Potentiates the Ribopuromycylation Method (Rpm) Applied to Inert Cells. Histochem. Cell Biol. 2013, 139, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Tom Dieck, S.; Müller, A.; Nehring, A.; Hinz, F.I.; Bartnik, I.; Schuman, E.M.; Dieterich, D.C. Metabolic Labeling with Noncanonical Amino Acids and Visualization by Chemoselective Fluorescent Tagging. Curr. Protoc. Cell Biol. 2012, 56, 7.11.1–7.11.29. [Google Scholar]
- Alvarez-Castelao, B.; Schanzenbächer, C.T.; Langer, J.D.; Schuman, E.M. Cell-Type-Specific Metabolic Labeling, Detection and Identification of Nascent Proteomes in Vivo. Nat. Protoc. 2019, 14, 556–575. [Google Scholar] [CrossRef] [PubMed]
- Wade, O.K.; Woehrstein, J.B.; Nickels, P.C.; Strauss, S.; Stehr, F.; Stein, J.; Schueder, F.; Strauss, M.T.; Ganji, M.; Schnitzbauer, J.; et al. 124-Color Super-Resolution Imaging by Engineering DNA-Paint Blinking Kinetics. Nano Lett. 2019, 19, 2641–2646. [Google Scholar] [CrossRef] [PubMed]
- Tanenbaum, M.E.; Gilbert, L.A.; Qi, L.S.; Weissman, J.S.; Vale, R.D. A Protein-Tagging System for Signal Amplification in Gene Expression and Fluorescence Imaging. Cell 2014, 159, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Boersma, S.; Khuperkar, D.; Verhagen, B.M.P.; Sonneveld, S.; Grimm, J.B.; Lavis, L.D.; Tanenbaum, M.E. Multi-Color Single-Molecule Imaging Uncovers Extensive Heterogeneity in Mrna Decoding. Cell 2019, 178, 458–472.e19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Kamijo, K.; Fox, P.D.; Oda, H.; Morisaki, T.; Sato, Y.; Kimura, H.; Stasevich, T.J. A Genetically Encoded Probe for Imaging Nascent and Mature Ha-Tagged Proteins in Vivo. Nat. Commun. 2019, 10, 2947. [Google Scholar] [CrossRef] [PubMed]
- Pramparo, T.; Pierce, K.; Lombardo, M.V.; Barnes, C.C.; Marinero, S.; Ahrens-Barbeau, C.; Murray, S.S.; Lopez, L.; Xu, R.; Courchesne, E. Prediction of Autism by Translation and Immune/Inflammation Coexpressed Genes in Toddlers from Pediatric Community Practices. JAMA Psychiatry 2015, 72, 386–394. [Google Scholar] [CrossRef]
- Südhof, T.C. Neuroligins and Neurexins Link Synaptic Function to Cognitive Disease. Nature 2008, 455, 903–911. [Google Scholar] [CrossRef]
- Tabuchi, K.; Blundell, J.; Etherton, M.R.; Hammer, R.E.; Liu, X.; Powell, C.M.; Südhof, T.C. A Neuroligin-3 Mutation Implicated in Autism Increases Inhibitory Synaptic Transmission in Mice. Science 2007, 318, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Jamain, S.; Radyushkin, K.; Hammerschmidt, K.; Granon, S.; Boretius, S.; Varoqueaux, F.; Ramanantsoa, N.; Gallego, J.; Ronnenberg, A.; Winter, D.; et al. Reduced Social Interaction and Ultrasonic Communication in a Mouse Model of Monogenic Heritable Autism. Proc. Natl. Acad. Sci. USA 2008, 105, 1710–1715. [Google Scholar] [CrossRef]
