
Late Health Effects of Partial Body Irradiation Injury in a Minipig Model Are Associated with Changes in Systemic and Cardiac IGF-1 Signaling

 ,
,
Abstract
:1. Introduction
2. Results
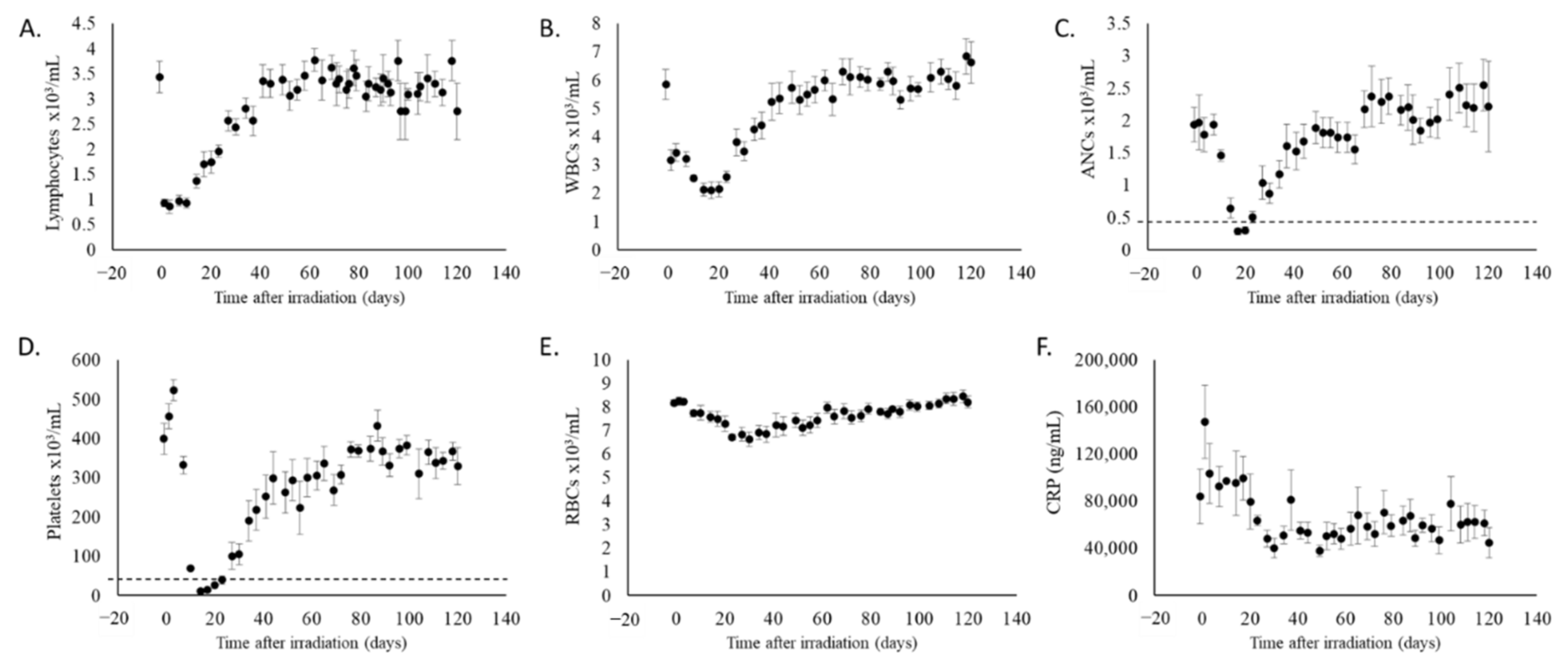
2.1. Partial Body Irradiation Allowed for Recovery from Hematopoietic Cell Loss and Induction of Inflammation
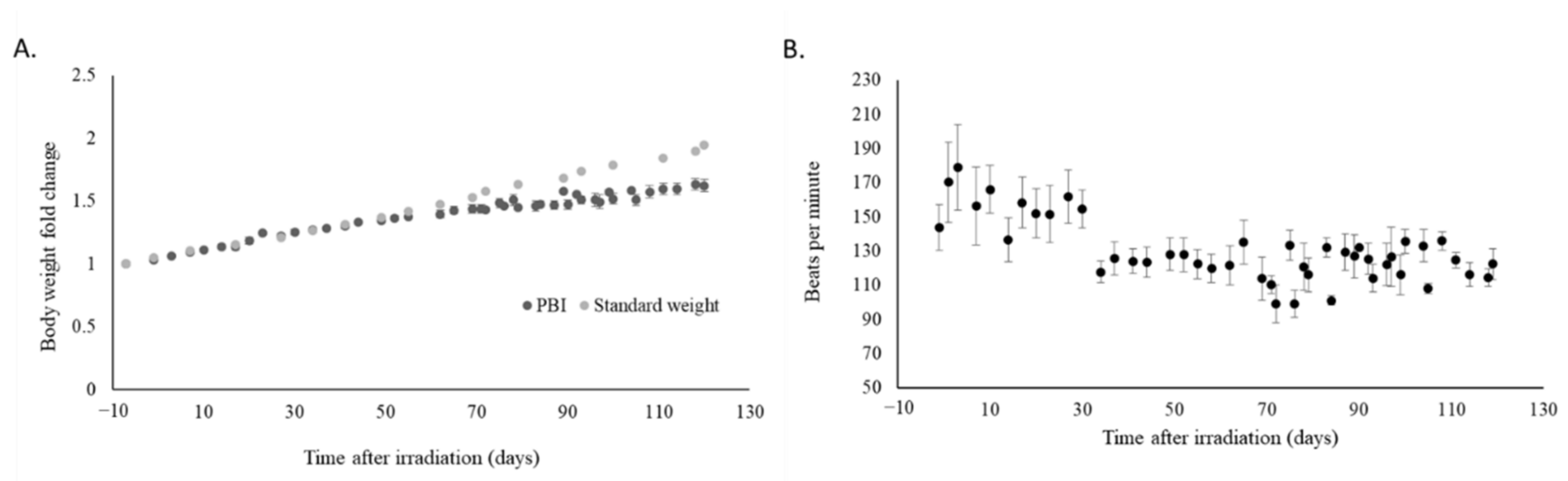
2.2. Irradiation Slowed Weight Gain and Decreased Heart Rate
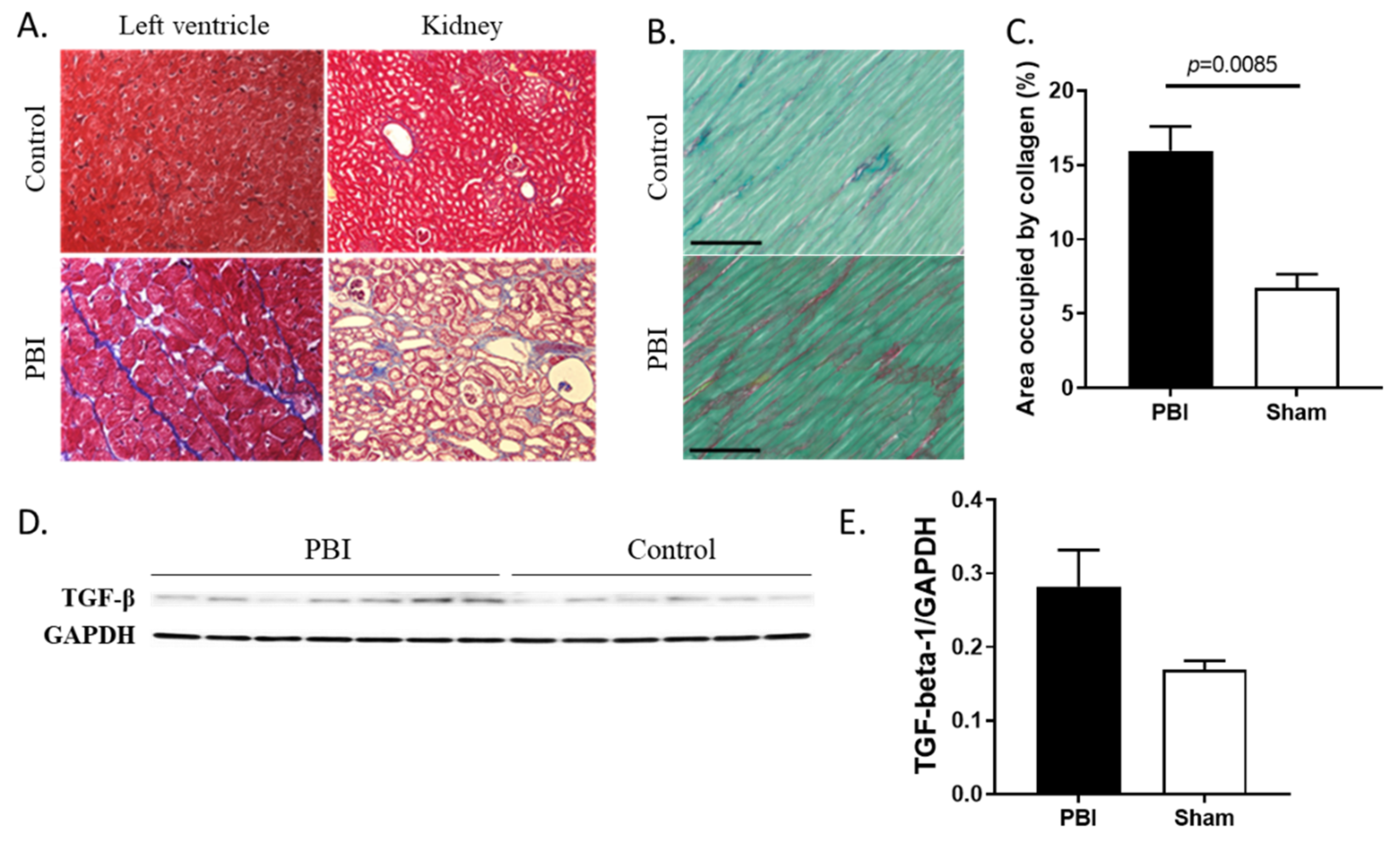
2.3. Organ Damage and Fibrosis Were Induced by Radiation
2.4. Collagen Deposition Increased in Heart and Kidneys of Irradiated Animals
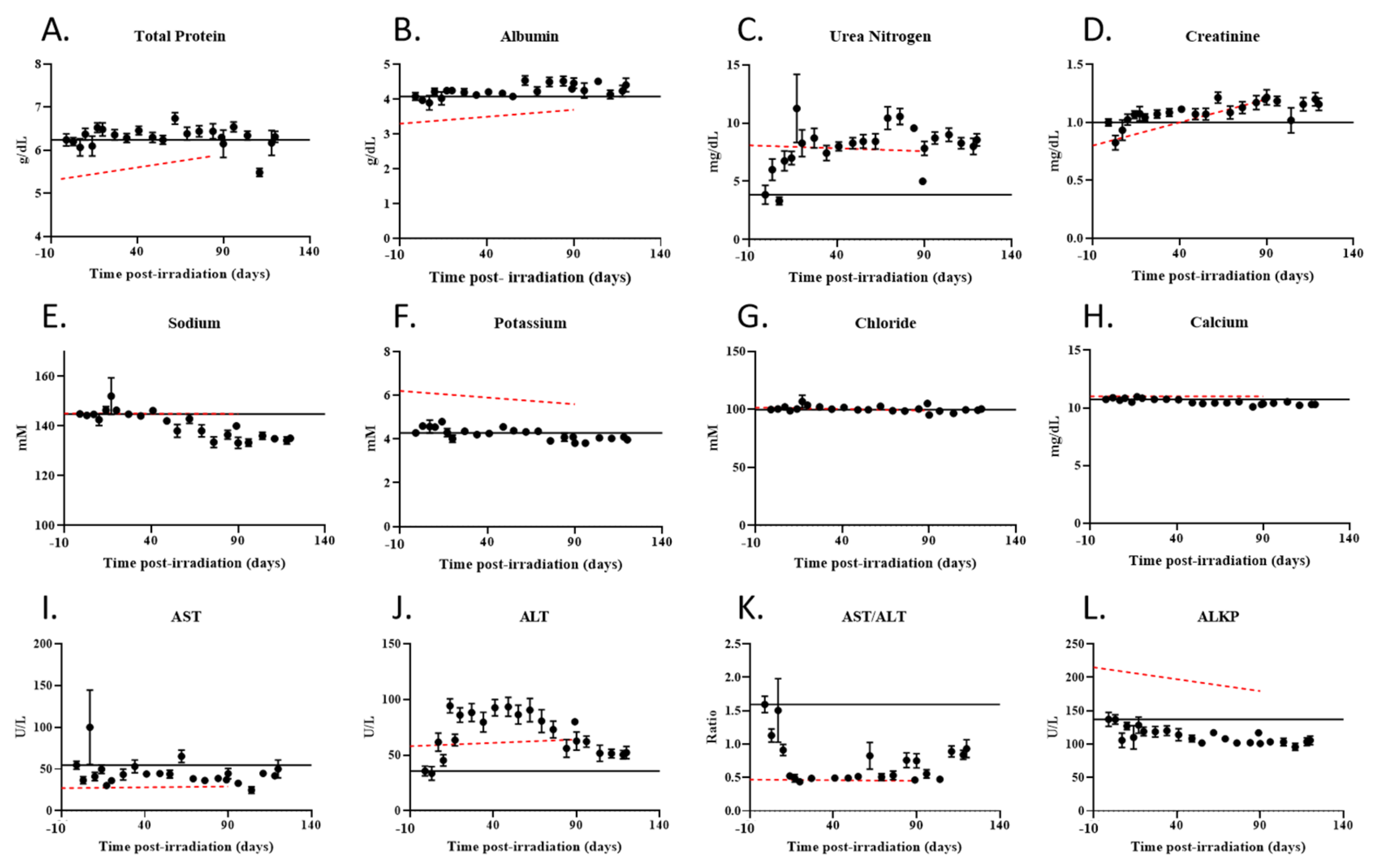
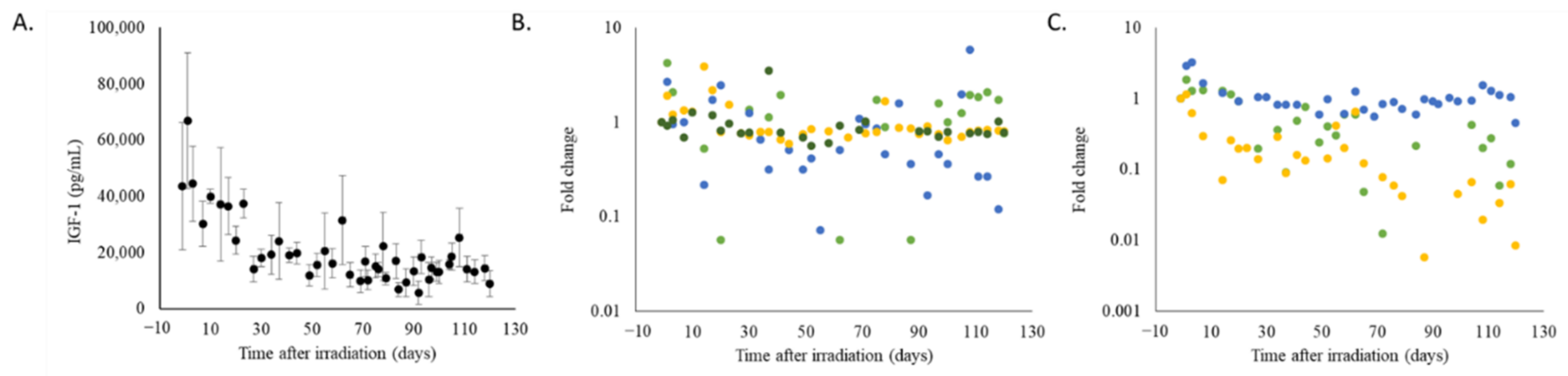
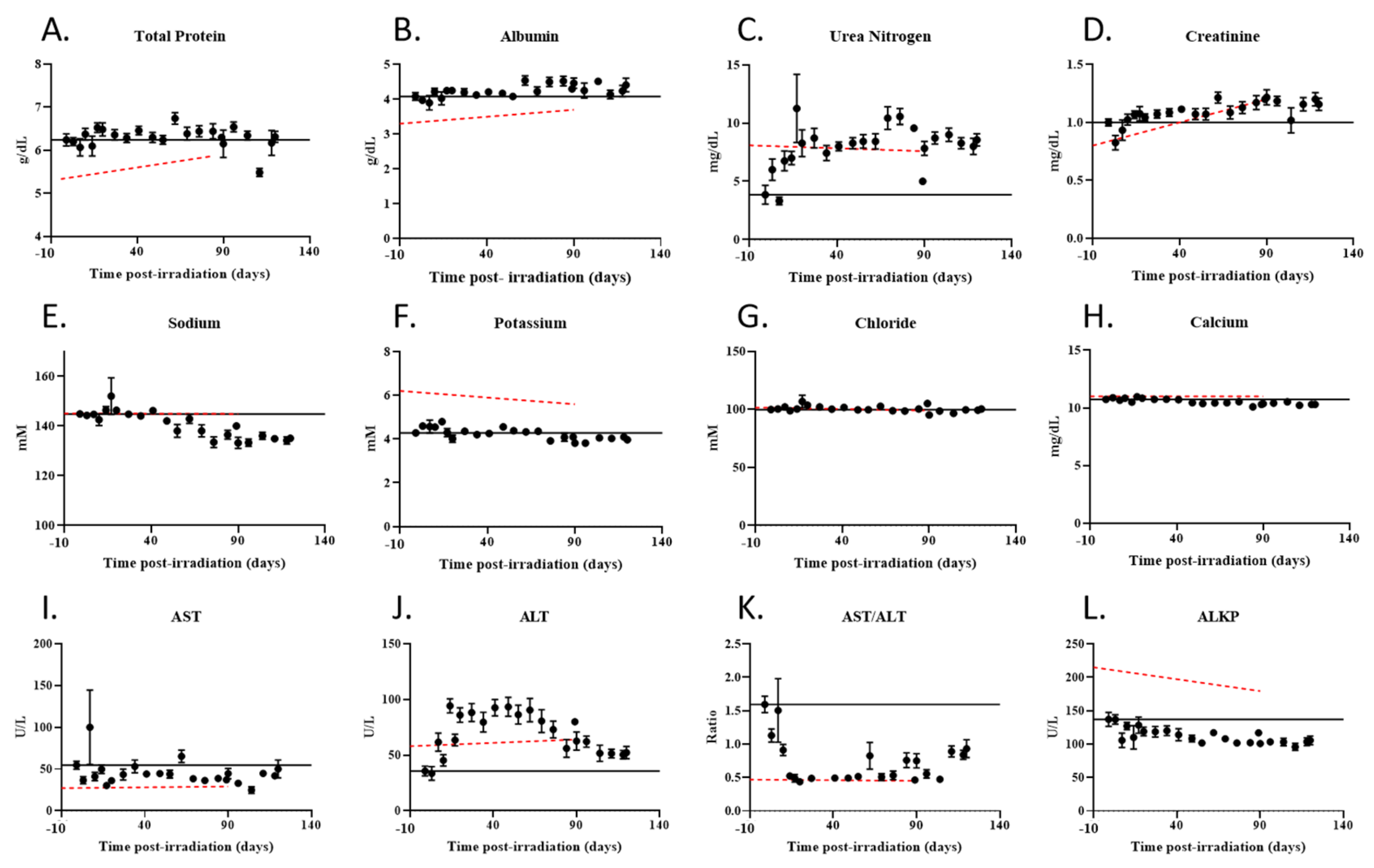
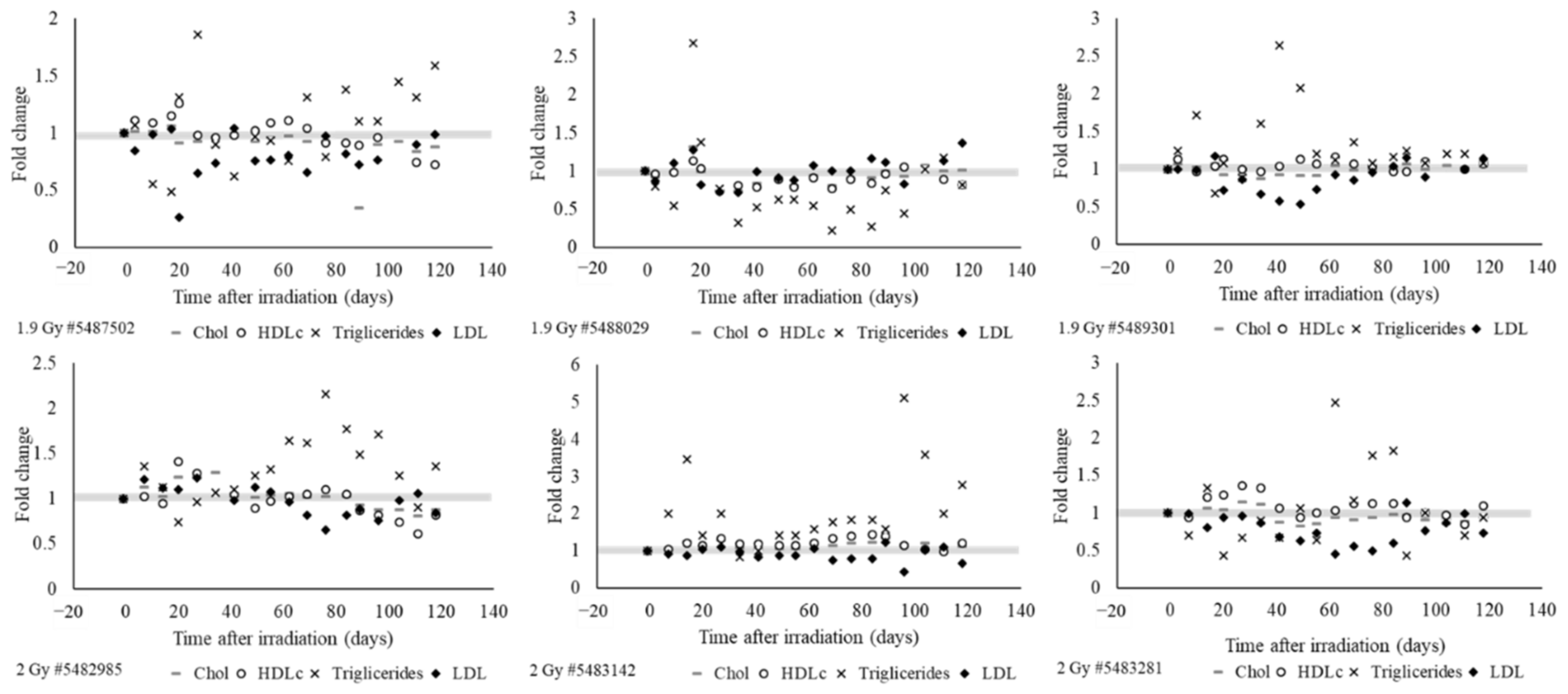
2.5. Radiation Exposured Altered Plasma IGF-1 Levels, Lipid Profiles, and Liver and Kidney Function
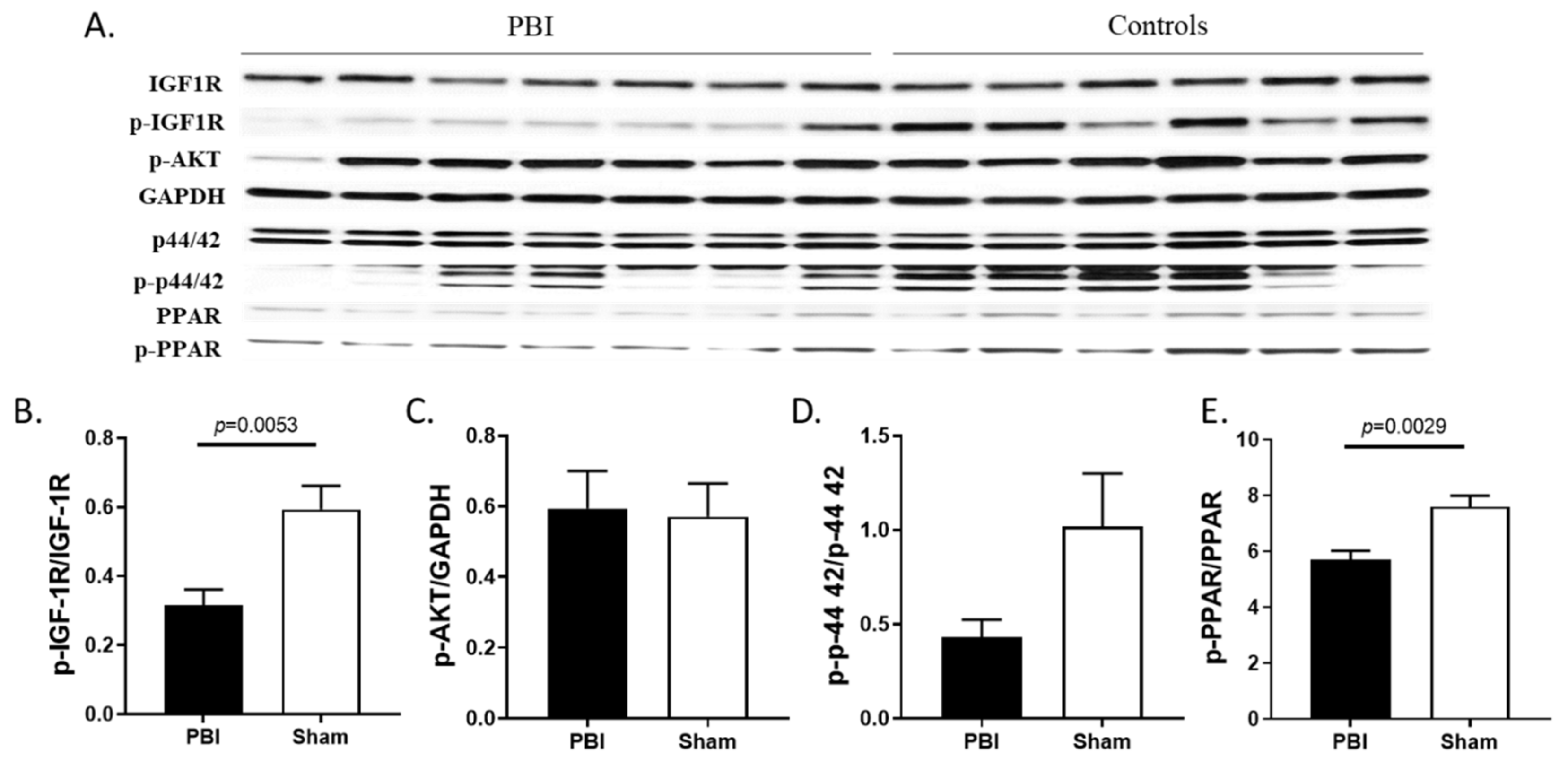
2.6. Radiation Blunted the Cardiac IGF-1 Signaling Pathway
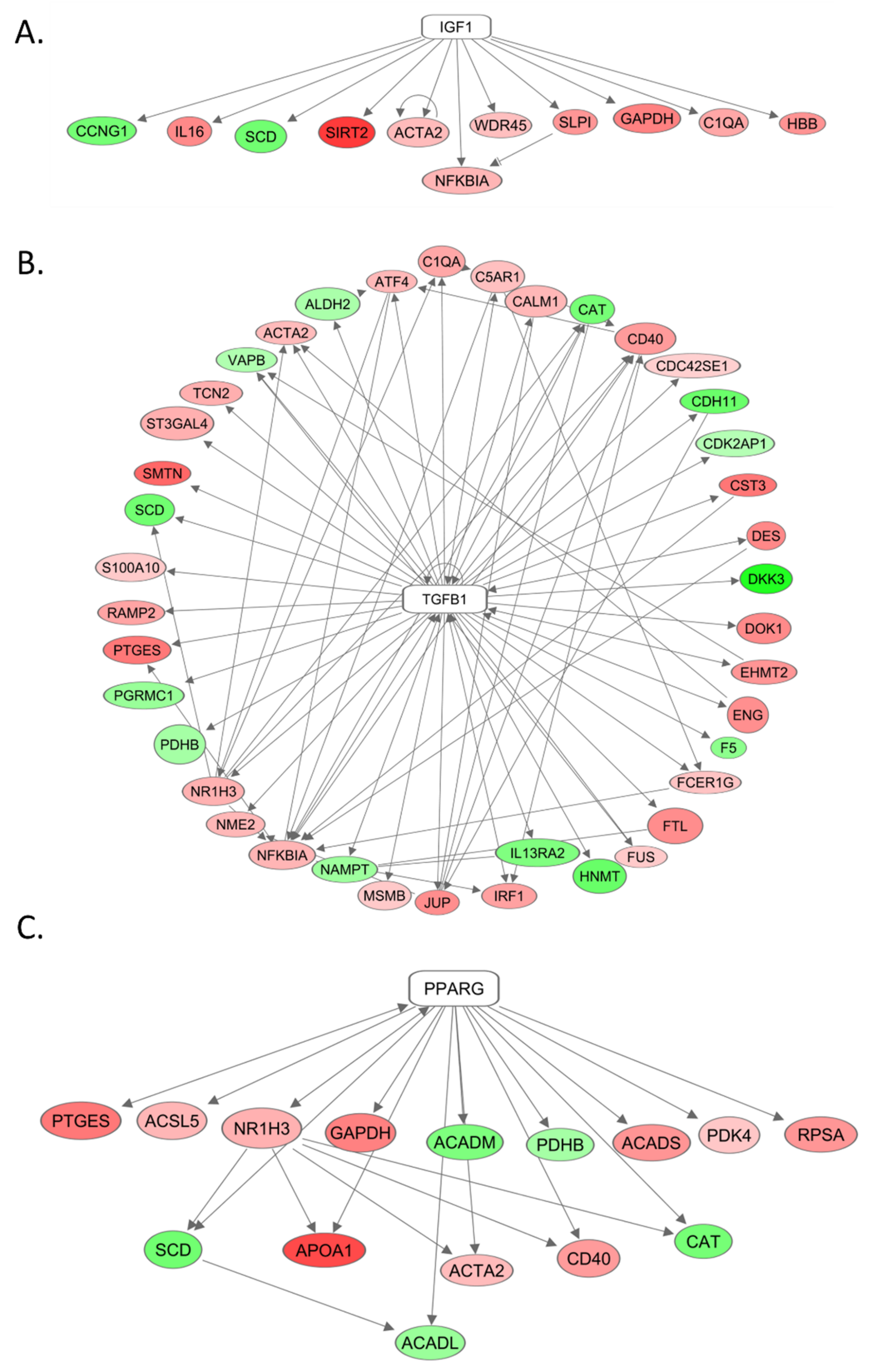
2.7. The Role of IGF-1, TGF-beta, and PPAR-gamma in Differential Gene Expression as Seen by mRNA Sequencing
3. Discussion
Conclusions and Limitations
4. Materials and Methods
4.1. Animal Care and Irradiation
4.2. Blood Sampling and Plasma and Tissue Collection
4.3. Histology
