
Grainyhead-Like 3 Influences Migration and Invasion of Urothelial Carcinoma Cells

 ,
,
Abstract
:1. Introduction
2. Results
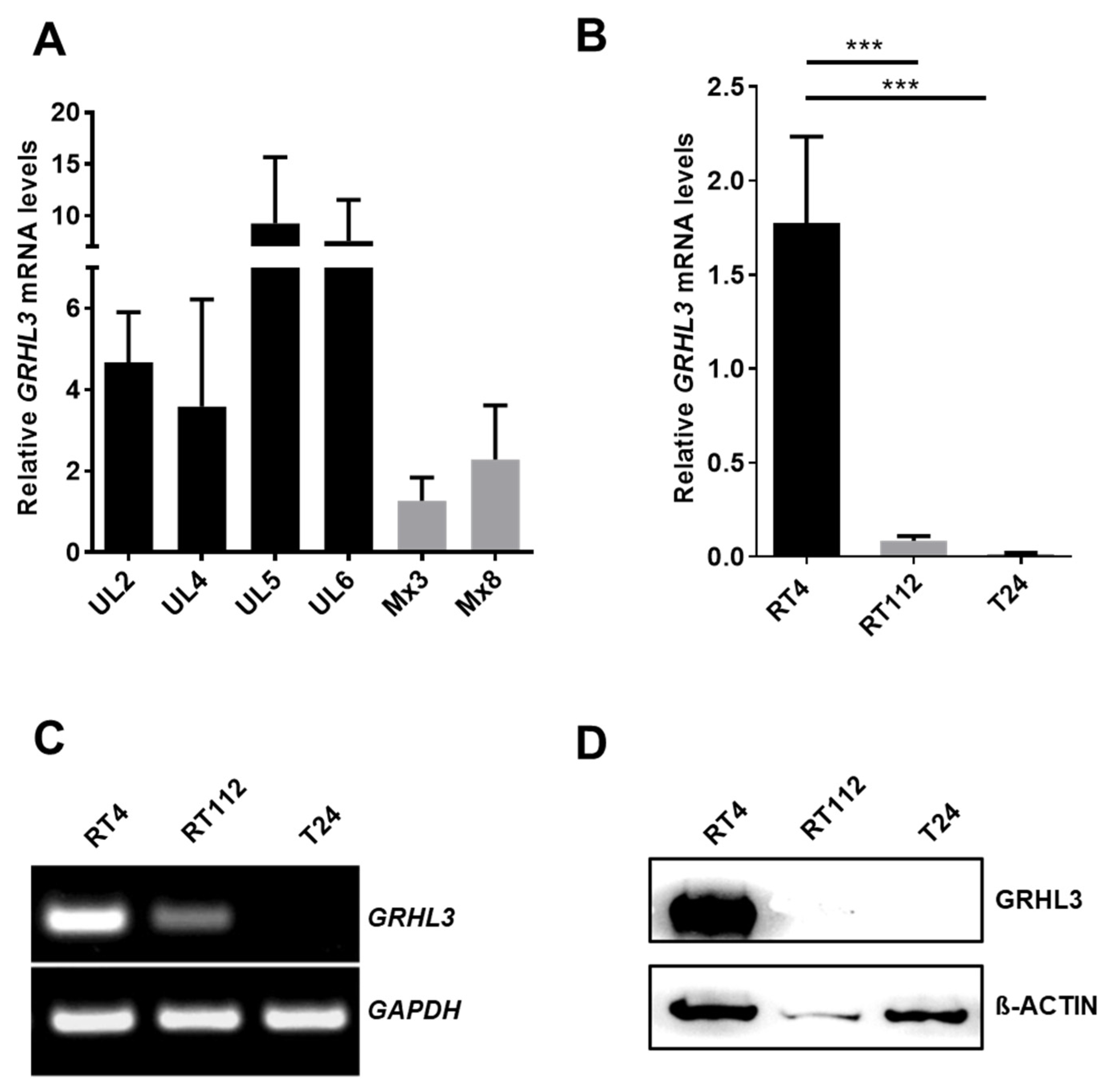
2.1. GRHL3 Expression Is Downregulated in Bladder Cancer Cells
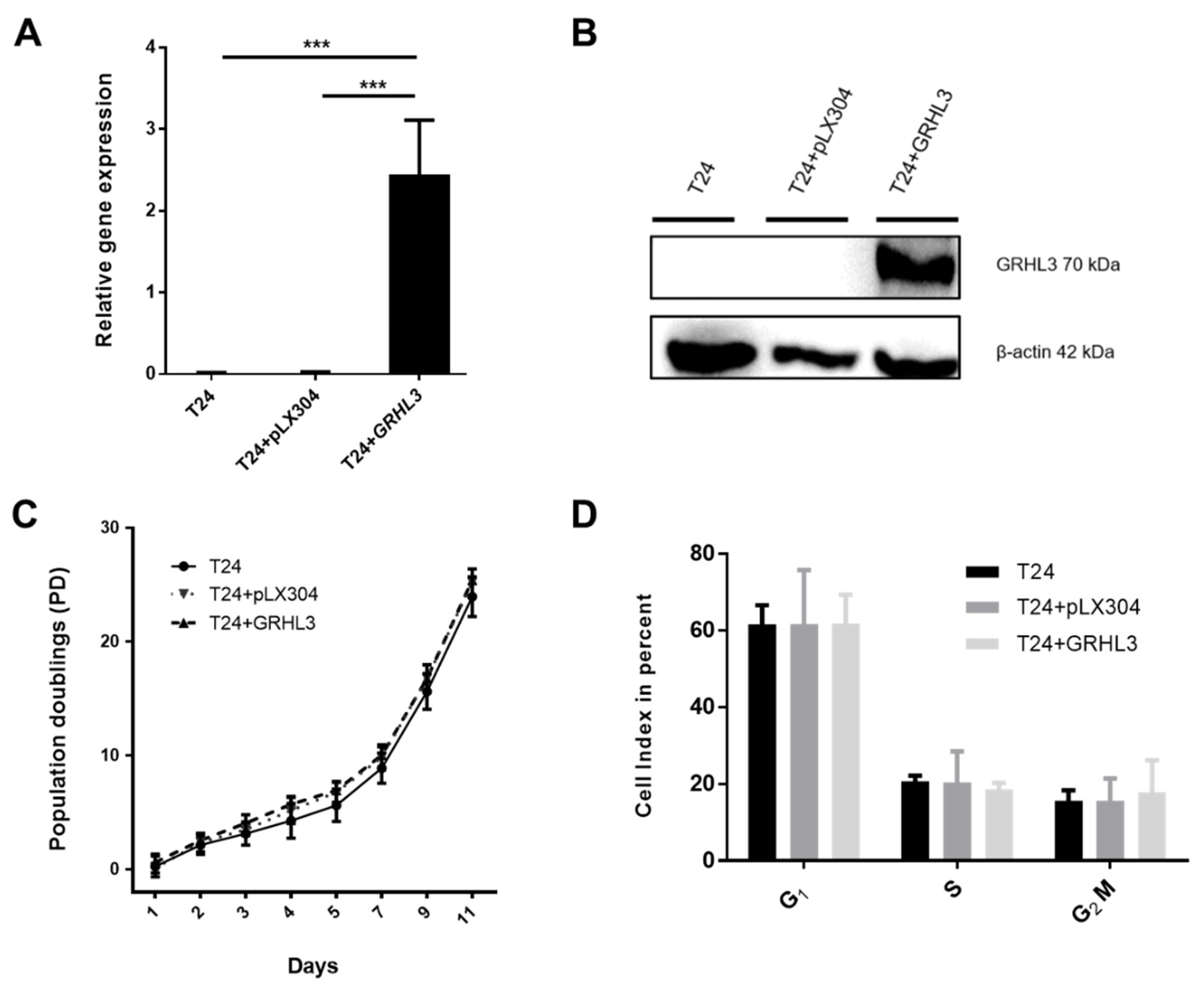
2.2. Ectopic GRHL3 Does Not Influence Cell Survival and Cell Cycle Profile of T24 Cells
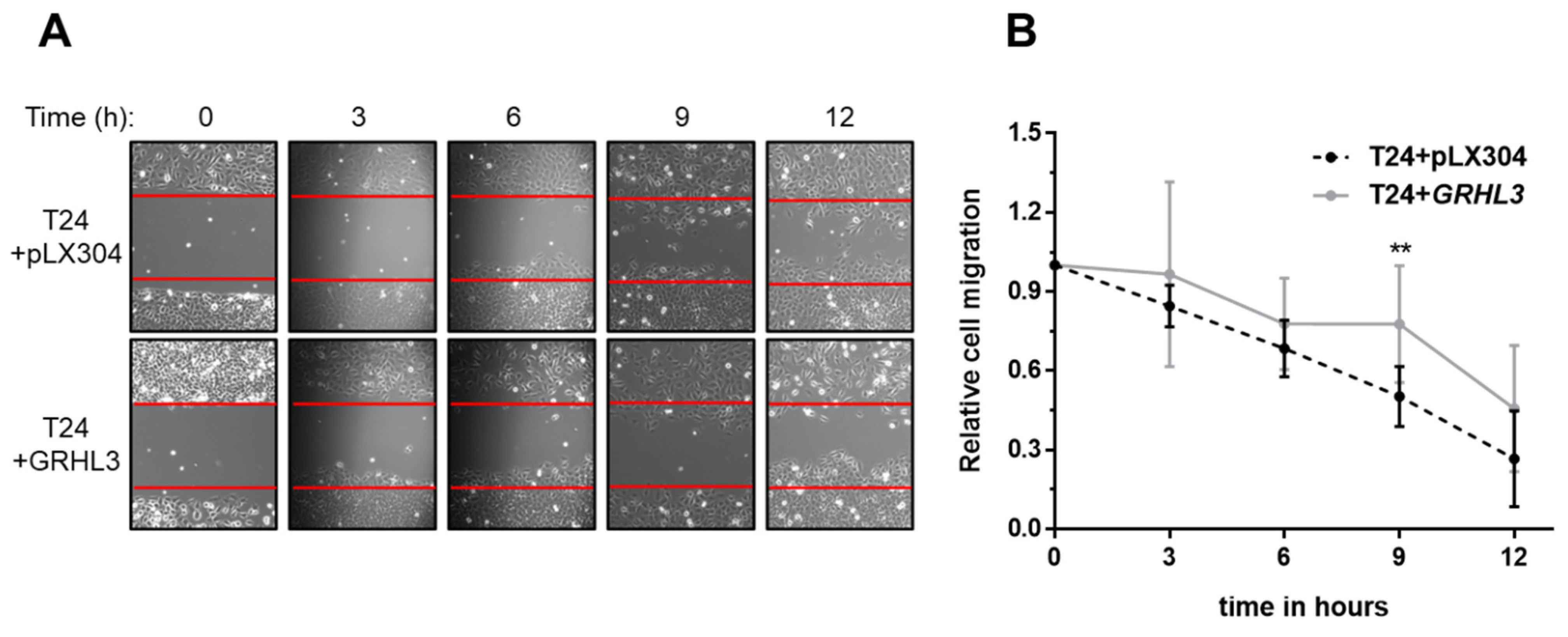
2.3. Ectopic GRHL3 Impairs Migration Capacity of T24 Cells
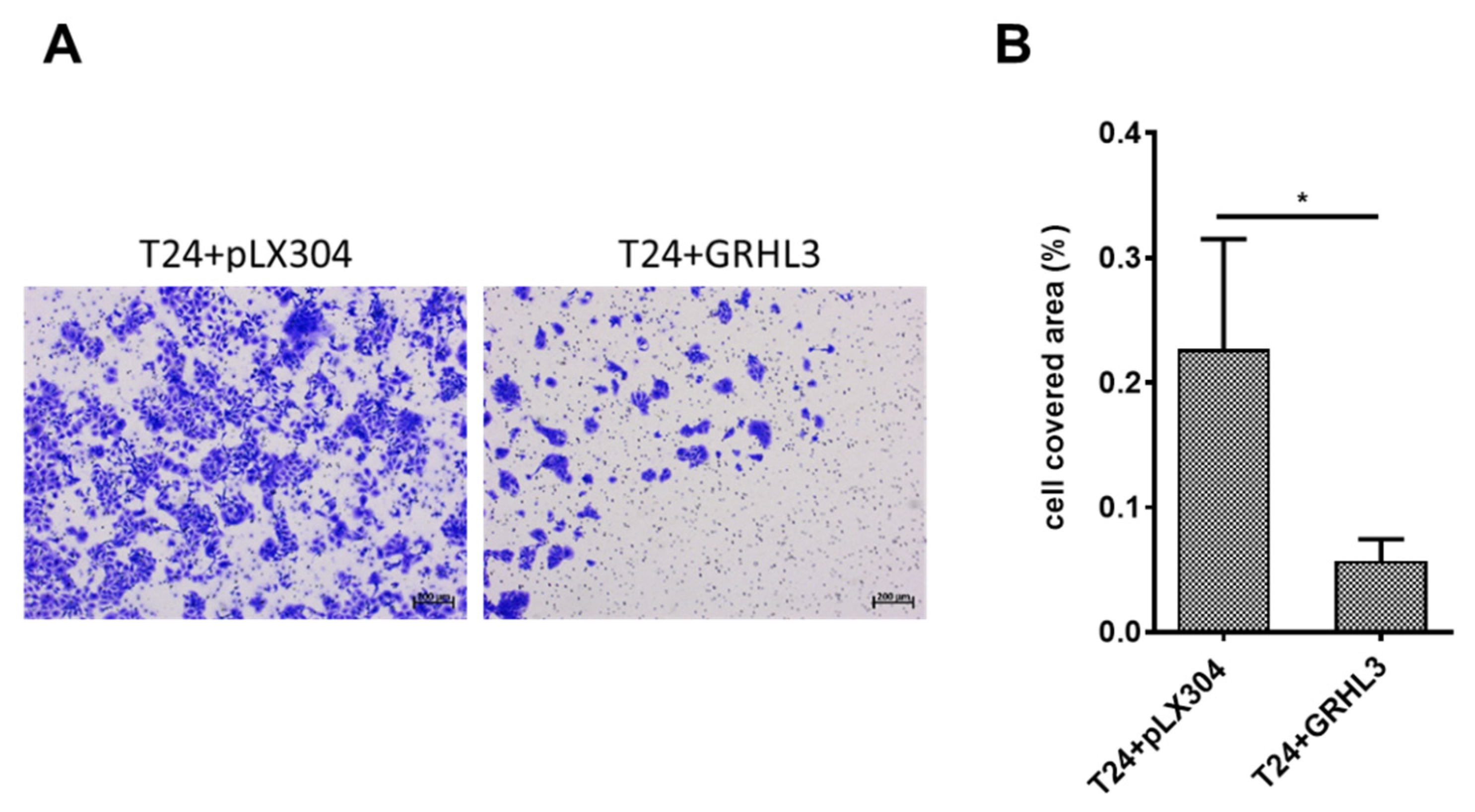
2.4. Forced Ectopic GRHL3 Expression Impairs Invasion Capacity of T24 Cells
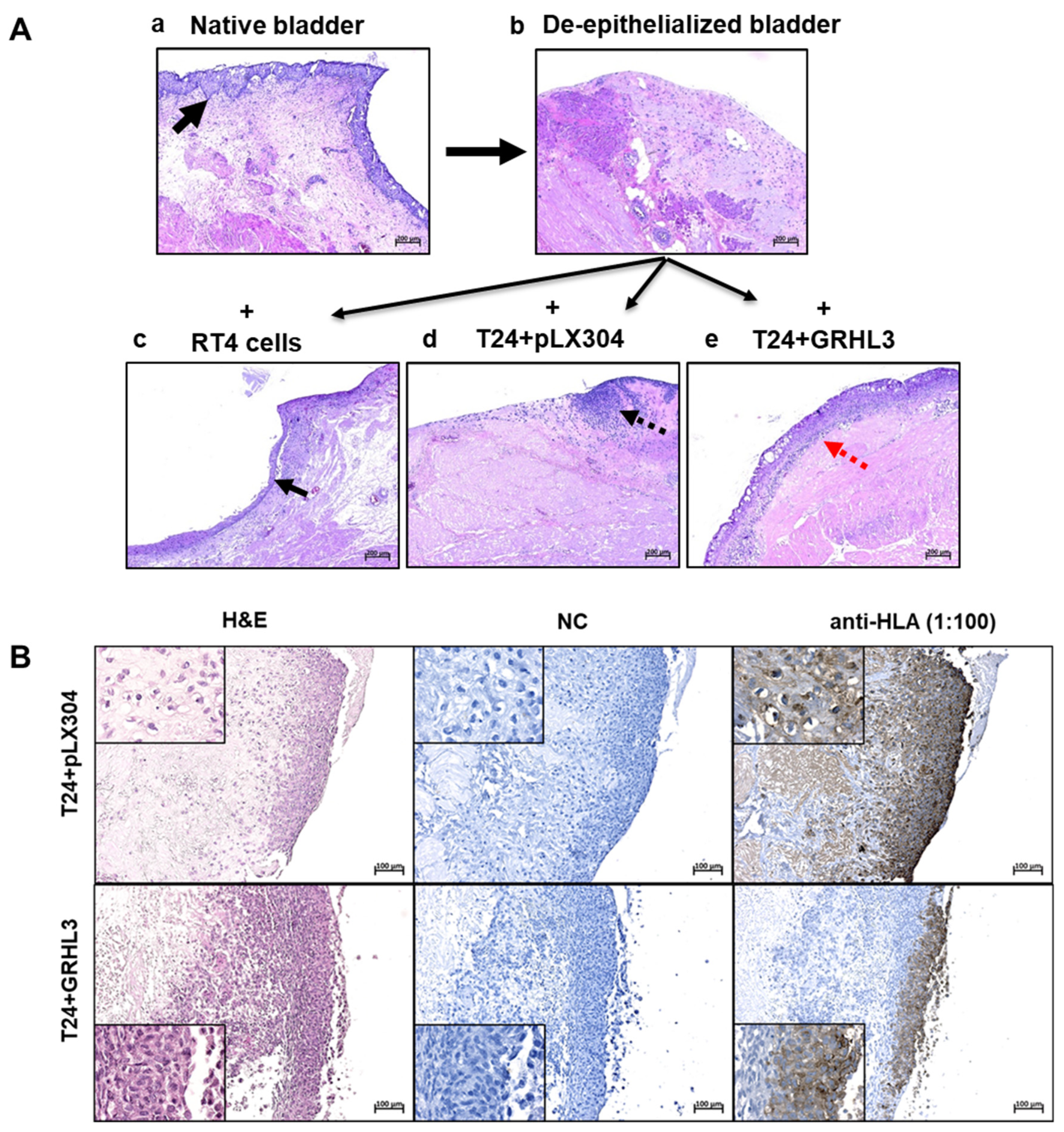
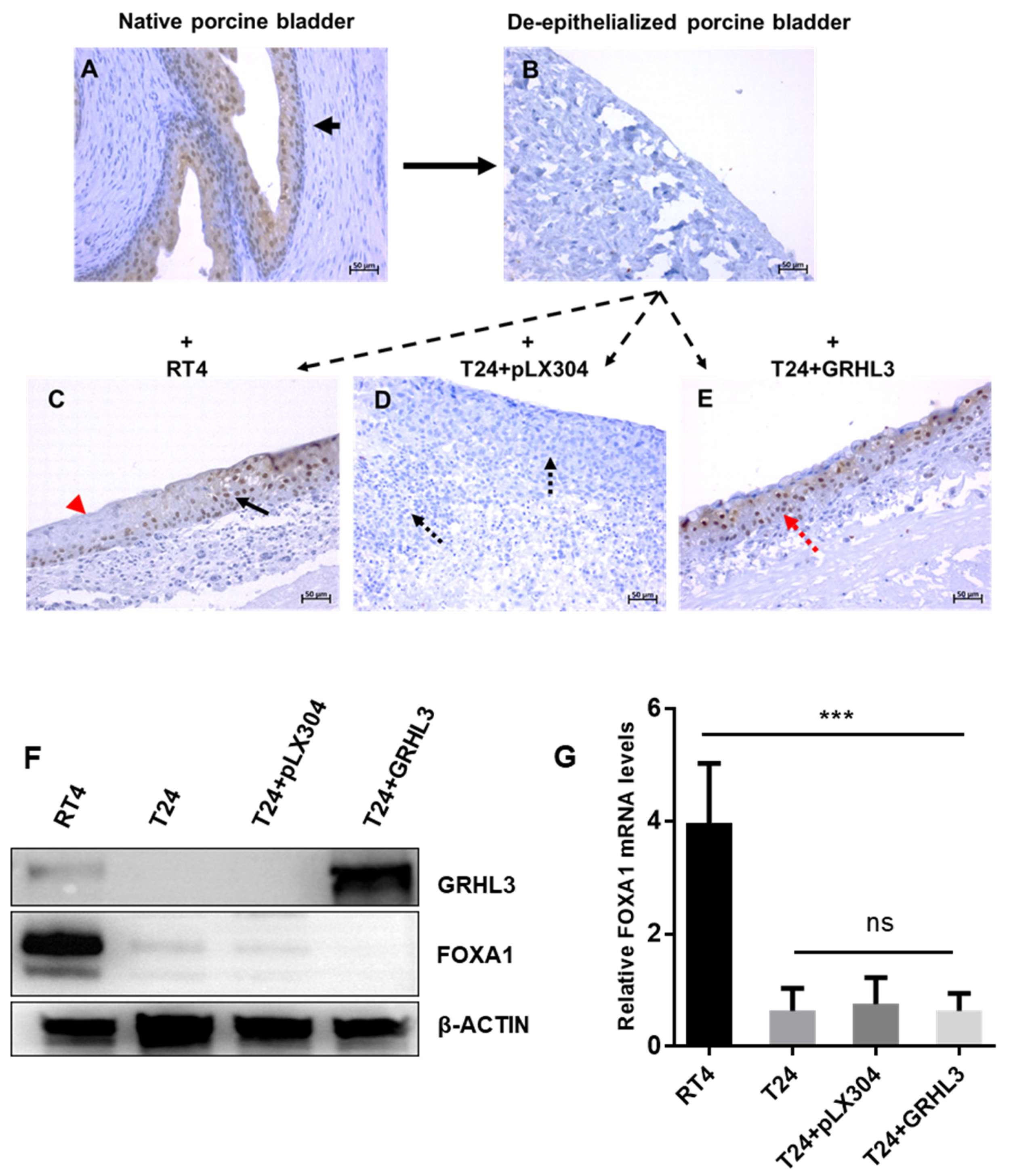
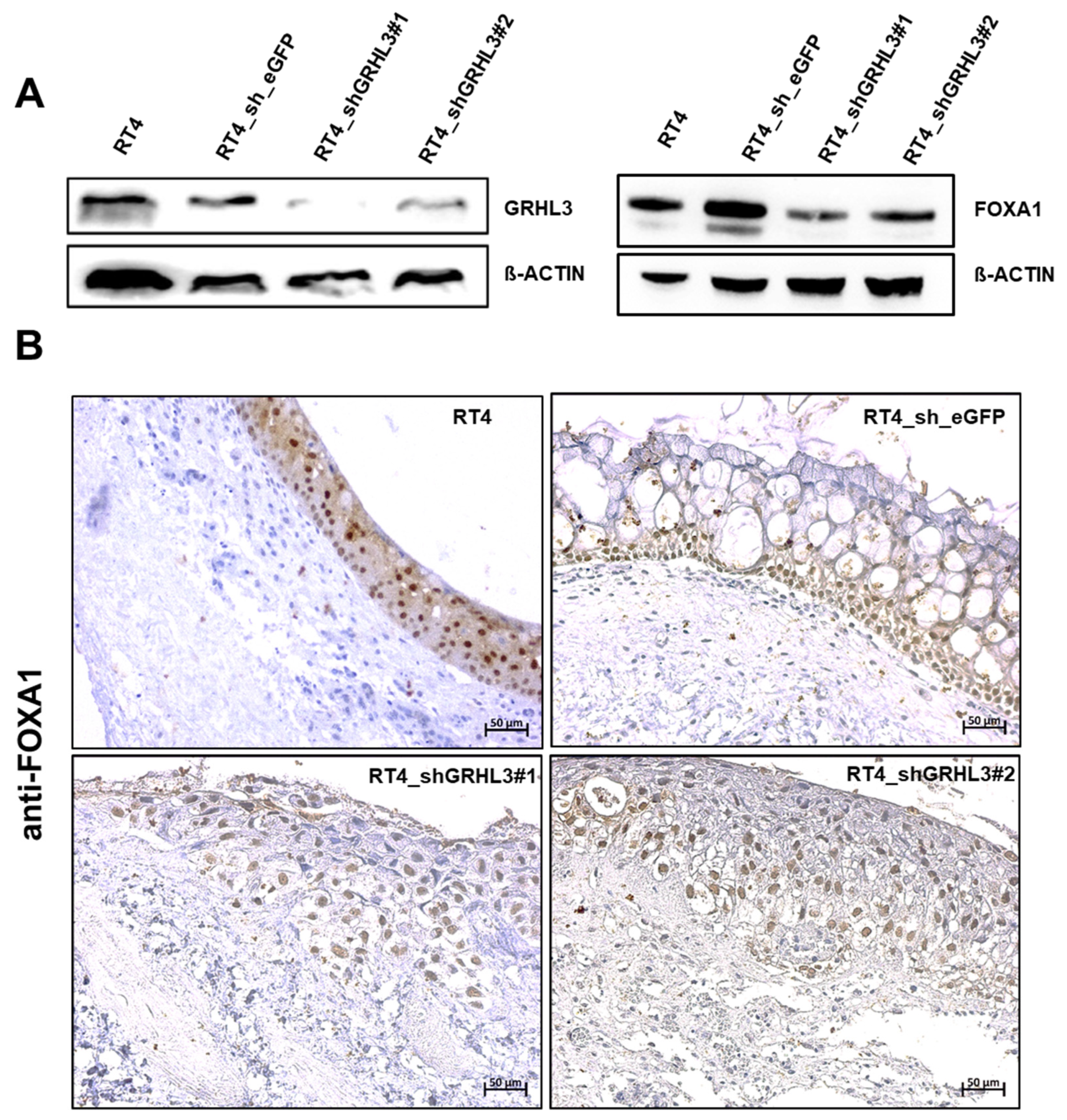
2.5. GRLH3 Knockdown Increases Invasive Potential of RT4 in the Ex Vivo Organ Culture Model
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Ectopic Expression of GRHL3 T24 Cells
4.3. shRNAs and Transfection in RT4 Cells
4.4. PCR Primer
4.5. Western Blots
4.6. Extraction of Genomic DNA
4.7. Extraction of Total RNA
4.8. Reverse Transcription (RT)
4.9. Polymerase Chain Reaction (PCR) and Real-Time
4.10. Real-Time Quantitative PCR (RT-qPCR)
4.11. Population Doubling Assay
4.12. Wound Healing Assay
4.13. Boyden Chamber Assay
4.14. Porcine Bladder Organ Culture Model
4.15. Data Processing, Statistical Analyses and Graphic Representations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seiler, R.; Ashab, H.A.D.; Erho, N.; van Rhijn, B.W.G.; Winters, B.; Douglas, J.; Van Kessel, K.E.; Fransen van de Putte, E.E.; Sommerlad, M.; Wang, N.Q.; et al. Impact of Molecular Subtypes in Muscle-invasive Bladder Cancer on Predicting Response and Survival after Neoadjuvant Chemotherapy. Eur. Urol. 2017, 72, 544–554. [Google Scholar] [CrossRef]
