
An Arabidopsis Oxalyl-CoA Decarboxylase, AtOXC, Is Important for Oxalate Catabolism in Plants

 ,
,
Abstract
:1. Introduction
2. Results
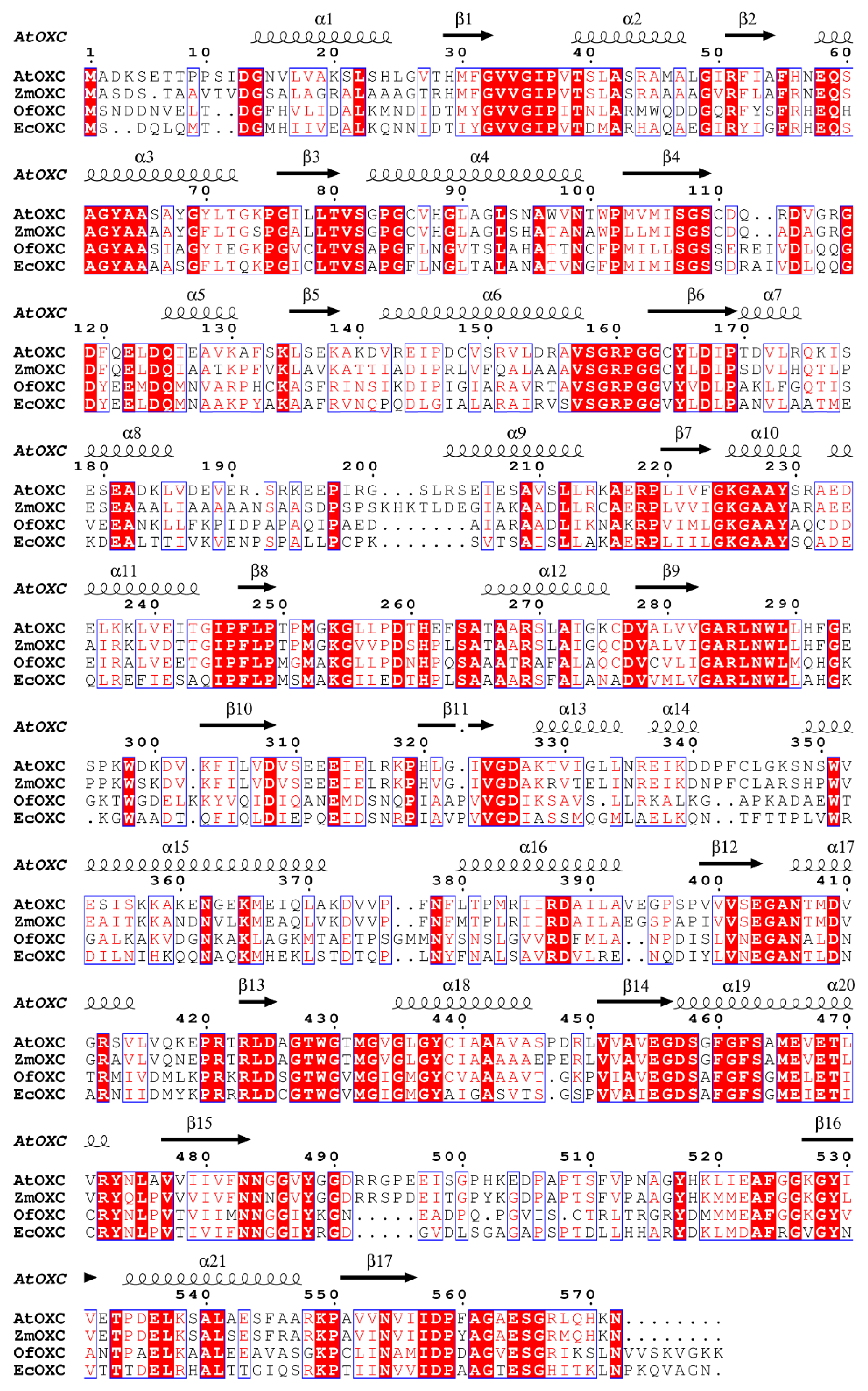
2.1. Identification and Cloning of an Arabidopsis OXC Homolog
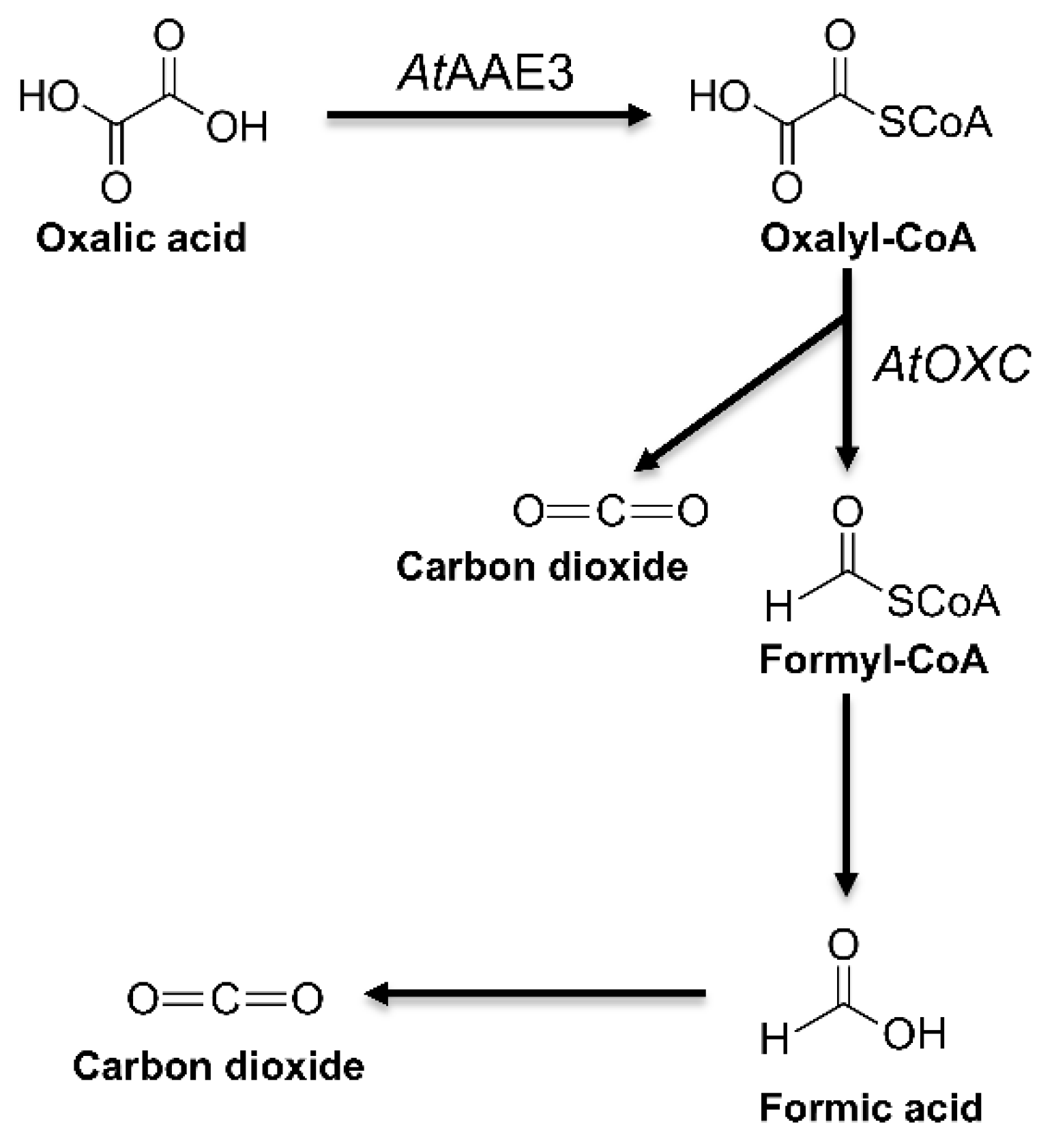
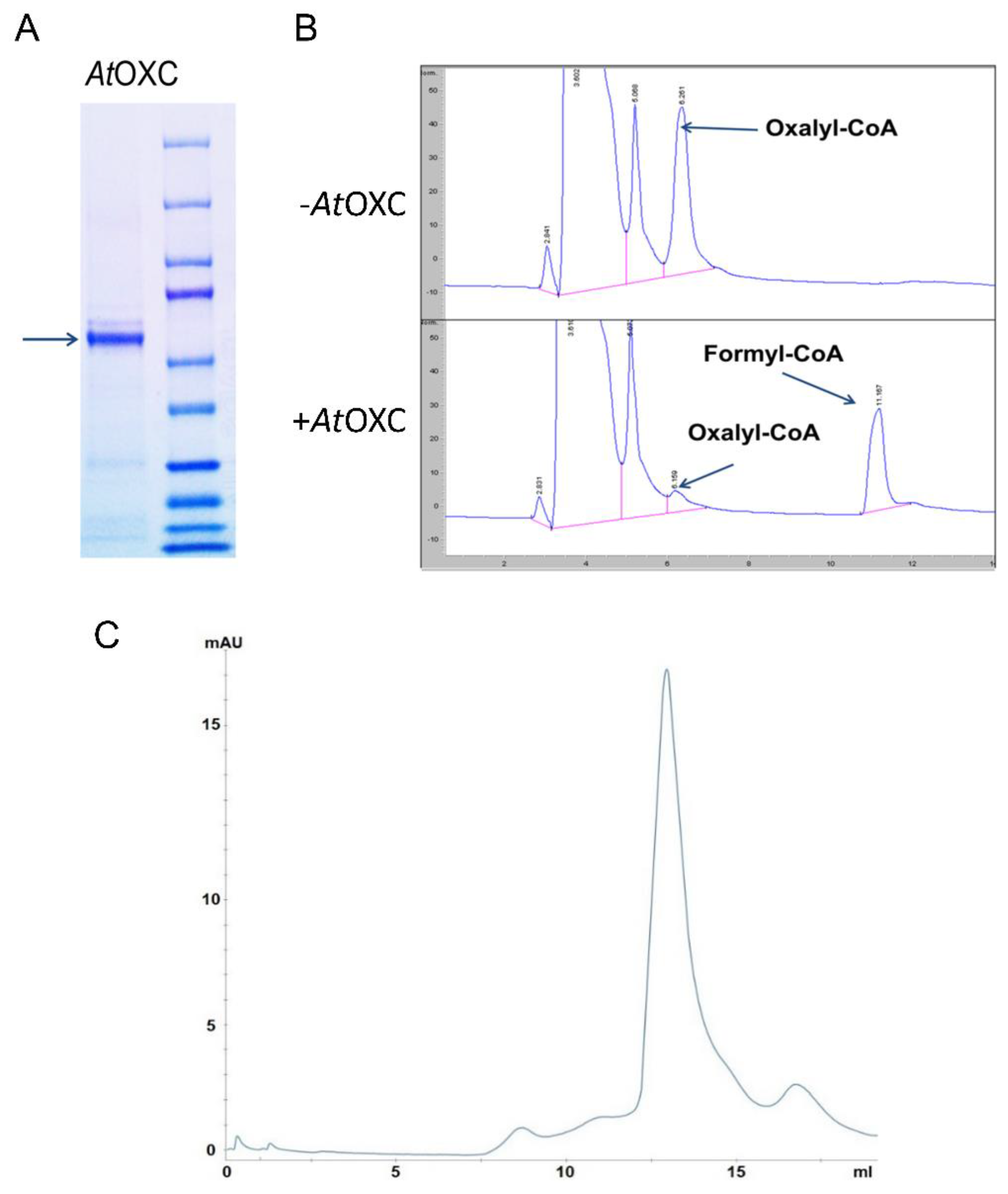
2.2. Arabidopsis OXC Possesses an Oxalyl-CoA Decarboxylase Activity
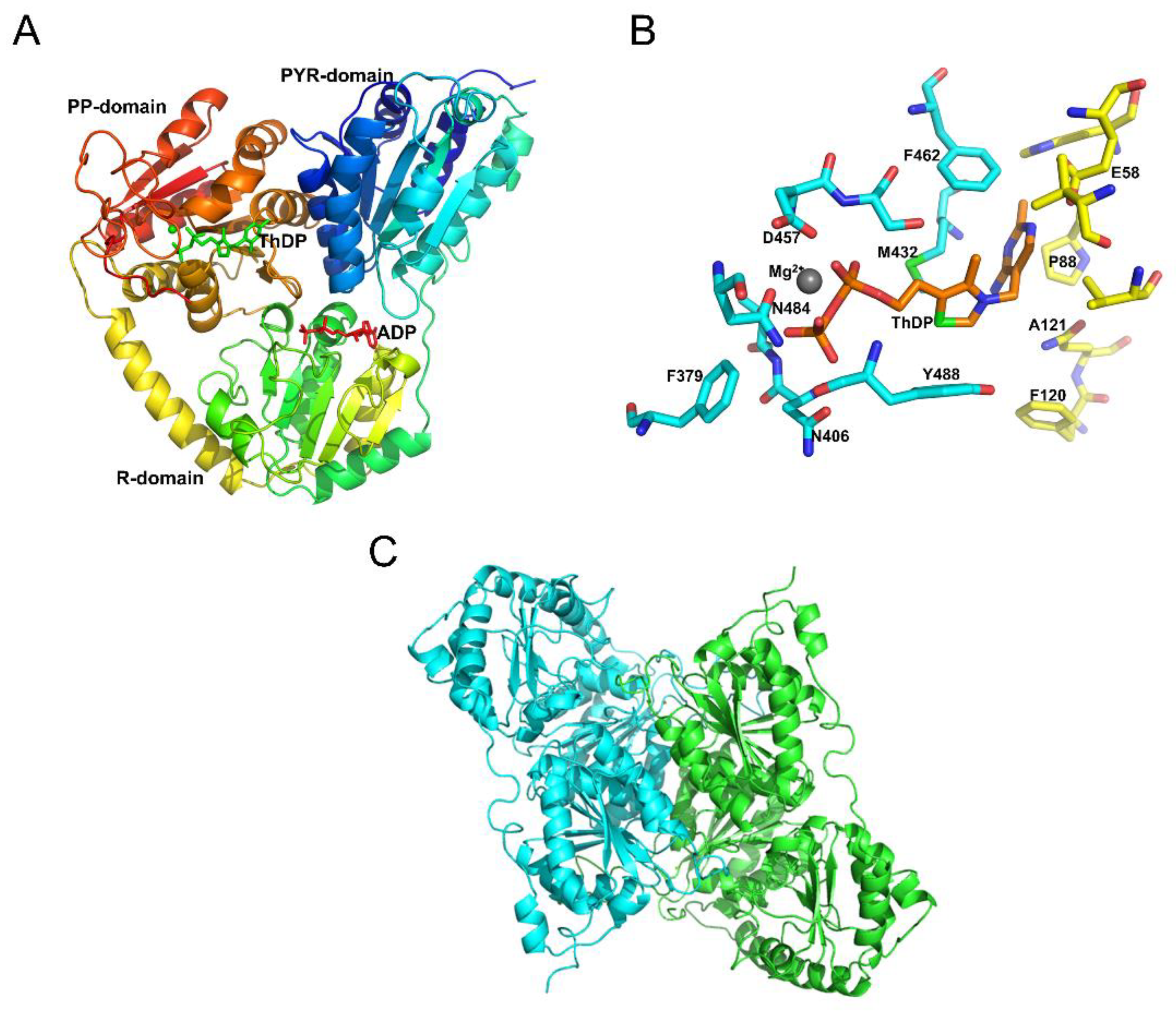
2.3. AtOXC Structural Model
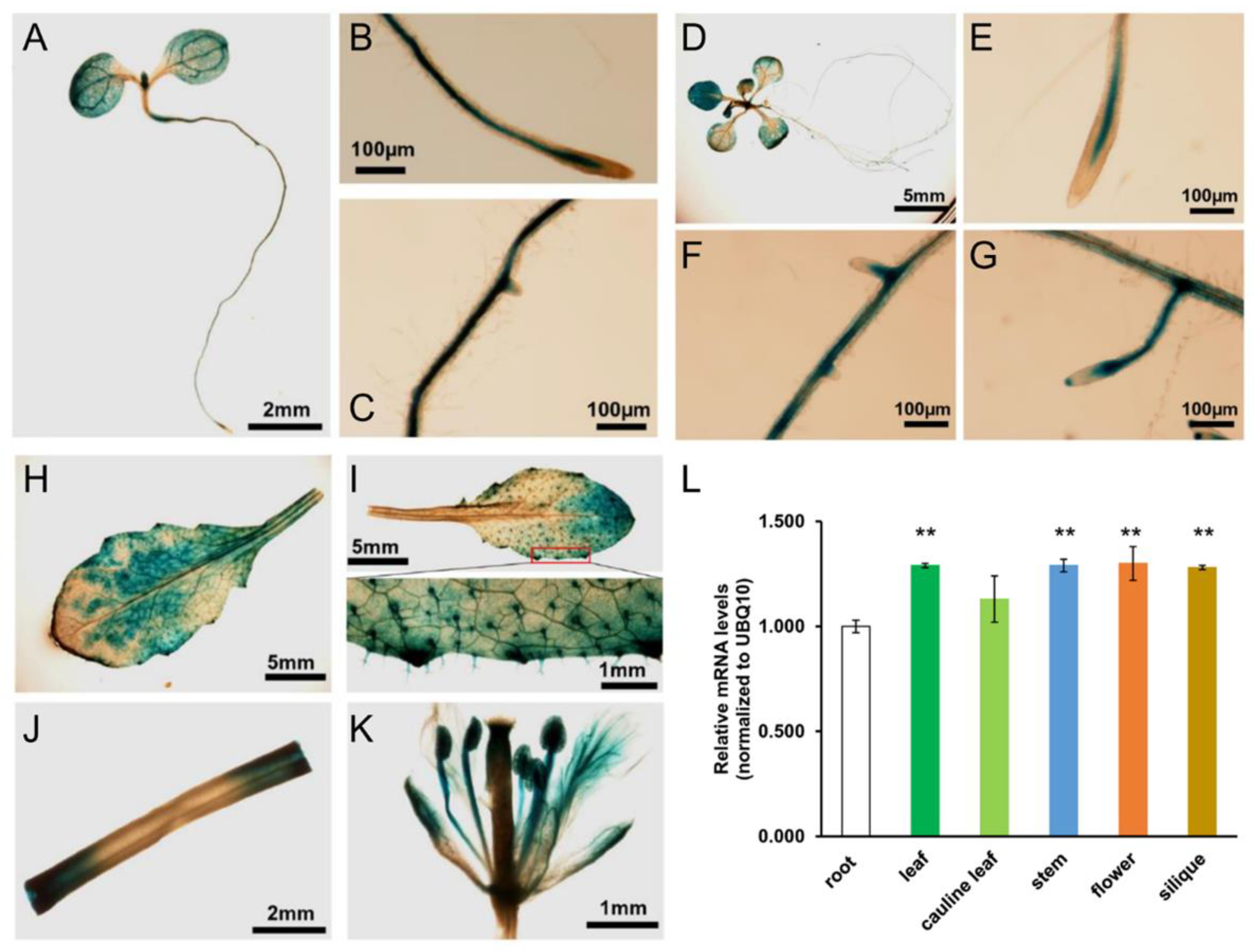
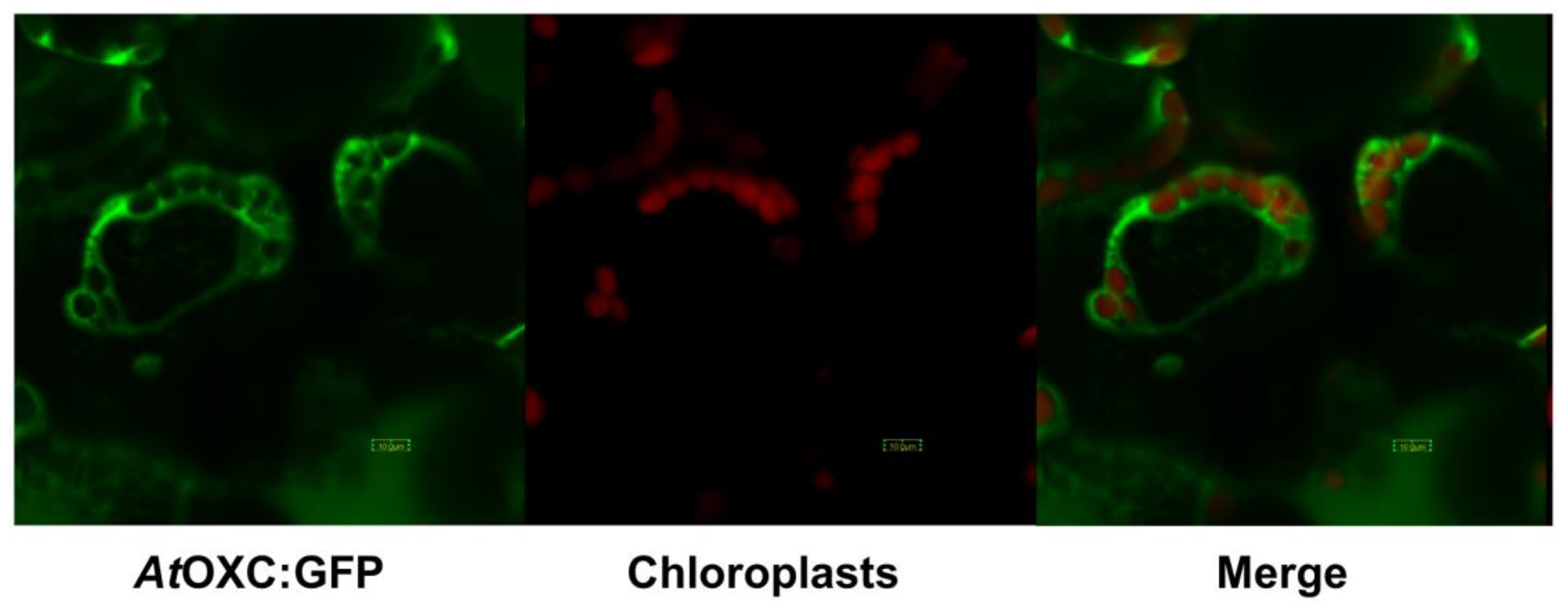
2.4. AtOXC Gene Expression and Protein Subcellular Localization
2.5. Deletion of AtOXC Leads to Embryonic Lethality
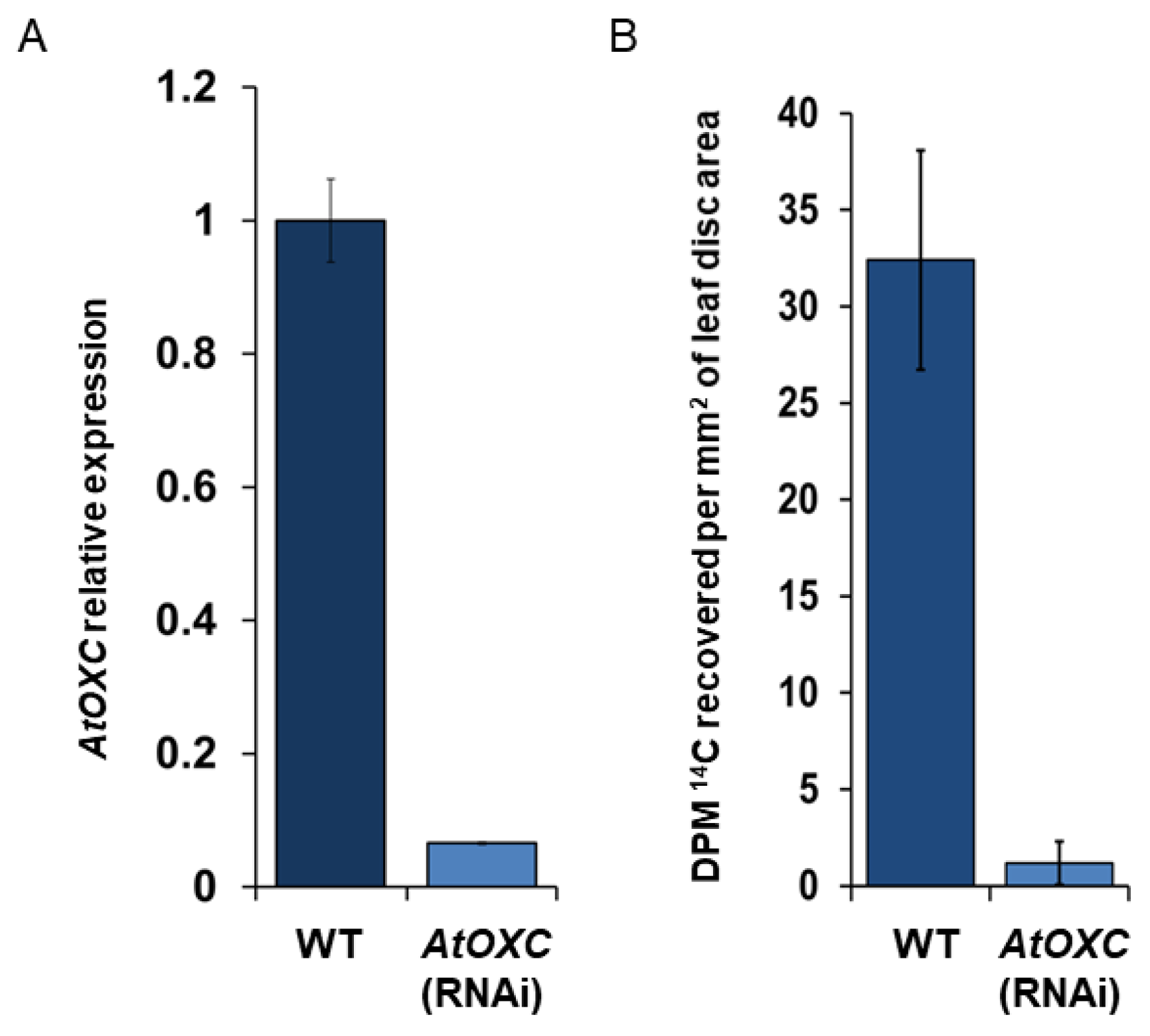
2.6. Reduction of AtOXC Impairs the Catabolism of Oxalate
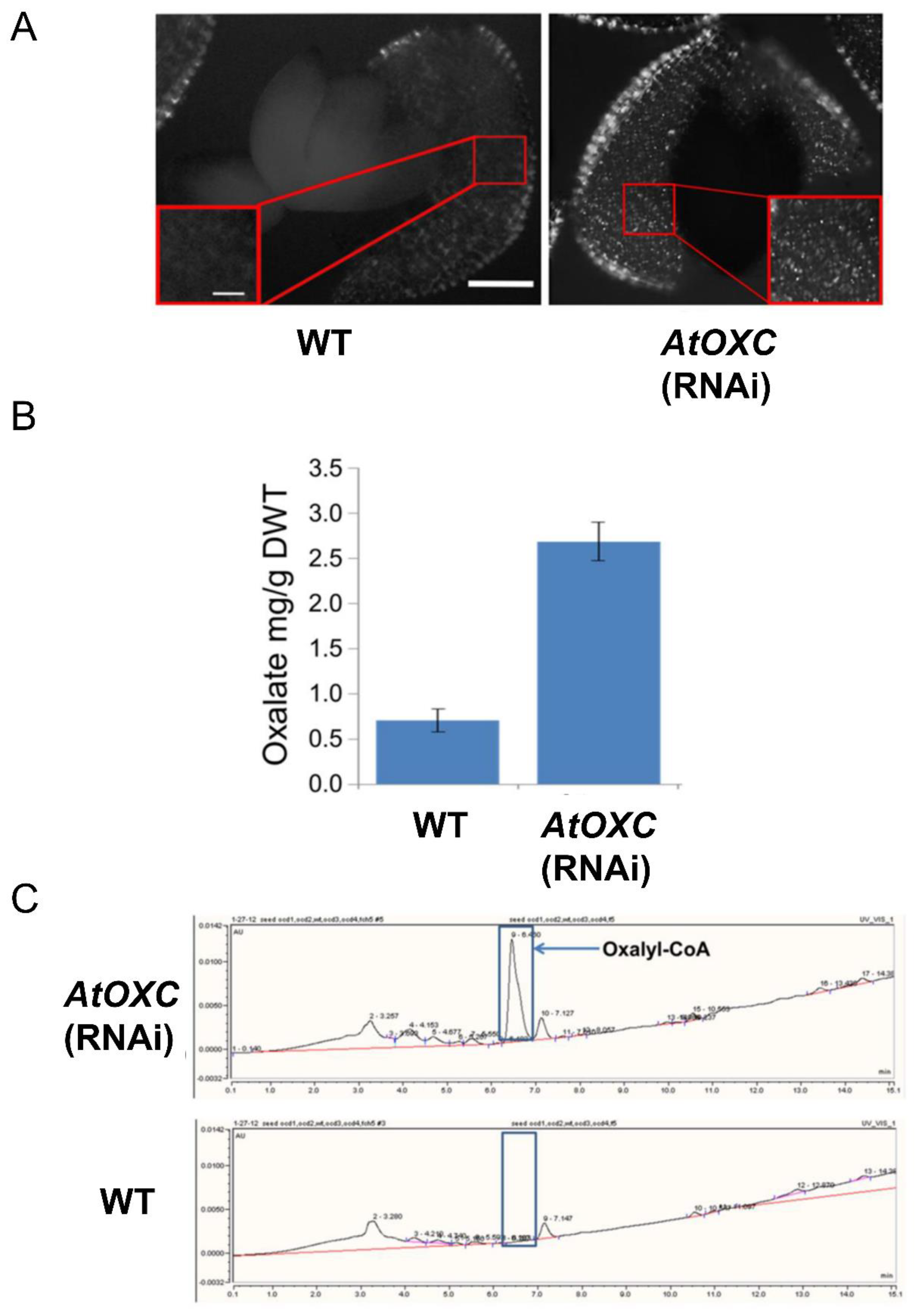
2.7. Reduction in AtOXC Results in the Accumulation of Oxalate and Oxalyl-CoA
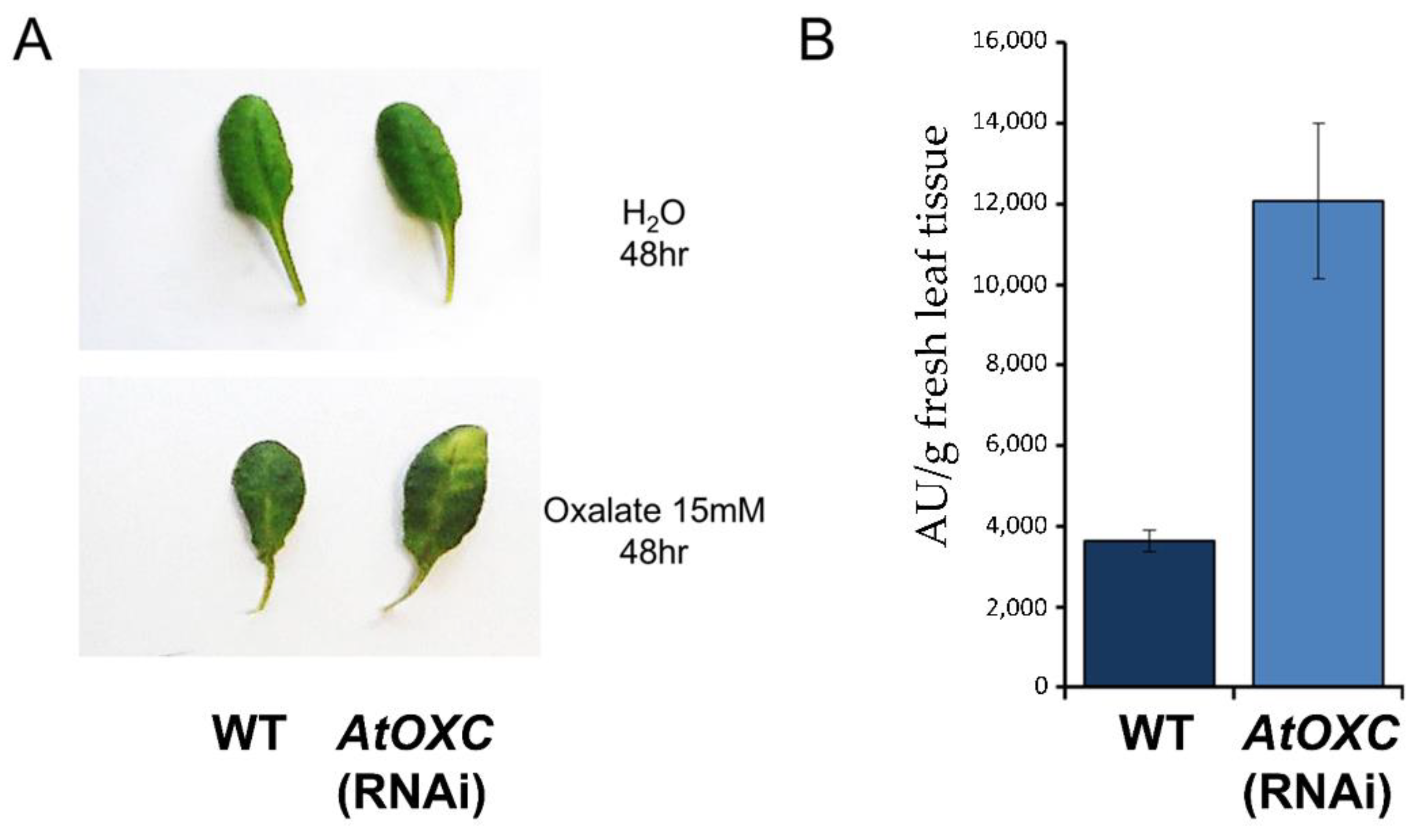
2.8. Reduction in AtOXC Results in an Increase in Sensitivity to Exogenous Oxalic Acid and Accumulation of Oxalyl-CoA
3. Discussion
4. Materials and Methods
4.1. AtOXC cDNA Isolation
4.2. OXC Alignment and Homology Modeling
4.3. His-Tagged AtOXC Recombinant Protein Purification
4.4. Size Exclusion Analysis of AtOXC
4.5. Assessment of AtOXC Activity
4.6. Generation of AtOXCp::Gus Lines and Histochemical Analysis
4.7. Plant Growth Conditions, RNA Isolation, cDNA Synthesis, and qRT-PCR Analysis
4.8. Subcellular Localization of AtOXC
4.9. AtOXC T-DNA Insertional Mutant and Generation of Atoxc Knock-Down Mutant
4.10. T-DNA Transmission Analysis
4.11. Radiolabeled Oxalate Feeding
4.12. Oxalate Sensitivity Assay
4.13. Microscopic Analysis of Calcium Oxalate Crystal Phenotype