4.4. Preparation of Cytosolic and Membrane Fractions and Immunoblotting
4.5. Determination of Plasma IGF-1 and C-Reactive Protein (CRP)
4.6. Collagen Staining and Measurement
4.7. Sample Preparation and mRNA Sequencing
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IGF-1 | Insulin-like growth factor-1 |
| GH | Growth hormone |
| PBI | Partial body irradiation |
| CRP | C-reactive protein |
| MV | Megavolt |
| LINAC | Linear accelerator |
| TGF-beta | Transforming growth factor beta |
| AST | Aspartate transaminase |
| ALT | Alanine transferase |
| HDL | High-density lipoprotein |
| LDL | Low-density lipoprotein |
| IGF-1R | Insulin-like growth factor-1 receptor |
| MAPK | MAP kinase |
| PPAR | Peroxisome proliferator-activated receptor |
| H-ARS | Hematopoietic Acute Radiation Syndrome, |
| GI-ARS | Gastrointestinal Acute Radiation Syndrome |
| mRNA | Messenger Ribonucleic Acid |
| URA | Upstream Regulator Analysis |
| IPA | Ingenuity Pathway Analysis |
| DEG | Differentially expressed genes |
| sem | Standard Error of Mean |
References
- Douple, E.B.; Mabuchi, K.; Cullings, H.M.; Preston, D.L.; Kodama, K.; Shimizu, Y.; Fujiwara, S.; Shore, R.E. Long-term radiation-related health effects in a unique human population: Lessons learned from the atomic bomb survivors of Hiroshima and Nagasaki. Disaster Med. Public Health Prep. 2011, 