
The Effects of Cold Atmospheric Pressure Plasma on Germination Parameters, Enzyme Activities and Induction of DNA Damage in Barley

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Germination Parameters

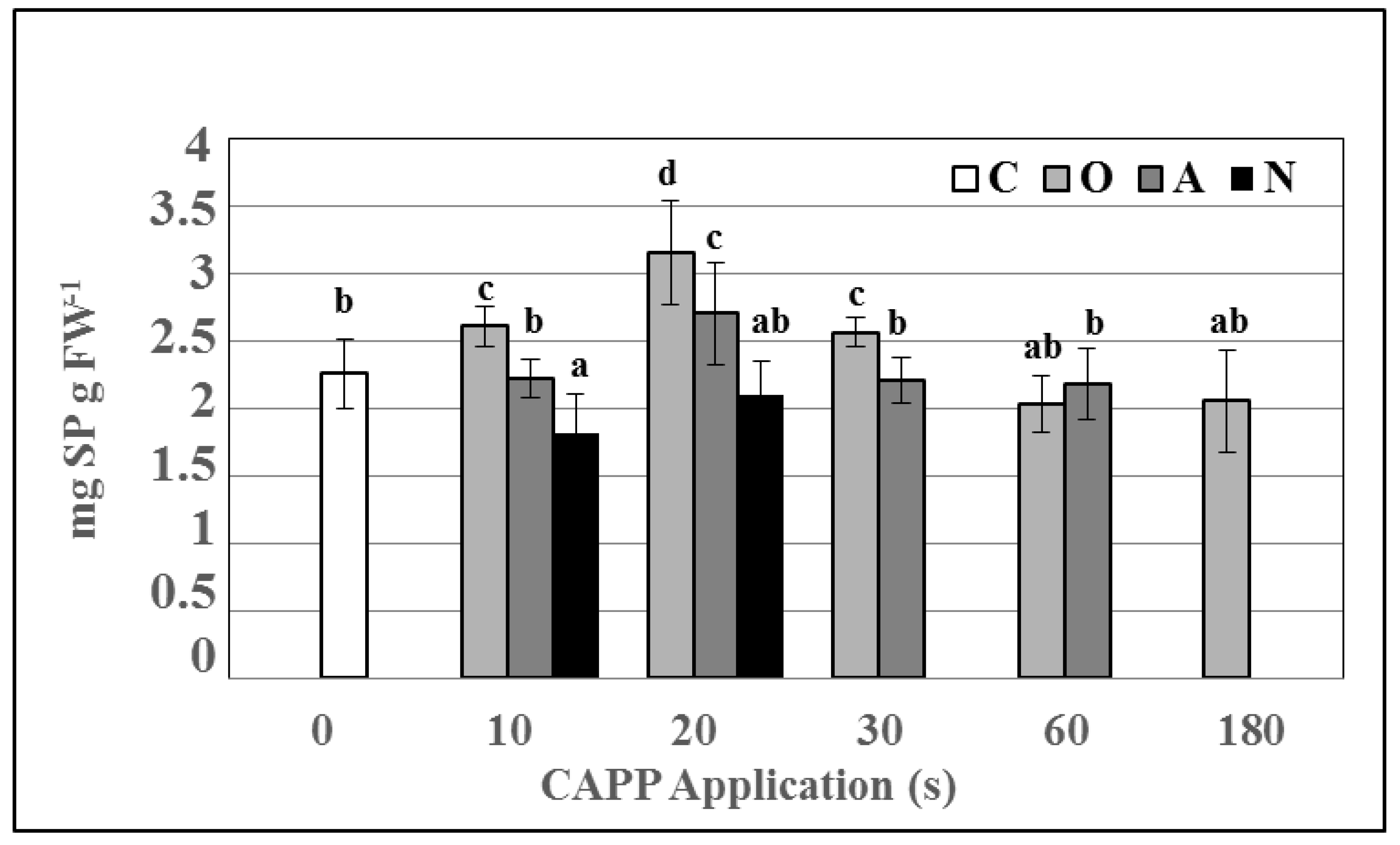
2.2. Enzyme Activities
2.3. Genotoxic Effects
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Plasma Generation and Samples Treatment
4.3. Germination and Growth Conditions
4.4. Total Soluble Proteins Content
4.5. Protease and Glucanase Activity Assays
4.6. Assays of Guaiacol Peroxidase (POX, E.C.1.11.1.7) and Superoxide Dismutase (SOD, E.C.1.15.1.1) Activities
4.7. Alkaline Comet Assay
4.8. Constant Field Gel Electrophoresis
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ekanayake, U.M.; Seo, D.H.; Faershteyn, K.; O’Mullane, A.P.; Shon, H.; MacLeod, J.; Golberg, D.; Ostrikov, K. Atmospheric-pressure plasma seawater desalination: Clean energy, agriculture, and resource recovery nexus for a blue planet. Sustain. Mater. Technol. 2020, 25, e00181. [Google Scholar] [CrossRef]
- Lee, M.; Fan, C.-S.; Chen, Y.-W.; Chang, K.-C.; Chiueh, P.-T.; Hou, C.-H. Membrane capacitive deionization for low-salinity desalination in the reclamation of domestic wastewater effluents. Chemosphere 2019, 235, 413–422. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, R.; Tian, Y.; Su, B.; Wang, K.; Yu, S.; Zhang, J.; Fang, J. Sterilization Efficiency of a Novel Electrochemical Disinfectant against Staphylococcus aureus. Environ. Sci. Technol. 2016, 50, 3184–3192. [Google Scholar] [CrossRef]
- Wang, T.C.; Qu, G.; Li, J.; Liang, D.; Hu, S. Depth dependence of p-nitrophenol removal in soil by pulsed discharge plasma. Chem. Eng. J. 2014, 239, 178–184. [Google Scholar] [CrossRef]
- Stryczewska, H.D.; Pawłat, J.; Ebihara, K. Non-Thermal Plasma Aided Soil Decontamination. J. Adv. Oxid. Technol. 2016, 16. [Google Scholar] [CrossRef] [Green Version]
- Štěpánová, V.; Slavíček, P.; Kelar, J.; Prášil, J.; Smékal, M.; Stupavská, M.; Jurmanová, J.; Černák, M. Atmospheric pressure plasma treatment of agricultural seeds of cucumber (Cucumis sativus L.) and pepper (Capsicum annuum L.) with effect on reduction of diseases and germination improvement. Plasma Process. Polym. 2017, 15, 1–9. [Google Scholar] [CrossRef]
- Abedi, S.; Iranbakhsh, A.; Ardebili, Z.O.; Ebadi, M. Seed priming with cold plasma improved early growth, flowering, and protection of Cichorium intybus against selenium nanoparticle. J. Theor. Appl. Phys. 2020, 14, 113–119. [Google Scholar] [CrossRef]
- Ma, R.; Wang, G.; Tian, Y.; Wang, K.; Zhang, J.; Fang, J. Non-thermal plasma-activated water inactivation of food-borne pathogen on fresh produce. J. Hazard. Mater. 2015, 300, 643–651. [Google Scholar] [CrossRef]