4.14. Measurement of Seed Oxalate Concentrations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franceschi, V.R.; Nakata, P.A. Calcium Oxalate in Plants: Formation and Function. Annu. Rev. Plant Biol. 2005, 56, 41–71. [Google Scholar] [CrossRef]
- Nakata, P.A. Advances in our understanding of calcium oxalate crystal formation and function in plants. Plant Sci. 2003, 164, 901–909. [Google Scholar] [CrossRef]
- Nakata, P.A. Engineering Calcium Oxalate Crystal Formation in Arabidopsis. Plant Cell Physiol. 2012, 53, 1275–1282. [Google Scholar] [CrossRef] [Green Version]
- Scheid, C.; Koul, H.; Hill, W.A.; Luber-Narod, J.; Kennington, L.; Honeyman, T.; Jonassen, J.; Menon, M. Oxalate toxicity in LLC-PK1 cells: Role of free radicals. Kidney Int. 1996, 49, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Bateman, D.F.; Beer, S.V. Simultaneous Production and Synergistic Action of Oxalic Acid and Polygalacturonase during Pathogenesis by Sclerotium Rolfsii. Phytopathology 1965, 55, 204–211. [Google Scholar]
- Dong, X.; Ji, R.; Guo, X.; Foster, S.J.; Chen, H.; Dong, C.; Liu, Y.; Hu, Q.; Liu, S. Expressing a gene encoding wheat oxalate oxidase enhances resistance to Sclerotinia sclerotiorum in oilseed rape (Brassica napus). Planta 2008, 228, 331–340. [Google Scholar] [CrossRef]
- Guimarães, R.L.; Stotz, H.U. Oxalate Production by Sclerotinia sclerotiorum Deregulates Guard Cells during Infection. Plant Physiol. 2004, 136, 3703–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lumsden, R.D. Pectolytic enzymes of Sclerotinia sclerotiorum and their localization in infected bean. Can. J. Bot. 1976, 54, 2630–2641. [Google Scholar] [CrossRef]