5 (Suppl. S1), S122–S133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armanious, M.A.; Mohammadi, H.; Khodor, S.; Oliver, D.E.; Johnstone, P.A.; Fradley, M.G. Cardiovascular effects of radiation therapy. Curr. Probl. Cancer 2018, 42, 433–442. [Google Scholar] [CrossRef]
- Bazyka, D.; Prysyazhnyuk, A.; Gudzenko, N.; Dyagil, I.; Belyi, D.; Chumak, V.; Buzunov, V. Epidemiology of Late Health Effects in Ukrainian Chornobyl Cleanup Workers. Health Phys. 2018, 115, 161–169. [Google Scholar] [CrossRef]
- Niazi, A.K.; Niazi, S.K. Endocrine effects of Fukushima: Radiation-induced endocrinopathy. Indian J. Endocrinol. Metab. 2011, 15, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Rose, S.R.; Horne, V.E.; Howell, J.; Lawson, S.A.; Rutter, M.M.; Trotman, G.E.; Corathers, S.D. Late endocrine effects of childhood cancer. Nat. Rev. Endocrinol. 2016, 12, 319–336. [Google Scholar] [CrossRef]
- Chemaitilly, W.; Cohen, L.E.; Mostoufi-Moab, S.; Patterson, B.C.; Simmons, J.H.; Meacham, L.R.; van Santen, H.M.; Sklar, C.A. Endocrine Late Effects in Childhood Cancer Survivors. J. Clin. Oncol. 2018, 36, 2153–2159. [Google Scholar] [CrossRef]
- Ito, C. Trends in the prevalence of diabetes mellitus among Hiroshima atomic bomb survivors. Diabetes Res. Clin. Pract. 1994, 24, S29–S35. [Google Scholar] [CrossRef]