- Kamoun, A.; de Reynies, A.; Allory, Y.; Sjodahl, G.; Robertson, A.G.; Seiler, R.; Hoadley, K.A.; Groeneveld, C.S.; Al-Ahmadie, H.; Choi, W.; et al. A Consensus Molecular Classification of Muscle-invasive Bladder Cancer. Eur. Urol. 2020, 77, 420–433. [Google Scholar] [CrossRef]
- Robertson, A.G.; Kim, J.; Al-Ahmadie, H.; Bellmunt, J.; Guo, G.; Cherniack, A.D.; Hinoue, T.; Laird, P.W.; Hoadley, K.A.; Akbani, R.; et al. Comprehensive Molecular Characterization of Muscle-Invasive Bladder Cancer. Cell 2017, 171, 540–556.e525. [Google Scholar] [CrossRef] [Green Version]
- Volkmer, J.P.; Sahoo, D.; Chin, R.K.; Ho, P.L.; Tang, C.; Kurtova, A.V.; Willingham, S.B.; Pazhanisamy, S.K.; Contreras-Trujillo, H.; Storm, T.A.; et al. Three differentiation states risk-stratify bladder cancer into distinct subtypes. Proc. Natl. Acad. Sci. USA 2012, 109, 2078–2083. [Google Scholar] [CrossRef] [Green Version]
- Choi, W.; Porten, S.; Kim, S.; Willis, D.; Plimack, E.R.; Hoffman-Censits, J.; Roth, B.; Cheng, T.; Tran, M.; Lee, I.L.; et al. Identification of distinct basal and luminal subtypes of muscle-invasive bladder cancer with different sensitivities to frontline chemotherapy. Cancer Cell 2014, 25, 152–165. [Google Scholar] [CrossRef] [Green Version]