- Guo, J.; Huang, K.; Wang, X.; Lyu, C.; Yang, N.; Li, Y.; Wang, J. Inactivation of Yeast on Grapes by Plasma-Activated Water and Its Effects on Quality Attributes. J. Food Prot. 2017, 80, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.I.; Chen, W.; Liao, X.; Xiang, Q.; Liu, D.; Ye, X.; Ding, T. Effects of Plasma-Activated Water and Blanching on Microbial and Physicochemical Properties of Tiger Nuts. Food Bioprocess Technol. 2019, 12, 1721–1732. [Google Scholar] [CrossRef]
- Ghorashi, A.H.; Tasouji, M.A.R.; Kargarian, A. Optimum cold plasma generating device for treatment of Aspergillus flavus from nuts surface. J. Food Sci. Technol. 2020, 57, 3988–3994. [Google Scholar] [CrossRef]
- Hertwig, C.; Leslie, A.; Meneses, N.; Reineke, K.; Rauh, C.; Schlüter, O. Inactivation of Salmonella Enteritidis PT30 on the surface of unpeeled almonds by cold plasma. Innov. Food Sci. Emerg. Technol. 2017, 44, 242–248. [Google Scholar] [CrossRef]
- Šerá, B.; Šerý, M. Non-thermal plasma treatment as a new biotechnology in relation to seeds, dry fruits, and grains. Plasma Sci. Technol. 2018, 20, 044012. [Google Scholar] [CrossRef] [Green Version]
- Butscher, D.; Van Loon, H.; Waskow, A.; von Rohr, P.R.; Schuppler, M. Plasma inactivation of microorganisms on sprout seeds in a dielectric barrier discharge. Int. J. Food Microbiol. 2016, 238, 222–232. [Google Scholar] [CrossRef]
- Koga, K.; Thapanut, S.; Amano, T.; Seo, H.; Itagaki, N.; Hayashi, N.; Shiratani, M. Simple method of improving harvest by nonthermal air plasma irradiation of seeds of Arabidopsis thaliana (L.). Appl. Phys. Express 2016, 9, 016201. [Google Scholar] [CrossRef]
- Stolárik, T.; Henselová, M.; Martinka, M.; Novák, O.; Zahoranová, A.; Černák, M. Effect of Low-Temperature Plasma on the Structure of Seeds, Growth and Metabolism of Endogenous Phytohormones in Pea (Pisum sativum L.). Plasma Chem. Plasma Process. 2015, 35, 659–676. [Google Scholar] [CrossRef]
- Zahoranová, A.; Hoppanová, L.; Šimončicová, J.; Tučeková, Z.; Medvecká, V.; Hudecová, D.; Kaliňáková, B.; Kováčik, D.; Černák, M. Effect of Cold Atmospheric Pressure Plasma on Maize Seeds: Enhancement of Seedlings Growth and Surface Microorganisms Inactivation. Plasma Chem. Plasma Process. 2018, 38, 969–988. [Google Scholar] [CrossRef]
- Selcuk, M.; Oksuz, L.; Basaran, P. Decontamination of grains and legumes infected with Aspergillus spp. and Penicillum spp. by cold plasma treatment. Bioresour. Technol. 2008, 99, 5104–5109. [Google Scholar] [CrossRef] [PubMed]