- Zueva, N.A.; Kovalenko, A.N.; Gerasimenko, T.I.; Man’kovskii, B.N.; Korpachova, T.I.; Efimov, A.S. Analysis of irradiation dose, body mass index and insulin blood concentration in personnel cleaning up after the Chernobyl nuclear plant accident. Likars’ka Sprava 2001, 4, 26–28. [Google Scholar]
- Narendran, N.; Luzhna, L.; Kovalchuk, O. Sex Difference of Radiation Response in Occupational and Accidental Exposure. Front. Genet. 2019, 10, 260. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, K.; Dendinger, M.D.; Davis, A.T.; Register, T.C.; DeBo, R.; Dugan, G.; Cline, J.M. Type 2 Diabetes is a Delayed Late Effect of Whole-Body Irradiation in Nonhuman Primates. Radiat. Res. 2015, 183, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Martinou, M.; Gaya, A. Cardiac complications after radical radiotherapy. Semin. Oncol. 2013, 40, 178–185. [Google Scholar] [CrossRef]
- Tapio, S. Pathology and biology of radiation-induced cardiac disease. J. Radiat. Res. 2016, 57, 439–448. [Google Scholar] [CrossRef]
- Darby, S.C.; Ewertz, M.; McGale, P.; Bennet, A.M.; Blom-Goldman, U.; Brønnum, D.; Correa, C.; Cutter, D.; Gagliardi, G.; Gigante, B.; et al. Risk of ischemic heart disease in women after radiotherapy for breast cancer. N. Engl. J. Med. 2013, 368, 