- Hegedus, D.D.; Rimmer, S.R. Sclerotinia sclerotiorum: When “to be or not to be” a pathogen? FEMS Microbiol. Lett. 2005, 251, 177–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.S.; Min, J.-Y.; Dickman, M.B. Oxalic Acid Is an Elicitor of Plant Programmed Cell Death during Sclerotinia sclerotiorum Disease Development. Mol. Plant-Microbe Interact. 2008, 21, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Dutton, M.V.; Evans, C.S. Oxalate production by fungi: Its role in pathogenicity and ecology in the soil environment. Can. J. Microbiol. 1996, 42, 881–895. [Google Scholar] [CrossRef]
- Williams, B.; Kabbage, M.; Kim, H.-J.; Britt, R.; Dickman, M.B. Tipping the Balance: Sclerotinia sclerotiorum Secreted Oxalic Acid Suppresses Host Defenses by Manipulating the Host Redox Environment. PLoS Pathog. 2011, 7, e1002107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschi, V.R.; Loewus, F.A. Oxalate biosynthesis and function in plants and fungi. In Calcium Oxalate in Biological Systems; Khan, S.R., Ed.; CRC Press: Boca Raton, FL, USA, 1995; pp. 113–130. [Google Scholar]
- Lane, B.G. Oxalate, Germins, and Higher-Plant Pathogens. IUBMB Life 2002, 53, 67–75. [Google Scholar] [CrossRef]
- Svedružić, D.; Jonsson, S.; Toyota, C.G.; Reinhardt, L.A.; Ricagno, S.; Lindqvist, Y.; Richards, N.G. The enzymes of oxalate metabolism: Unexpected structures and mechanisms. Arch. Biochem. Biophys. 2005, 433, 176–192. [Google Scholar] [CrossRef] [PubMed]
- Lane, B.; Dunwell, J.; Ray, J.; Schmitt, M.; Cuming, A. Germin, a protein marker of early plant development, is an oxalate oxidase. J. Biol. Chem. 1993, 268, 12239–12242. [Google Scholar] [CrossRef]