- Bell, S.M.; Zhang, L.; Mendell, A.; Xu, Y.; Haitchi, H.M.; Lessard, J.L.; Whitsett, J.A. Kruppel-like factor 5 is required for formation and differentiation of the bladder urothelium. Dev. Biol. 2011, 358, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Mannik, J.; Soto, A.; Lin, K.K.; Andersen, B. The epidermal differentiation-associated Grainyhead gene Get1/Grhl3 also regulates urothelial differentiation. EMBO J. 2009, 28, 1890–1903. [Google Scholar] [CrossRef] [Green Version]
- Bock, M.; Hinley, J.; Schmitt, C.; Wahlicht, T.; Kramer, S.; Southgate, J. Identification of ELF3 as an early transcriptional regulator of human urothelium. Dev. Biol. 2014, 386, 321–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborn, S.L.; Thangappan, R.; Luria, A.; Lee, J.H.; Nolta, J.; Kurzrock, E.A. Induction of human embryonic and induced pluripotent stem cells into urothelium. Stem Cells Transl. Med. 2014, 3, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Frisch, S.M.; Farris, J.C.; Pifer, P.M. Roles of Grainyhead-like transcription factors in cancer. Oncogene 2017, 36, 6067–6073. [Google Scholar] [CrossRef]
- Liu, W.; Ha, M.; Wang, X.; Yin, N. Clinical significance of GRHL3 expression in diffuse large B cell lymphoma. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 9657–9661. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.K.; Zhou, F.F.; Tao, H.R.; Wang, X.; Zhang, C.; Su, F.; Wang, S.P.; Xu, L.H.; Pan, X.K.; Feng, M.H.; et al. Knockdown of GRHL3 inhibits activities and induces cell cycle arrest and apoptosis of human colorectal