987–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, Y.; Kodama, K.; Nishi, N.; Kasagi, F.; Suyama, A.; Soda, M.; Grant, E.J.; Sugiyama, H.; Sakata, R.; Moriwaki, H.; et al. Radiation exposure and circulatory disease risk: Hiroshima and Nagasaki atomic bomb survivor data, 1950–2003. BMJ 2010, 340, b5349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, M.P.; Azizova, T.V.; Bazyka, D.; Bouffler, S.D.; Cardis, E.; Chekin, S.; Chumak, V.V.; Cucinotta, F.A.; de Vathaire, F.; Hall, P.; et al. Systematic review and meta-analysis of circulatory disease from exposure to low-level ionizing radiation and estimates of potential population mortality risks. Environ. Health Perspect. 2012, 120, 1503–1511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, J.R.; Fajardo, L.F.; Gillette, S.M.; Constine, L.S. Radiation injury to the heart. Int. J. Radiat. Oncol. Biol. Phys. 1995, 31, 1205–1211. [Google Scholar] [CrossRef]
- Slezak, J.; Kura, B.; Ravingerová, T.; Tribulova, N.; Okruhlicova, L.; Barancik, M. Mechanisms of cardiac radiation injury and potential preventive approaches. Can. J. Physiol. Pharm. 2015, 93, 737–753. [Google Scholar] [CrossRef]
- Ren, J.; Anversa, P. The insulin-like growth factor I system: Physiological and pathophysiological implication in cardiovascular diseases associated with metabolic syndrome. Biochem. Pharm. 2015, 93, 409–417. [Google Scholar] [CrossRef]