- Druka, A.; Kudrna, D.; Kannangara, C.G.; Von Wettstein, D.; Kleinhofs, A. Physical and genetic mapping of barley (Hordeum vulgare) germin-like cDNAs. Proc. Natl. Acad. Sci. USA 2002, 99, 850–855. [Google Scholar] [CrossRef] [Green Version]
- Membré, N.; Berna, A.; Neutelings, G.; David, A.; David, H.; Staiger, R.; Vásquez, J.S.; Raynal, M.; Delseny, M.; Bernier, F. cDNA sequence, genomic organization and differential expression of three Arabidopsis genes for germin/oxalate oxidase-like proteins. Plant Mol. Biol. 1997, 35, 459–469. [Google Scholar] [CrossRef]
- Membré, N.; Bernier, F.; Staiger, D.; Berna, A. Arabidopsis thaliana germin-like proteins: Common and specific features point to a variety of functions. Planta 2000, 211, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Giovanelli, J.; Tobin, N.F. Adenosine Triphosphate- and Coenzyme A-dependent Decarboxylation of Oxalate by Extracts of Peas. Nat. Cell Biol. 1961, 190, 1006–1007. [Google Scholar] [CrossRef]
- Foster, J.; Kim, H.U.; Nakata, P.A.; Browse, J. A Previously Unknown Oxalyl-CoA Synthetase Is Important for Oxalate Catabolism in Arabidopsis. Plant Cell 2012, 24, 1217–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shockey, J.M.; Fulda, M.S.; Browse, J. Arabidopsis Contains a Large Superfamily of Acyl-Activating Enzymes. Phylogenetic and Biochemical Analysis Reveals a New Class of Acyl-Coenzyme A Synthetases. Plant Physiol. 2003, 132, 1065–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shockey, J.; Browse, J. Genome-level and biochemical diversity of the acyl-activating enzyme superfamily in plants. Plant J. 2011, 66, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.; Luo, B.; Nakata, P.A. An