
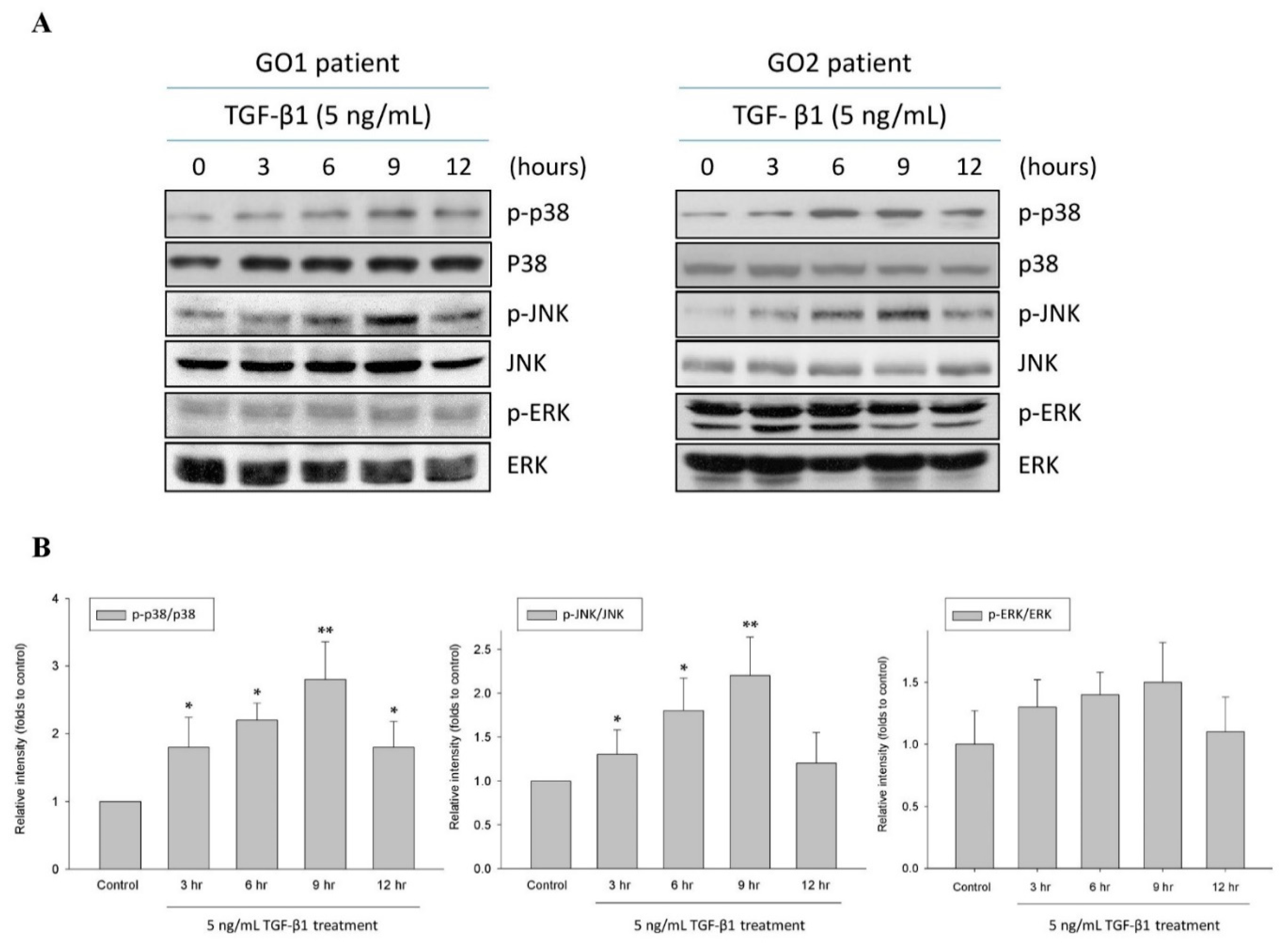
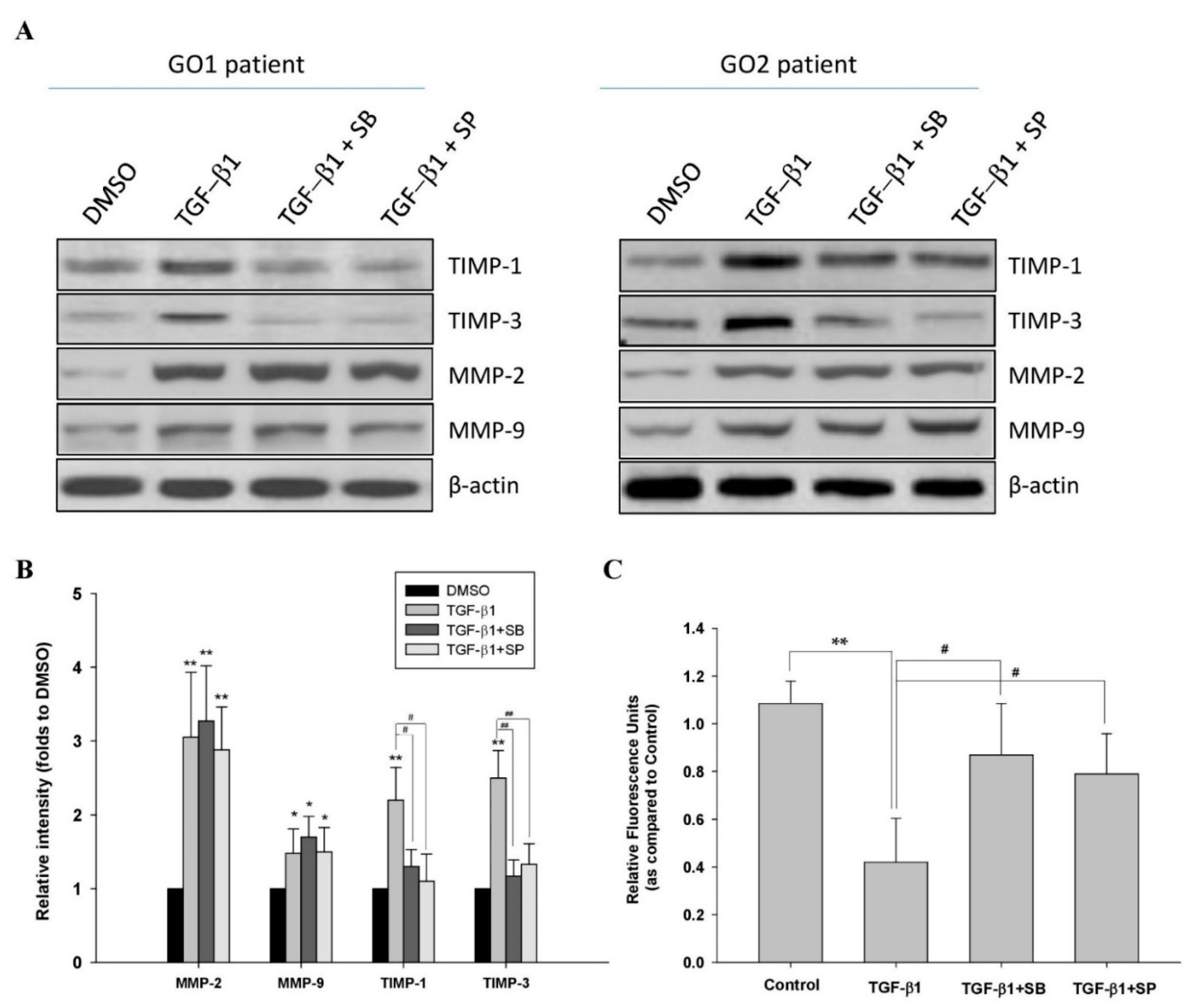
JNK and p38 Inhibitors Prevent Transforming Growth Factor-β1-Induced Myofibroblast Transdifferentiation in Human Graves’ Orbital Fibroblasts




{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Hou, T.-Y.; Wu, S.-B.; Kau, H.-C.; Tsai, C.-C. JNK and p38 Inhibitors Prevent Transforming Growth Factor-β1-Induced Myofibroblast Transdifferentiation in Human Graves’ Orbital Fibroblasts. Int. J. Mol. Sci. 2021, 22, 2952. https://doi.org/10.3390/ijms22062952
Hou T-Y, Wu S-B, Kau H-C, Tsai C-C. JNK and p38 Inhibitors Prevent Transforming Growth Factor-β1-Induced Myofibroblast Transdifferentiation in Human Graves’ Orbital Fibroblasts. International Journal of Molecular Sciences. 2021; 22(6):2952. https://doi.org/10.3390/ijms22062952
Chicago/Turabian StyleHou, Tzu-Yu, Shi-Bei Wu, Hui-Chuan Kau, and Chieh-Chih Tsai. 2021. "JNK and p38 Inhibitors Prevent Transforming Growth Factor-β1-Induced Myofibroblast Transdifferentiation in Human Graves’ Orbital Fibroblasts" International Journal of Molecular Sciences 22, no. 6: 2952. https://doi.org/10.3390/ijms22062952
APA StyleHou, T.-Y., Wu, S.-B., Kau, H.-C., & Tsai, C.-C. (2021). JNK and p38 Inhibitors Prevent Transforming Growth Factor-β1-Induced Myofibroblast Transdifferentiation in Human Graves’ Orbital Fibroblasts. International Journal of Molecular Sciences, 22(6), 2952. https://doi.org/10.3390/ijms22062952