- Aguirre, G.A.; De Ita, J.R.; de la Garza, R.G.; Castilla-Cortazar, I. Insulin-like growth factor-1 deficiency and metabolic syndrome. J. Transl. Med. 2016, 14, 3. [Google Scholar] [CrossRef] [Green Version]
- Sádaba, M.C.; Martín-Estal, I.; Puche, J.E.; Castilla-Cortázar, I. Insulin-like growth factor 1 (IGF-1) therapy: Mitochondrial dysfunction and diseases. Biochim. Biophys. Acta 2016, 1862, 1267–1278. [Google Scholar] [CrossRef]
- Aghdam, S.Y.; Kenchegowda, D.; Sharma, N.K.; Holmes-Hampton, G.P.; Legesse, B.; Moroni, M.; Ghosh, S.P. Dysregulated Cardiac IGF-1 Signaling and Antioxidant Response Are Associated with Radiation Sensitivity. Int. J. Mol. Sci. 2020, 21, 5049. [Google Scholar] [CrossRef] [PubMed]
- Puche, J.E.; García-Fernández, M.; Muntané, J.; Rioja, J.; González-Barón, S.; Castilla Cortazar, I. Low doses of insulin-like growth factor-I induce mitochondrial protection in aging rats. Endocrinology 2008, 149, 2620–2627. [Google Scholar] [CrossRef] [Green Version]
- Arcopinto, M.; Bobbio, E.; Bossone, E.; Perrone-Filardi, P.; Napoli, R.; Sacca, L.; Cittadini, A. The GH/IGF-1 axis in chronic heart failure. Endocr. Metab. Immune Disord. Drug Targets 2013, 13, 76–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isgaard, J.; Arcopinto, M.; Karason, K.; Cittadini, A. GH and the cardiovascular