Oxalyl-CoA Dependent Pathway of Oxalate Catabolism Plays a Role in Regulating Calcium Oxalate Crystal Accumulation and Defending against Oxalate-Secreting Phytopathogens in Medicago truncatula. PLoS ONE 2016, 11, e0149850. [Google Scholar] [CrossRef] [Green Version]
- Lou, H.Q.; Fan, W.; Xu, J.M.; Gong, Y.L.; Jin, J.F.; Chen, W.W.; Liu, L.Y.; Hai, M.R.; Yang, J.L.; Zheng, S.J. An Oxalyl-CoA Synthetase Is Involved in Oxalate Degradation and Aluminum Tolerance. Plant Physiol. 2016, 172, 1679–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, J.; Nakata, P.A. An oxalyl-CoA synthetase is important for oxalate metabolism inSaccharomyces cerevisiae. FEBS Lett. 2014, 588, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Fu, M.; Ji, C.; Huang, Y.; Wu, Y. Maize Oxalyl-CoA Decarboxylase1 Degrades Oxalate and Affects the Seed Metabolome and Nutritional Quality. Plant Cell 2018, 30, 2447–2462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obayashi, T.; Hayashi, S.; Saeki, M.; Ohta, H.; Kinoshita, K. ATTED-II provides coexpressed gene networks for Arabidopsis. Nucleic Acids Res. 2008, 37, D987–D991. [Google Scholar] [CrossRef] [Green Version]
- Toufighi, K.; Brady, S.M.; Austin, R.; Ly, E.; Provart, N.J. The Botany Array Resource: E-Northerns, Expression Angling, and promoter analyses. Plant J. 2005, 43, 153–163. [Google Scholar] [CrossRef]
- Lung, H.Y.; Baetz, A.L.; Peck, A.B. Molecular cloning, DNA sequence, and gene expression of the oxalyl-coenzyme A decarboxylase gene, oxc, from the bacterium Oxalobacter formigenes. J. Bacteriol. 1994, 176, 2468–2472. [Google Scholar] [CrossRef] [Green Version]