cancer cells. J. Huazhong Univ. Sci. Tech. Med. Sci. 2017, 37, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Taniwaki, M.; Daigo, Y.; Ishikawa, N.; Takano, A.; Tsunoda, T.; Yasui, W.; Inai, K.; Kohno, N.; Nakamura, Y. Gene expression profiles of small-cell lung cancers: Molecular signatures of lung cancer. Int. J. Oncol. 2006, 29, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Guo, S.; Tu, Z.; Di, L.; Zha, X.; Zhou, H.; Zhang, X. Grhl3 induces human epithelial tumor cell migration and invasion via downregulation of E-cadherin. Acta Biochim. Biophys. Sin. 2016, 48, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Liu, C.; Zhao, Z.; Gao, N.; Chen, G.; Wang, Y.; Cui, J. Clinical implications of GRHL3 protein expression in breast cancer. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 1827–1831. [Google Scholar] [CrossRef]
- Li, J.; Yen, C.; Liaw, D.; Podsypanina, K.; Bose, S.; Wang, S.I.; Puc, J.; Miliaresis, C.; Rodgers, L.; McCombie, R.; et al. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. N. Y. Sci. J. 1997, 275, 1943–1947. [Google Scholar] [CrossRef]
- Darido, C.; Georgy, S.; Wilanowski, T.; Dworkin, S.; Auden, A.; Zhao, Q.; Rank, G.; Srivastava, S.; Finlay, M.; Papenfuss, A.; et al. Targeting of the Tumor Suppressor GRHL3 by a miR-21-Dependent Proto-Oncogenic Network Results in PTEN Loss and Tumorigenesis. Cancer Cell 2011, 20, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, A.; Gordon, W.; Dizon, D.; Hopkin, A.S.; Gordon, E.; Yu, Z.; Andersen, B. The Grainyhead