system: An update on a topic at heart. Endocrine 2015, 48, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, E.; Andreotti, F.; Sciahbasi, A.; Riccardi, P.; Marra, G.; Menini, E.; Ghirlanda, G.; Maseri, A. Markedly reduced insulin-like growth factor-1 in the acute phase of myocardial infarction. J. Am. Coll. Cardiol. 2001, 38, 26–32. [Google Scholar] [CrossRef]
- Juul, A.; Scheike, T.; Davidsen, M.; Gyllenborg, J.; Jørgensen, T. Low serum insulin-like growth factor I is associated with increased risk of ischemic heart disease: A population-based case-control study. Circulation 2002, 106, 939–944. [Google Scholar] [CrossRef] [Green Version]
- Ungvari, Z.; Csiszar, A. The emerging role of IGF-1 deficiency in cardiovascular aging: Recent advances. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Brown-Borg, H.M. Impaired cardiac excitation–contraction coupling in ventricular myocytes from Ames dwarf mice with IGF-I deficiency. Growth Horm. IGF Res. 2002, 12, 99–105. [Google Scholar] [CrossRef]
- Aghdam, S.Y.; Kenchegowda, D.; Holmes-Hampton, G.P.; Moroni, M.; Ghosh, S.P. Impairment of IGF-1 Signaling and Antioxidant Response Are Associated with Radiation Sensitivity and Mortality. Int. J. Mol. Sci. 2021, 22, 451. [Google Scholar] [CrossRef]