- Makkapati, S.; D’Agati, V.D.; Balsam, L. “Green Smoothie Cleanse” Causing Acute Oxalate Nephropathy. Am. J. Kidney Dis. 2018, 71, 281–286. [Google Scholar] [CrossRef]
- Cheng, N.; Foster, J.; Mysore, K.S.; Wen, J.; Rao, X.; Nakata, P.A. Effect of Acyl Activating Enzyme (AAE) 3 on the growth and development of Medicago truncatula. Biochem. Biophys. Res. Commun. 2018, 505, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Baetz, A.L.; Allison, M.J. Purification and characterization of oxalyl-coenzyme A decarboxylase from Oxalobacter formigenes. J. Bacteriol. 1989, 171, 2605–2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.S.; Duncan, S.H.; Cave, D.R. Oxalobacter formigenes and its role in oxalate metabolism in the human gut. FEMS Microbiol. Lett. 2004, 230, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Coté, G.G.; Gibernau, M. Distribution of calcium oxalate crystals in floral organs of Araceae in relation to pollination strategy. Am. J. Bot. 2012, 99, 1231–1242. [Google Scholar] [CrossRef]
- Gębura, J.; Winiarczyk, K. A study on calcium oxalate crystals in Tinantia anomala (Commelinaceae) with special reference to ultrastructural changes during anther development. J. Plant Res. 2016, 129, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Paiva, E.A.S. Are calcium oxalate crystals a dynamic calcium store in plants? New Phytol. 2019, 223, 1707–1711. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, N.; Ghosh, R.; Ghosh, S.; Narula, K.; Tayal, R.; Datta, A.; Chakraborty, S. Reduction of Oxalate Levels in Tomato Fruit and Consequent Metabolic Remodeling Following Overexpression of a Fungal Oxalate Decarboxylase. Plant Physiol. 