- Monteiro, P.; Feng, G. Shank Proteins: Roles at the Synapse and in Autism Spectrum Disorder. Nat. Rev. Neurosci. 2017, 18, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Chmielewska, J.J.; Kuzniewska, B.; Milek, J.; Urbanska, K.; Dziembowska, M. Neuroligin 1, 2, and 3 Regulation at the Synapse: Fmrp-Dependent Translation and Activity-Induced Proteolytic Cleavage. Mol. Neurobiol. 2019, 56, 2741–2759. [Google Scholar] [CrossRef] [PubMed]
- Epstein, I.; Tushev, G.; Will, T.J.; Vlatkovic, I.; Cajigas, I.J.; Schuman, E.M. Alternative Polyadenylation and Differential Expression of Shank Mrnas in the Synaptic Neuropil. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130137. [Google Scholar] [CrossRef]
- Zalfa, F.; Giorgi, M.; Primerano, B.; Moro, A.; di Penta, A.; Reis, S.; Oostra, B.; Bagni, C. The Fragile X Syndrome Protein Fmrp Associates with Bc1 Rna and Regulates the Translation of Specific Mrnas at Synapses. Cell 2003, 112, 317–327. [Google Scholar] [CrossRef]
- Schenck, A.; Bardoni, B.; Moro, A.; Bagni, C.; Mandel, J.L. A Highly Conserved Protein Family Interacting with the Fragile X Mental Retardation Protein (Fmrp) and Displaying Selective Interactions with Fmrp-Related Proteins Fxr1p and Fxr2p. Proc. Natl. Acad. Sci. USA 2001, 98, 8844–8849. [Google Scholar] [CrossRef]
- Abekhoukh, S.; Bardoni, B. Cyfip Family Proteins between Autism and Intellectual Disability: Links with Fragile X Syndrome. Front. Cell. Neurosci. 2014, 8, 81. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Shen, W.; Guo, R.; Xue, Y.; Peng, W.; Sima, J.; Yang, J.; Sharov, A.; Srikantan, S.; Fox, D.; et al. Top3β Is an Rna Topoisomerase That Works with Fragile X Syndrome Protein to Promote Synapse Formation. Nat. Neurosci. 2013, 16, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, P.J. Topoisomerases and the Regulation of Neural Function. Nat. Rev. Neurosci. 2016, 17, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Joo, Y.; Xue, Y.; Wang, Y.; McDevitt, R.A.; Sah, N.; Bossi, S.; Su, S.; Lee, S.K.; Peng, W.; Xie, A.; et al. Topoisomerase 3β Knockout Mice Show Transcriptional and Behavioural Impairments Associated with Neurogenesis and Synaptic Plasticity. Nat. Commun. 2020, 11, 3143. [Google Scholar] [CrossRef]
- Crunkhorn, S. Neurological Disorders: Targeting Translation in Autism. Nat. Rev. Drug Discov. 2013, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Baj, G.; Pinhero, V.; Vaghi, V.; Tongiorgi, E. Signaling Pathways Controlling Activity-Dependent Local Translation of Bdnf and Their Localization in Dendritic Arbors. J. Cell Sci. 2016, 129, 2852–2864. [Google Scholar] [CrossRef]
- Sawazaki, R.; Imai, S.; Yokogawa, M.; Hosoda, N.; Hoshino, S.I.; Mio, M.; Mio, K.; Shimada, I.; Osawa, M. Characterization of the Multimeric Structure of Poly(a)-Binding Protein on a Poly(a) Tail. Sci. Rep. 2018, 8, 1455. [Google Scholar] [CrossRef] [PubMed]