transcription factor Grhl3/Get1 suppresses miR-21 expression and tumorigenesis in skin: Modulation of the miR-21 target MSH2 by RNA-binding protein DND1. Oncogene 2013, 32, 1497–1507. [Google Scholar] [CrossRef] [Green Version]
- Booth, C.; Harnden, P.; Trejdosiewicz, L.K.; Scriven, S.; Selby, P.J.; Southgate, J. Stromal and vascular invasion in an human in vitro bladder cancer model. Lab. Investig. 1997, 76, 843–857. [Google Scholar]
- Fujiyama, C.; Jones, A.; Fuggle, S.; Bicknell, R.; Cranston, D.; Harris, A.L. Human bladder cancer invasion model using rat bladder in vitro and its use to test mechanisms and therapeutic inhibitors of invasion. Br. J. Cancer 2001, 84, 558–564. [Google Scholar] [CrossRef] [Green Version]
- Puzio-Kuter, A.M.; Castillo-Martin, M.; Kinkade, C.W.; Wang, X.; Shen, T.H.; Matos, T.; Shen, M.M.; Cordon-Cardo, C.; Abate-Shen, C. Inactivation of p53 and Pten promotes invasive bladder cancer. Genes Dev. 2009, 23, 675–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgescu, M.M. PTEN Tumor Suppressor Network in PI3K-Akt Pathway Control. Genes Cancer 2010, 1, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Mlacki, M.; Kikulska, A.; Krzywinska, E.; Pawlak, M.; Wilanowski, T. Recent discoveries concerning the involvement of transcription factors from the Grainyhead-like family in cancer. Exp. Biol. Med. 2015, 240, 1396–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjostedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McConkey, D.J.; Choi, W.; Marquis, L.; Martin, F.; Williams, M.B.; Shah, J.; Svatek, R.; Das, A.; Adam, L.; Kamat, A.; et