2013, 162, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Chattopadhyay, A.; Ghosh, S.; Irfan, M.; Chakraborty, N.; Chakraborty, S.; Datta, A. Improving nutritional quality and fungal tolerance in soya bean and grass pea by expressing an oxalate decarboxylase. Plant Biotechnol. J. 2016, 14, 1394–1405. [Google Scholar] [CrossRef]
- Chenna, R. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef] [Green Version]
- Gouet, P.; Courcelle, E. ENDscript: A workflow to display sequence and structure information. Bioinformatics 2002, 18, 767–768. [Google Scholar] [CrossRef] [Green Version]
- Fiser, A.; Šali, A. Modeller: Generation and Refinement of Homology-Based Protein Structure Models. Methods Enzymol. 2003, 374, 461–491. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Demoz, A.; Garras, A.; Asiedu, D.K.; Netteland, B.; Berge, R.K. Rapid method for the separation and detection of tissue short-chain coenzyme A esters by reversed-phase high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1995, 667, 148–152. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Cheng, N.-H. AtGRX4, an Arabidopsis chloroplastic monothiol glutaredoxin, is able to suppress yeast grx5 mutant phenotypes and respond to oxidative stress. FEBS Lett. 2008, 582, 848–854. [Google Scholar] [CrossRef] [Green Version]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY™ vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Alonso, J.M.; Stepanova, A.N.; Leisse, T.J.; Kim, C.J.; Chen, H.; Shinn, P.; Stevenson, D.K.; Zimmerman, J.; Barajas, P.; Cheuk, R.; et al. Genome-Wide Insertional Mutagenesis of Arabidopsis thaliana. Science 2003, 301, 653–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansen, L.K.; Carrington, J.C. Silencing on the Spot. Induction and Suppression of RNA Silencing in the Agrobacterium-Mediated Transient Expression System. Plant Physiol. 