- Eura, N.; Sugie, K.; Suzuki, N.; Kiriyama, T.; Izumi, T.; Shimakura, N.; Kato, M.; Aoki, M. A Juvenile Sporadic Amyotrophic Lateral Sclerosis Case with P525l Mutation in the Fus Gene: A Rare Co-Occurrence of Autism Spectrum Disorder and Tremor. J. Neurol. Sci. 2019, 398, 67–68. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.Y.; Chang, J.C.; Tyan, S.H.; Yen, Y.C.; Lim, K.; Tan, B.S.Y.; Ong, J.; Tucker-Kellogg, G.; Wong, P.; Koo, E.; et al. Fus-Mediated Dysregulation of Sema5a, an Autism-Related Gene, in Fus Mice with Hippocampus-Dependent Cognitive Deficits. Hum. Mol. Genet. 2019, 28, 3777–3791. [Google Scholar] [CrossRef]
- Berg, J.M.; Lee, C.; Chen, L.; Galvan, L.; Cepeda, C.; Chen, J.Y.; Peñagarikano, O.; Stein, J.L.; Li, A.; Oguro-Ando, A.; et al. Jakmip1, a Novel Regulator of Neuronal Translation, Modulates Synaptic Function and Autistic-Like Behaviors in Mouse. Neuron 2015, 88, 1173–1191. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.F.; Elwell, C.; Johnson, M.H. Mitochondrial Dysfunction in Autism Spectrum Disorders. Autism Open Access 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Citrigno, L.; Muglia, M.; Qualtieri, A.; Spadafora, P.; Cavalcanti, F.; Pioggia, G.; Cerasa, A. The Mitochondrial Dysfunction Hypothesis in Autism Spectrum Disorders: Current Status and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 5785. [Google Scholar] [CrossRef] [PubMed]
- Kuzniewska, B.; Cysewski, D.; Wasilewski, M.; Sakowska, P.; Milek, J.; Kulinski, T.M.; Winiarski, M.; Kozielewicz, P.; Knapska, E.; Dadlez, M.; et al. Mitochondrial Protein Biogenesis in the Synapse Is Supported by Local Translation. EMBO Rep. 2020, 21, e48882. [Google Scholar] [CrossRef]
- Si, K.; Giustetto, M.; Etkin, A.; Hsu, R.; Janisiewicz, A.M.; Miniaci, M.C.; Kim, J.H.; Zhu, H.; Kandel, E.R. A Neuronal Isoform of Cpeb Regulates Local Protein Synthesis and Stabilizes Synapse-Specific Long-Term Facilitation in Aplysia. Cell 2003, 115, 893–904. [Google Scholar] [CrossRef]
- Sinnamon, J.R.; Czaplinski, K. Mrna Trafficking and Local Translation: The Yin and Yang of Regulating Mrna Localization in Neurons. Acta Biochim. Biophys. Sin. 2011, 43, 663–670. [Google Scholar] [CrossRef][Green Version]
- Oruganty-Das, A.; Ng, T.; Udagawa, T.; Goh, E.L.; Richter, J.D. Translational Control of Mitochondrial Energy Production Mediates Neuron Morphogenesis. Cell Metab. 2012, 16, 789–800. [Google Scholar] [CrossRef]
- Anitha, A.; Nakamura, K.; Thanseem, I.; Yamada, K.; Iwayama, Y.; Toyota, T.; Matsuzaki, H.; Miyachi, T.; Yamada, S.; Tsujii, M.; et al. Brain Region-Specific Altered Expression and Association of Mitochondria-Related Genes in Autism. Mol. Autism 2012, 3, 12. [Google Scholar] [CrossRef]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic Rna Modifications in Gene Expression Regulation. Cell 2017, 169, 1187–1200. [Google Scholar] [CrossRef]
- Marcotrigiano, J.; Gingras, A.C.; Sonenberg, N.; Burley, S.K. X-ray Studies of the Messenger Rna 5′ Cap-Binding Protein (Eif4e) Bound to 7-Methyl-Gdp. Nucleic Acids Symp. Ser. 1997, 36, 8–11. [Google Scholar]
- Sonenberg, N.; Gingras, A.C. The Mrna 5′ Cap-Binding Protein Eif4e and Control of Cell Growth. Curr. Opin. Cell Biol. 1998, 10, 268–275. [Google Scholar] [CrossRef]