al. Role of epithelial-to-mesenchymal transition (EMT) in drug sensitivity and metastasis in bladder cancer. Cancer Metastas. Rev. 2009, 28, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Wezel, F.; Pearson, J.; Southgate, J. Plasticity of in vitro-generated urothelial cells for functional tissue formation. Tissue Eng. Part A 2014, 20, 1358–1368. [Google Scholar] [CrossRef]
- Chopra, B.; Hinley, J.; Oleksiewicz, M.B.; Southgate, J. Trans-species comparison of PPAR and RXR expression by rat and human urothelial tissues. Toxicol. Pathol. 2008, 36, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Southgate, J.; Hutton, K.A.; Thomas, D.F.; Trejdosiewicz, L.K. Normal human urothelial cells in vitro: Proliferation and induction of stratification. Lab. Investig. 1994, 71, 583–594. [Google Scholar]
- Kumar, M.; Witt, B.; Knippschild, U.; Koch, S.; Meena, J.K.; Heinlein, C.; Weise, J.M.; Krepulat, F.; Kuchenbauer, F.; Iben, S.; et al. CEBP factors regulate telomerase reverse transcriptase promoter activity in whey acidic protein-T mice during mammary carcinogenesis. Int. J. Cancer. 2013, 9, 2032–2043. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | Plasmid ID | Cat No | Sequence in 5′-3′ |
|---|---|---|---|
| shGRHL3#1 | NM_021180.2-439s1c1 | NM_021180.2 | CCGGGCTCAAGAAGAATAACCTGATCTCGAGATCAGGTTATTCTTCTTGAGCTTTTT |
| shGRHL3#2 | NM_021180.2-236s1c1 | NM_021180.2 | CCGGGCCTTGAGCTTCCTCTATGATCTCGAGATCATAGAGGAAGCTCAAGGCTTTTT |