- Waskow, A.; Betschart, J.; Butscher, D.; Oberbossel, G.; Klöti, D.; Büttner-Mainik, A.; Adamcik, J.; Von Rohr, P.R.; Schuppler, M. Characterization of Efficiency and Mechanisms of Cold Atmospheric Pressure Plasma Decontamination of Seeds for Sprout Production. Front. Microbiol. 2018, 9, 3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leymarie, J.; Vitkauskaité, G.; Hoang, H.H.; Gendreau, E.; Chazoule, V.; Meimoun, P.; Corbineau, F.; El-Maarouf-Bouteau, H.; Bailly, C. Role of Reactive Oxygen Species in the Regulation of Arabidopsis Seed Dormancy. Plant. Cell Physiol. 2012, 53, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ye, N.; Liu, R.; Chen, M.; Zhang, J. H2O2 mediates the regulation of ABA catabolism and GA biosynthesis in Arabidopsis seed dormancy and germination. J. Exp. Bot. 2010, 61, 2979–2990. [Google Scholar] [CrossRef] [Green Version]
- Filatova, I.; Azharonok, V.; Kadyrov, M.; Beljavsky, V.; Gvozdov, A.; Shik, A.; Antonuk, A. The effect of plasma treatment of seeds of some grain and legumes on their sowing quality and productivity. Rom. J. Phys. 2011, 56, 139–143. [Google Scholar]
- Bormashenko, E.; Grynyov, R.; Bormashenko, Y.; Drori, E. Cold Radiofrequency Plasma Treatment Modifies Wettability and Germination Speed of Plant Seeds. Sci. Rep. 2012, 2, 741. [Google Scholar] [CrossRef]
- Wannicke, N.; Wagner, R.; Stachowiak, J.; Nishime, T.M.C.; Ehlbeck, J.; Weltmann, K.; Brust, H. Efficiency of plasma-processed air for biological decontamination of crop seeds on the premise of unimpaired seed germination. Plasma Process. Polym. 2021, 18, 1–14. [Google Scholar] [CrossRef]
- Molina, R.; López-Santos, C.; Gómez-Ramírez, A.; Vílchez, A.; Espinós, J.P.; González-Elipe, A.R. Influence of irrigation conditions in the germination of plasma treated Nasturtium seeds. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor Determination in Soybean Seed by Multiple Criteria 1. Crop. Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-Dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Volin, J.C.; Denes, F.S.; Young, R.A.; Park, S.M.T. Modification of Seed Germination Performance through Cold Plasma Chemistry Technology. Crop. Sci. 2000, 40, 1706–1718. [Google Scholar] [CrossRef]
- Meng, Y.; Qu, G.; Wang, T.; Sun, Q.; Liang, D.; Hu, S. Enhancement of Germination and Seedling Growth of Wheat Seed Using Dielectric Barrier Discharge Plasma with Various Gas Sources. Plasma Chem. Plasma Process. 2017, 37, 1105–1119. [Google Scholar] [CrossRef]
- Sera, B.; Spatenka, P.; Sery, M.; Vrchotova, N.; Hruskova, I. Influence of Plasma Treatment on Wheat and Oat Germination and Early Growth. IEEE Trans. Plasma Sci. 2010, 38, 2963–2968. [Google Scholar] [CrossRef]
- Meiqiang, Y.; Mingjing, H.; Buzhou, M.; Tengcai, M. Stimulating Effects of Seed Treatment by Magnetized Plasma on Tomato Growth and Yield. Plasma Sci. Technol. 2005, 7, 3143–3147. [Google Scholar] [CrossRef]
- Park, Y.; Oh, K.S.; Oh, J.; Seok, D.C.; Kim, S.B.; Yoo, S.J.; Lee, M. The biological effects of surface dielectric barrier discharge on seed germination and plant growth with barley. Plasma Process. Polym. 2018, 15, e1600056. [Google Scholar] [CrossRef]