- Gkogkas, C.G.; Khoutorsky, A.; Ran, I.; Rampakakis, E.; Nevarko, T.; Weatherill, D.B.; Vasuta, C.; Yee, S.; Truitt, M.; Dallaire, P.; et al. Autism-Related Deficits Via Dysregulated Eif4e-Dependent Translational Control. Nature 2013, 493, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Santini, E.; Huynh, T.N.; MacAskill, A.F.; Carter, A.G.; Pierre, P.; Ruggero, D.; Kaphzan, H.; Klann, E. Exaggerated Translation Causes Synaptic and Behavioural Aberrations Associated with Autism. Nature 2013, 493, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Patil, D.P.; Zhou, J.; Zinoviev, A.; Skabkin, M.A.; Elemento, O.; Pestova, T.V.; Qian, S.B.; Jaffrey, S.R. 5′ Utr M(6)a Promotes Cap-Independent Translation. Cell 2015, 163, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-Methyladenosine in Nuclear Rna Is a Major Substrate of the Obesity-Associated Fto. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vågbø, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. Alkbh5 Is a Mammalian Rna Demethylase That Impacts Rna Metabolism and Mouse Fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef]
- Yang, C.; Hu, Y.; Zhou, B.; Bao, Y.; Li, Z.; Gong, C.; Yang, H.; Wang, S.; Xiao, Y. The Role of M 6 a Modification in Physiology and Disease. Cell Death Dis. 2020, 11, 960. [Google Scholar] [CrossRef]
- Yoon, K.J.; Ringeling, F.R.; Vissers, C.; Jacob, F.; Pokrass, M.; Jimenez-Cyrus, D.; Su, Y.; Kim, N.S.; Zhu, Y.; Zheng, L.; et al. Temporal Control of Mammalian Cortical Neurogenesis by M 6 a Methylation. Cell 2017, 171, 877–889.e17. [Google Scholar] [CrossRef] [PubMed]
- Edens, B.M.; Vissers, C.; Su, J.; Arumugam, S.; Xu, Z.; Shi, H.; Miller, N.; Ringeling, F.R.; Ming, G.L.; He, C.; et al. Fmrp Modulates Neural Differentiation through M 6 a-Dependent Mrna Nuclear Export. Cell Rep. 2019, 28, 845–854.e5. [Google Scholar] [CrossRef]
- Walters, B.J.; Mercaldo, V.; Gillon, C.J.; Yip, M.; Neve, R.L.; Boyce, F.M.; Frankland, P.W.; Josselyn, S.A. The Role of the Rna Demethylase Fto (Fat Mass and Obesity-Associated) and Mrna Methylation in Hippocampal Memory Formation. Neuropsychopharmacology 2017, 42, 1502–1510. [Google Scholar] [CrossRef] [PubMed]
- Widagdo, J.; Zhao, Q.Y.; Kempen, M.J.; Tan, M.C.; Ratnu, V.S.; Wei, W.; Leighton, L.; Spadaro, P.A.; Edson, J.; Anggono, V.; et al. Experience-Dependent Accumulation of N6-Methyladenosine in the Prefrontal Cortex Is Associated with Memory Processes in Mice. J. Neurosci. 2016, 36, 6771–6777. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yu, C.; Guo, M.; Zheng, X.; Ali, S.; Huang, H.; Zhang, L.; Wang, S.; Huang, Y.; Qie, S.; et al. Down-Regulation of M6a Mrna Methylation Is Involved in Dopaminergic Neuronal Death. ACS Chem. Neurosci. 2019, 10, 2355–2363. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.; Sun, B.F.; Chen, Y.S.; Xu, J.W.; Lai, W.Y.; Li, A.; Wang, X.; Bhattarai, D.P.; Xiao, W.; et al. 5-Methylcytosine Promotes Mrna Export-Nsun2 as the Methyltransferase and Alyref as An m 5 C Reader. Cell Res. 2017, 27, 606–625. [Google Scholar] [CrossRef] [PubMed]