| sheGFP | MISSION® pLKO.1-puro eGFP shRNA Control | SHC005 | CCGGTACAACAGCCACAACGTCTATCTCGAGATAGACGTTGTGGCTGTTGTATTTTT |
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| Forkhead box protein A1 (FOXA1) (qPCR) | ACTGTGAAGATGGAAGGGCA | AGTAGGCCTCCCTGCTTGT |
| Glyceraldehyde 3-phosphate Dehydrogenase (GAPDH) (qPCR) | AGATCCCTCCAAAATCAAGTGG | AAAAGGGTCATCATCTCTGCC |
| Grainyhead Like Transcription Factor (GRHL3) (qPCR, RT-PCR) | ATTGACGTGGCTGACTGCAA | GCTCAGACAGTTTACGCCGA |
| Peroxisome proliferator-activated receptor gamma (PPARγ) (qPCR) | TATTCTCAGTGGAGACCGCC | AGGGCTTGTAGCAGGTTGTC |
| Phosphatase and tensin homolog (PTEN) (qPCR) | GGCACCGCATATTAAAACGTA | ATGCCATTTTTCCATTTCCA |
| Uroplakin 1a (UPK1a) (qPCR) | GTGGTGGGCCTGCTAGTTG | TATACACGCTACTGGTCGGCT |
| Uroplakin 1b (UPK1b) (qPCR) | CCAAAGACAACTCAACTGTTCGT | AATGCCGCAACAACCAATAATC |
| Uroplakin 2 (UPK2) (qPCR) | ACCAGGTGACAAACCTCGTG | TGTTCCTTCGAGGGAGTGTG |
| Uroplakin 3a (UPK3a) (qPCR) | CGTGGACATGGGGAGTTCTG | TCACGGACGTGTAGGAAGACT |
| Uroplakin 3b (UPK3b) (qPCR) | CCTCTACCATGCGCTTCTCC | ATGTGGTGGGTCATGTAGCG |
| GRHL3-3′-untranslated region (UTR) (RT-PCR) | CACACAACCTCTCCACATGC | TCAGGGAGCAGATTCAAGCA |
| GRHL3-coding (RT-PCR) | TCAACGGAAAAGCAGTGTGG | CGTTAGGCCGTGCTTCTTAC |
| Scheme | Antigen | Clone | Dilution | Provider |
|---|---|---|---|---|
| Mouse | GRHL3 | C-12, sc-398838, IgG2b | 1:1000 | Santa Cruz, Dallas, TX, USA |
| Mouse | GRHL3 | Aa529-578 | 1:500 | Lifespan Biosciences, Seattle, WA, USA |