- Szőke, C.; Nagy, Z.; Gierczik, K.; Székely, A.; Spitkól, T.; Zsuboril, Z.T.; Galiba, G.; Marton, C.L.; Kutasi, K. Effect of the afterglows of low pressure Ar/N2-O2 surface-wave microwave discharges on barley and maize seeds. Plasma Process. Polym. 2018, 15, 1700138. [Google Scholar] [CrossRef]
- Henselová, M.; Slováková, Ľ.; Martinka, M.; Zahoranová, A. Growth, anatomy and enzyme activity changes in maize roots induced by treatment of seeds with low-temperature plasma. Biologia 2012, 67, 490–497. [Google Scholar] [CrossRef]
- Nishime, T.M.C.; Wannicke, N.; Horn, S.; Weltmann, K.-D.; Brust, H. A Coaxial Dielectric Barrier Discharge Reactor for Treatment of Winter Wheat Seeds. Appl. Sci. 2020, 10, 7133. [Google Scholar] [CrossRef]
- Chen, H.H.; Chen, Y.K.; Chang, H.C. Evaluation of physicochemical properties of plasma treated brown rice. Food Chem. 2012, 135, 74–79. [Google Scholar] [CrossRef]
- Šerá, B.; Sery, M.; Stranak, V.; Spatenka, P.; Tichy, M. Does Cold Plasma Affect Breaking Dormancy and Seed Germination? A Study on Seeds of Lamb’s Quarters (Chenopodium album agg.). Plasma Sci. Technol. 2009, 11, 750–754. [Google Scholar] [CrossRef]
- Švubová, R.; Kyzek, S.; Medvecká, V.; Slováková, Ľ.; Gálová, E.; Zahoranová, A. Novel insight at the Effect of Cold Atmospheric Pressure Plasma on the Activity of Enzymes Essential for the Germination of Pea (Pisum sativum L. cv. Prophet) Seeds. Plasma Chem. Plasma Process. 2020, 40, 1221–1240. [Google Scholar] [CrossRef]
- Li, L.; Jiang, J.F.; Li, J.; Shen, M.; He, X.; Shao, H.; Dong, Y. Effects of cold plasma treatment on seed germination and seedling growth of soybean. Sci. Rep. 2015, 4, srep05859. [Google Scholar] [CrossRef] [Green Version]
- Švubová, R.; Slováková, Ľ.; Holubová, Ľ.; Rovňanová, D.; Gálová, E.; Tomeková, J. Evaluation of the Impact of Cold Atmospheric Pressure Plasma on Soybean Seed Germination. Plants 2021, 10, 177. [Google Scholar] [CrossRef]
- Kyzek, S.; Holubová, Ľ.; Medvecká, V.; Tomeková, J.; Gálová, E.; Zahoranová, A. Cold Atmospheric Pressure Plasma Can Induce Adaptive Response in Pea Seeds. Plasma Chem. Plasma Process. 2018, 39, 475–486. [Google Scholar] [CrossRef]
- Tomeková, J.; Kyzek, S.; Medvecká, V.; Gálová, E.; Zahoranová, A. Influence of Cold Atmospheric Pressure Plasma on Pea Seeds: DNA Damage of Seedlings and Optical Diagnostics of Plasma. Plasma Chem. Plasma Process. 2020, 40, 1–14. [Google Scholar] [CrossRef]
- Sang, T. Genes and Mutations Underlying Domestication Transitions in Grasses: Figure 1. Plant. Physiol. 2009, 149, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.-M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant. Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef]