- Svitkin, Y.V.; Cheng, Y.M.; Chakraborty, T.; Presnyak, V.; John, M.; Sonenberg, N. N1-Methyl-Pseudouridine in Mrna Enhances Translation through Eif2α-Dependent and Independent Mechanisms by Increasing Ribosome Density. Nucleic Acids Res. 2017, 45, 6023–6036. [Google Scholar] [CrossRef]
- Eyler, D.E.; Franco, M.K.; Batool, Z.; Wu, M.Z.; Dubuke, M.L.; Dobosz-Bartoszek, M.; Jones, J.D.; Polikanov, Y.S.; Roy, B.; Koutmou, K.S. Pseudouridinylation of Mrna Coding Sequences Alters Translation. Proc. Natl. Acad. Sci. USA 2019, 116, 23068–23074. [Google Scholar] [CrossRef]
- Darvish, H.; Azcona, L.J.; Alehabib, E.; Jamali, F.; Tafakhori, A.; Ranji-Burachaloo, S.; Jen, J.C.; Paisán-Ruiz, C. A Novel pus7 mutation Causes Intellectual Disability with Autistic and Aggressive Behaviors. Neurol. Genet. 2019, 5, e356. [Google Scholar] [CrossRef] [PubMed]
- Lipovich, L.; Tarca, A.L.; Cai, J.; Jia, H.; Chugani, H.T.; Sterner, K.N.; Grossman, L.I.; Uddin, M.; Hof, P.R.; Sherwood, C.C.; et al. Developmental Changes in the Transcriptome of Human Cerebral Cortex Tissue: Long Noncoding Rna Transcripts. Cereb. Cortex 2014, 24, 1451–1459. [Google Scholar] [CrossRef] [PubMed]
- Ladd, P.D.; Smith, L.E.; Rabaia, N.A.; Moore, J.M.; Georges, S.A.; Hansen, R.S.; Hagerman, R.J.; Tassone, F.; Tapscott, S.J.; Filippova, G.N. An Antisense Transcript Spanning the Cgg Repeat Region of Fmr1 Is Upregulated in Premutation Carriers but Silenced in Full Mutation Individuals. Hum. Mol. Genet. 2007, 16, 3174–3187. [Google Scholar] [CrossRef] [PubMed]
- Peschansky, V.J.; Pastori, C.; Zeier, Z.; Wentzel, K.; Velmeshev, D.; Magistri, M.; Silva, J.P.; Wahlestedt, C. The Long Non-Coding Rna Fmr4 Promotes Proliferation of Human Neural Precursor Cells and Epigenetic Regulation of Gene Expression in Trans. Mol. Cell. Neurosci. 2016, 74, 49–57. [Google Scholar] [CrossRef]
- Centonze, D.; Rossi, S.; Napoli, I.; Mercaldo, V.; Lacoux, C.; Ferrari, F.; Ciotti, M.T.; de Chiara, V.; Prosperetti, C.; Maccarrone, M.; et al. The Brain Cytoplasmic Rna Bc1 Regulates Dopamine D2 Receptor-Mediated Transmission in the Striatum. J. Neurosci. 2007, 27, 8885–8892. [Google Scholar] [CrossRef]
- Briz, V.; Restivo, L.; Pasciuto, E.; Juczewski, K.; Mercaldo, V.; Lo, A.C.; Baatsen, P.; Gounko, N.V.; Borreca, A.; Girardi, T.; et al. The Non-Coding Rna Bc1 Regulates Experience-Dependent Structural Plasticity and Learning. Nat. Commun. 2017, 8, 293. [Google Scholar] [CrossRef]
- Pruunsild, P.; Kazantseva, A.; Aid, T.; Palm, K.; Timmusk, T. Dissecting the Human Bdnf Locus: Bidirectional Transcription, Complex Splicing, and Multiple Promoters. Genomics 2007, 90, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Bernard, D.; Prasanth, K.V.; Tripathi, V.; Colasse, S.; Nakamura, T.; Xuan, Z.; Zhang, M.Q.; Sedel, F.; Jourdren, L.; Coulpier, F.; et al. A Long Nuclear-Retained Non-Coding Rna Regulates Synaptogenesis by Modulating Gene Expression. EMBO J. 2010, 29, 3082–3093. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Shen, Y.; Si, Y.; Zhang, Y.; Du, J.; Hu, X.; Cai, M.; Bao, H.; Xing, Y. Knockdown of Long Non-Coding Rna Sox2ot Downregulates Sox2 to Improve Hippocampal Neurogenesis and Cognitive Function in a Mouse Model of Sepsis-Associated Encephalopathy. J. Neuroinflamm. 2020, 17, 320. [Google Scholar] [CrossRef] [PubMed]