2001, 126, 930–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crossing | Resistant | Sensitive | Ratio | Transmission Efficiency (%) |
|---|---|---|---|---|
| atoxc-2 self-fertilized | 228 (BarR) | 233 (BarS) | 0.979 | / |
| atoxc-2 (♀) × wild type (♂) | 100 (BarR) | 107 (BarS) | 0.935 | 93.5 |
| wild type (♀) × atoxc-2 (♂) | 1 (BarR) | 76 (BarS) | 0.013 | 1.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foster, J.; Cheng, N.; Paris, V.; Wang, L.; Wang, J.; Wang, X.; Nakata, P.A. An Arabidopsis Oxalyl-CoA Decarboxylase, AtOXC, Is Important for Oxalate Catabolism in Plants. Int. J. Mol. Sci. 2021, 22, 3266. https://doi.org/10.3390/ijms22063266
Foster J, Cheng N, Paris V, Wang L, Wang J, Wang X, Nakata PA. An Arabidopsis Oxalyl-CoA Decarboxylase, AtOXC, Is Important for Oxalate Catabolism in Plants. International Journal of Molecular Sciences. 2021; 22(6):3266. https://doi.org/10.3390/ijms22063266
Chicago/Turabian StyleFoster, Justin, Ninghui Cheng, Vincent Paris, Lingfei Wang, Jin Wang, Xiaoqiang Wang, and Paul A. Nakata. 2021. "An Arabidopsis Oxalyl-CoA Decarboxylase, AtOXC, Is Important for Oxalate Catabolism in Plants" International Journal of Molecular Sciences 22, no. 6: 3266. https://doi.org/10.3390/ijms22063266
APA StyleFoster, J., Cheng, N., Paris, V., Wang, L., Wang, J., Wang, X., & Nakata, P. A. (2021). An Arabidopsis Oxalyl-CoA Decarboxylase, AtOXC, Is Important for Oxalate Catabolism in Plants. International Journal of Molecular Sciences, 22(6), 3266. https://doi.org/10.3390/ijms22063266