- Yang, L.; Xian, D.; Xiong, X.; Lai, R.; Song, J.; Zhong, J. Proanthocyanidins against Oxidative Stress: From Molecular Mechanisms to Clinical Applications. BioMed Res. Int. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dubinov, A.; Lazarenko, E.; Selemir, V. Effect of glow discharge air plasma on grain crops seed. IEEE Trans. Plasma Sci. 2000, 28, 180–183. [Google Scholar] [CrossRef]
- Masoni, A.; Pampana, S.; Arduini, I. Barley Response to Waterlogging Duration at Tillering. Crop. Sci. 2016, 56, 2722–2730. [Google Scholar] [CrossRef]
- Olsen, O.-A. ENDOSPERM DEVELOPMENT: Cellularization and Cell Fate Specification. Annu. Rev. Plant. Biol. 2001, 52, 233–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, O.-A. Nuclear Endosperm Development in Cereals and Arabidopsis thaliana. Plant. Cell 2004, 16, S214–S227. [Google Scholar] [CrossRef] [Green Version]
- Chrispeels, M.J.; Varner, J.E. Gibberellic Acid-Enhanced Synthesis and Release of α-Amylase and Ribonuclease by Isolated Barley and Aleurone Layers. Plant. Physiol. 1967, 42, 398–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCrate, A.J.; Nielsen, M.T.; Paulsen, G.M.; Heyne, E.G. Gibberellic acid effects on germination and α-amylase activity of winter wheats. Euphytica 1981, 30, 875–880. [Google Scholar] [CrossRef]
- Joshi, R. Role of enzymes in seed germination. Int. J. Creat. Res. Thoughts 2018, 6, 1481–1485. [Google Scholar]
- Los, A.; Ziuzina, D.; Boehm, D.; Cullen, P.J.; Bourke, P. Investigation of mechanisms involved in germination enhancement of wheat (Triticum aestivum) by cold plasma: Effects on seed surface chemistry and characteristics. Plasma Process. Polym. 2019, 16. [Google Scholar] [CrossRef]
- Yemeli, G.B.N.; Švubová, R.; Kostolani, D.; Kyzek, S.; Machala, Z. The effect of water activated by nonthermal air plasma on the growth of farm plants: Case of maize and barley. Plasma Process. Polym. 2021, 18. [Google Scholar] [CrossRef]
- O’Connell, D.; Cox, L.J.; Hyland, W.B.; McMahon, S.J.; Reuter, S.; Graham, W.G.; Gans, T.; Currell, F.J. Cold atmospheric pressure plasma jet interactions with plasmid DNA. Appl. Phys. Lett. 2011, 98, 043701. [Google Scholar] [CrossRef] [Green Version]
- Alkawareek, M.Y.; Alshraiedeh, N.H.; Higginbotham, S.; Flynn, P.B.; Algwari, Q.T.; Gorman, S.P.; Graham, W.G.; Gilmore, B.F. Plasmid DNA Damage Following Exposure to Atmospheric Pressure Nonthermal Plasma: Kinetics and Influence of Oxygen Admixture. Plasma Med. 2014, 4, 211–219. [Google Scholar] [CrossRef]
- Lee, G.J.; Choi, M.A.; Kim, D.; Kim, J.Y.; Ghimire, B.; Choi, E.H.; Kim, S.H. Influence of plasma-generated reactive species on the plasmid DNA structure and plasmid-mediated transformation of Escherichia coli cells. J. Appl. Phys. 2017, 122, 103303. [Google Scholar] [CrossRef]