- Ozkul, Y.; Taheri, S.; Bayram, K.K.; Sener, E.F.; Mehmetbeyoglu, E.; Öztop, D.B.; Aybuga, F.; Tufan, E.; Bayram, A.; Dolu, N.; et al. A Heritable Profile of Six Mirnas in Autistic Patients and Mouse Models. Sci. Rep. 2020, 10, 9011. [Google Scholar] [CrossRef] [PubMed]
- Edbauer, D.; Neilson, J.R.; Foster, K.A.; Wang, C.F.; Seeburg, D.P.; Batterton, M.N.; Tada, T.; Dolan, B.M.; Sharp, P.A.; Sheng, M. Regulation of Synaptic Structure and Function by Fmrp-Associated Micrornas Mir-125b and Mir-132. Neuron 2010, 65, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Zongaro, S.; Hukema, R.; D’Antoni, S.; Davidovic, L.; Barbry, P.; Catania, M.V.; Willemsen, R.; Mari, B.; Bardoni, B. The 3′ Utr of Fmr1 Mrna Is a Target of Mir-101, Mir-129-5p and Mir-221: Implications for the Molecular Pathology of Fxtas at the Synapse. Hum. Mol. Genet. 2013, 22, 1971–1982. [Google Scholar] [CrossRef]
- Sakaguchi, A.; Yamashita, Y.; Ishii, T.; Uehara, T.; Kosaki, K.; Takahashi, T.; Takenouchi, T. Further Evidence of a Causal Association between Ago1, a Critical Regulator of Microrna Formation, and Intellectual Disability/Autism Spectrum Disorder. Eur. J. Med. Genet. 2019, 62, 103537. [Google Scholar] [CrossRef]
- Meister, G. Argonaute Proteins: Functional Insights and Emerging Roles. Nat. Rev. Genet. 2013, 14, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular Rnas. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Chen, C.Y.; Mai, T.L.; Chuang, C.F.; Chen, Y.C.; Gupta, S.K.; Yen, L.; Wang, Y.D.; Chuang, T.J. Genome-Wide, Integrative Analysis of Circular Rna Dysregulation and the Corresponding Circular Rna-Microrna-Mrna Regulatory Axes in Autism. Genome Res. 2020, 30, 375–391. [Google Scholar] [CrossRef] [PubMed]
- Gokool, A.; Loy, C.T.; Halliday, G.M.; Voineagu, I. Circular Rnas: The Brain Transcriptome Comes Full Circle. Trends Neurosci. 2020, 43, 752–766. [Google Scholar] [CrossRef]
- Gasparini, S.; Licursi, V.; Presutti, C.; Mannironi, C. The Secret Garden of Neuronal Circrnas. Cells 2020, 9, 1815. [Google Scholar] [CrossRef]
- Montani, C.; Gritti, L.; Beretta, S.; Verpelli, C.; Sala, C. The Synaptic and Neuronal Functions of the X-Linked Intellectual Disability Protein Interleukin-1 Receptor Accessory Protein Like 1 (Il1rapl1). Dev. Neurobiol. 2019, 79, 85–95. [Google Scholar]