| Horse | Mouse IgG | HRP-linked, #7076 | 1:2000 | Cell Signaling Technology, Danvers, MA, USA |
| Rabbit | Total Akt | #75692S (D9R8K) | 1:2000 | Cell Signaling Technology, Danvers, MA, USA |
| Rabbit | Phospho-Akt (Ser473) | #4060 (D9E) | 1:2000 | Cell Signaling Technology, Danvers, MA, USA |
| Rabbit | PTEN | #9559 (138G6) | 1:1000 | Cell Signaling Technology, Danvers, MA, USA |
| Chicken | Rabbit IgG | HRP-linked, sc-2955 | 1:2000 | Santa Cruz, Dallas, TX, USA |
| Mouse | β-Actin | AC-15 | 1:10,000 | Sigma-Aldrich, St. Louis, MO, USA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wezel, F.; Lustig, J.; Azoitei, A.; Liu, J.; Meessen, S.; Najjar, G.; Zehe, V.; Faustmann, P.; Zengerling, F.; John, A.; et al. Grainyhead-Like 3 Influences Migration and Invasion of Urothelial Carcinoma Cells. Int. J. Mol. Sci. 2021, 22, 2959. https://doi.org/10.3390/ijms22062959
Wezel F, Lustig J, Azoitei A, Liu J, Meessen S, Najjar G, Zehe V, Faustmann P, Zengerling F, John A, et al. Grainyhead-Like 3 Influences Migration and Invasion of Urothelial Carcinoma Cells. International Journal of Molecular Sciences. 2021; 22(6):2959. https://doi.org/10.3390/ijms22062959
Chicago/Turabian StyleWezel, Felix, Johannes Lustig, Anca Azoitei, Junnan Liu, Sabine Meessen, Gregoire Najjar, Viktor Zehe, Philipp Faustmann, Friedemann Zengerling, Axel John, and et al. 2021. "Grainyhead-Like 3 Influences Migration and Invasion of Urothelial Carcinoma Cells" International Journal of Molecular Sciences 22, no. 6: 2959. https://doi.org/10.3390/ijms22062959