- Arjunan, K.P.; Sharma, V.K.; Ptasinska, S. Effects of Atmospheric Pressure Plasmas on Isolated and Cellular DNA—A Review. Int. J. Mol. Sci. 2015, 16, 2971–3016. [Google Scholar] [CrossRef] [Green Version]
- Tendero, C.; Tixier, C.; Tristant, P.; Desmaison, J.; Leprince, P. Atmospheric pressure plasmas: A review. Spectrochim. Acta Part B At. Spectrosc. 2006, 61, 2–30. [Google Scholar] [CrossRef]
- Ehlbeck, J.; Schnabel, U.; Polak, M.; Winter, J.; Von Woedtke, T.; Brandenburg, R.; von dem Hagen, T.; Weltmann, K.-D. Low temperature atmospheric pressure plasma sources for microbial decontamination. J. Phys. D Appl. Phys. 2011, 44, 013002. [Google Scholar] [CrossRef] [Green Version]
- Setsuhara, Y. Low-temperature atmospheric-pressure plasma sources for plasma medicine. Arch. Biochem. Biophys. 2016, 605, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Černák, M.; Černáková, L.; Hudec, I.; Kováčik, D.; Zahoranová, A. Diffuse Coplanar Surface Barrier Discharge and its applications for in-line processing of low-added-value materials. Eur. Phys. J. Appl. Phys. 2009, 47, 22806. [Google Scholar] [CrossRef] [Green Version]
- Mošovská, S.; Medvecká, V.; Gregová, M.; Tomeková, J.; Valík, Ľ.; Mikulajová, A.; Zahoranová, A. Plasma inactivation of Aspergillus flavus on hazelnut surface in a diffuse barrier discharge using different working gases. Food Control. 2019, 104, 256–261. [Google Scholar] [CrossRef]
- Matusikova, V.; Salaj, J.; Moravčíková, J.; Mlynárová, L.; Nap, J.-P.; Libantová, J. Tentacles of in vitro-grown round-leaf sundew (Drosera rotundifolia L.) show induction of chitinase activity upon mimicking the presence of prey. Planta 2005, 222, 1020–1027. [Google Scholar] [CrossRef]
- Somogyi, M. Notes on sugar determination. J. Biol. Chem. 1952, 195, 19–23. [Google Scholar] [CrossRef]
- Nelson, N. A photometric adaptation of the Somogyi method for determination of glucose. J. Biol. Chem. 1944, 153, 375–380. [Google Scholar] [CrossRef]
- Frič, F.; Fuchs, W.H. Veränderungen der Aktivität einiger Enzyme im Weizenblatt in Abhängigkeit von der temperaturlabilen Verträglichkeit für Puccinia graminis tritici. J. Phytopathol. 1970, 67, 161–174. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Hartmann, A. Recommendations for conducting the in vivo alkaline Comet assay. Mutagenesis 2003, 18, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.R. The Comet Assay for DNA Damage and Repair: Principles, Applications, and Limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Gichner, T.; Patková, Z.; Száková, J.; Žnidar, I.; Mukherjee, A. DNA damage in potato plants induced by cadmium, ethyl methanesulphonate and γ-rays. Environ. Exp. Bot. 2008, 62, 113–119. [Google Scholar] [CrossRef]
- Collins, A.R. Measuring oxidative damage to DNA and its repair with the comet assay. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2014, 1840, 794–800. [Google Scholar] [CrossRef]