- Yuan, N.; Chen, Y.; Xia, Y.; Dai, J.; Liu, C. Inflammation-Related Biomarkers in Major Psychiatric Disorders: A Cross-Disorder Assessment of Reproducibility and Specificity in 43 Meta-Analyses. Transl. Psychiatry 2019, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Murdock, M.H.; Jing, D.; Won, T.H.; Chung, H.; Kressel, A.M.; Tsaava, T.; Addorisio, M.E.; Putzel, G.G.; Zhou, L.; et al. The Microbiota Regulate Neuronal Function and Fear Extinction Learning. Nature 2019, 574, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Eshraghi, R.S.; Davies, C.; Iyengar, R.; Perez, L.; Mittal, R.; Eshraghi, A.A. Gut-Induced Inflammation During Development May Compromise the Blood-Brain Barrier and Predispose to Autism Spectrum Disorder. J. Clin. Med. 2020, 10, 27. [Google Scholar] [CrossRef]
- Moradi, K.; Ashraf-Ganjouei, A.; Tavolinejad, H.; Bagheri, S.; Akhondzadeh, S. The Interplay between Gut Microbiota and Autism Spectrum Disorders: A Focus on Immunological Pathways. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 106, 110091. [Google Scholar] [CrossRef]
- Rinaldi, C.; Wood, M.J.A. Antisense Oligonucleotides: The Next Frontier for Treatment of Neurological Disorders. Nat. Rev. Neurol. 2018, 14, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Scoles, D.R.; Minikel, E.V.; Pulst, S.M. Antisense Oligonucleotides: A Primer. Neurol. Genet. 2019, 5, e323. [Google Scholar] [CrossRef]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in Oligonucleotide Drug Delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Dammes, N.; Peer, D. Paving the Road for Rna Therapeutics. Trends Pharmacol. Sci. 2020, 41, 755–775. [Google Scholar] [CrossRef] [PubMed]
- Ruan, G.X.; Barry, E.; Yu, D.; Lukason, M.; Cheng, S.H.; Scaria, A. Crispr/Cas9-Mediated Genome Editing as a Therapeutic Approach for Leber Congenital Amaurosis 10. Mol. Ther. 2017, 25, 331–341. [Google Scholar] [CrossRef]
- Xie, N.; Gong, H.; Suhl, J.A.; Chopra, P.; Wang, T.; Warren, S.T. Reactivation of Fmr1 by Crispr/Cas9-Mediated Deletion of the Expanded Cgg-Repeat of the Fragile X Chromosome. PLoS ONE 2016, 10, e0165499. [Google Scholar] [CrossRef] [PubMed]
- Hicks, S.D.; Rajan, A.T.; Wagner, K.E.; Barns, S.; Carpenter, R.L.; Middleton, F.A. Validation of a Salivary Rna Test for Childhood Autism Spectrum Disorder. Front. Genet. 2018, 9, 534. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joo, Y.; Benavides, D.R. Local Protein Translation and RNA Processing of Synaptic Proteins in Autism Spectrum Disorder. Int. J. Mol. Sci. 2021, 22, 2811. https://doi.org/10.3390/ijms22062811
Joo Y, Benavides DR. Local Protein Translation and RNA Processing of Synaptic Proteins in Autism Spectrum Disorder. International Journal of Molecular Sciences. 2021; 22(6):2811. https://doi.org/10.3390/ijms22062811
Chicago/Turabian StyleJoo, Yuyoung, and David R. Benavides. 2021. "Local Protein Translation and RNA Processing of Synaptic Proteins in Autism Spectrum Disorder" International Journal of Molecular Sciences 22, no. 6: 2811. https://doi.org/10.3390/ijms22062811
APA StyleJoo, Y., & Benavides, D. R. (2021). Local Protein Translation and RNA Processing of Synaptic Proteins in Autism Spectrum Disorder. International Journal of Molecular Sciences, 22(6), 2811. https://doi.org/10.3390/ijms22062811