- Chankova, S.G.; Bryant, P.E. Acceleration of DNA-double strand rejoining during the adaptive response of Chlamydomonas reinhardtii. Radiat. Biol. Radioecol. 2002, 42, 600–603. [Google Scholar]
- Muslimović, A.; Nyström, S.; Gao, Y.; Hammarsten, O. Numerical Analysis of Etoposide Induced DNA Breaks. PLoS ONE 2009, 4, e5859. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAPP Application (s) | Average Shoot Length (mm) | Average Shoot Weight (mg) | Average Root Length (mm) | Average Root Weight (mg) | Adventitious Root Number |
|---|---|---|---|---|---|
| C (0) | 11.3 ± 0.43 d | 81.9 ± 17.2 e | 6.21 ± 0.96 e | 69.5 ± 16.1 c | 6.66 ± 0.59 d |
| O10 | 10.2 ± 1.11 b | 81.1 ± 19.3 de | 7.49 ± 1.33 f | 82.7 ± 22.2 de | 6.40 ± 0.70 abcd |
| O20 | 10.3 ± 1.91 bc | 79.4 ± 23.1 cde | 5.38 ± 1.13 bcd | 93.0 ± 23.6 e | 6.18 ± 0.60 abc |
| O30 | 11.3 ± 0.88 cd | 97.5 ± 21.7 e | 4.17 ± 1.11 a | 97.0 ± 15.7 e | 5.85 ± 0.69 a |
| O60 | 5.0 ± 1.05 a | 63.2 ± 13.3 ab | 4.61 ± 1.32 ab | 58.8 ± 17.2 ab | 6.47 ± 0.68 bcd |
| O180 | 4.9 ± 0.98 a | 65.0 ± 14.3 abc | 5.12 ± 1.29 bcd | 48.8 ± 15.5 a | 6.17 ± 0.85 ab |
| A10 | 11.3 ± 1.11 d | 86.4 ± 22.6 ef | 6.55 ± 0.84 ef | 84.7 ± 24.7 de | 6.60 ± 0.70 bcd |
| A20 | 11.0 ± 1.55 bcd | 87.3 ± 22.3 ef | 6.48 ± 0.77 e | 68.5 ± 19.6 bc | 6.90 ± 0.57 d |
| A30 | 10.9 ± 1.01 bcd | 87.8 ± 22.2 ef | 4.70 ± 0.78 abcd | 75.0 ± 21.4 cd | 6.75 ± 0.46 bcd |
| A60 | 5.3 ± 1.01 a | 54.7 ± 17.0 a | 4.78 ± 0.99 abc | 54.2 ± 17.3 ab | 6.67 ± 0.93 d |
| N10 | 11.5 ± 0.63 d | 80.1 ± 18.6 de | 5.51 ± 1.25 c | 76.4 ± 18.5 cd | 6.51 ± 0.76 bcd |
| N20 | 10.9 ± 1.64 bcd | 72.5 ± 17.7 bcd | 5.45 ± 0.59 cd | 53.3 ± 17.6 a | 6.65 ± 0.85 cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peťková, M.; Švubová, R.; Kyzek, S.; Medvecká, V.; Slováková, Ľ.; Ševčovičová, A.; Gálová, E. The Effects of Cold Atmospheric Pressure Plasma on Germination Parameters, Enzyme Activities and Induction of DNA Damage in Barley. Int. J. Mol. Sci. 2021, 22, 2833. https://doi.org/10.3390/ijms22062833
Peťková M, Švubová R, Kyzek S, Medvecká V, Slováková Ľ, Ševčovičová A, Gálová E. The Effects of Cold Atmospheric Pressure Plasma on Germination Parameters, Enzyme Activities and Induction of DNA Damage in Barley. International Journal of Molecular Sciences. 2021; 22(6):2833. https://doi.org/10.3390/ijms22062833
Chicago/Turabian StylePeťková, Mária, Renáta Švubová, Stanislav Kyzek, Veronika Medvecká, Ľudmila Slováková, Andrea Ševčovičová, and Eliška Gálová. 2021. "The Effects of Cold Atmospheric Pressure Plasma on Germination Parameters, Enzyme Activities and Induction of DNA Damage in Barley" International Journal of Molecular Sciences 22, no. 6: 2833. https://doi.org/10.3390/ijms22062833