- Troncoso, R.; Ibarra, C.; Vicencio, J.M.; Jaimovich, E.; Lavandero, S. New insights into IGF-1 signaling in the heart. Trends Endocrinol. Metab. 2014, 25, 128–137. [Google Scholar] [CrossRef]
- Badorff, C.; Dimmeler, S. NO Balance. Circulation 2003, 107, 1348–1349. [Google Scholar] [CrossRef] [Green Version]
- Castellano, G.; Affuso, F.; Conza, P.D.; Fazio, S. The GH/IGF-1 Axis and Heart Failure. Curr. Cardiol. Rev. 2009, 5, 203–215. [Google Scholar] [CrossRef]
- Donahue, L.R.; Beamer, W.G. Growth hormone deficiency in ‘little’ mice results in aberrant body composition, reduced insulin-like growth factor-I and insulin-like growth factor-binding protein-3 (IGFBP-3), but does not affect IGFBP-2, -1 or -4. J. Endocrinol. 1993, 136, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Downs, L.C.; Mitschelen, M.; Sosnowska, D.; Toth, P.; Pinto, J.T.; Ballabh, P.; Valcarcel-Ares, M.N.; Farley, J.; Koller, A.; Henthorn, J.C.; et al. Liver-specific knockdown of IGF-1 decreases vascular oxidative stress resistance by impairing the Nrf2-dependent antioxidant response: A novel model of vascular aging. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 313–329. [Google Scholar] [CrossRef]
- Csiszar, A.; Labinskyy, N.; Perez, V.; Recchia, F.A.; Podlutsky, A.; Mukhopadhyay, P.; Losonczy, G.; Pacher, P.; Austad, S.N.; Bartke, A.; et al. Endothelial function and vascular oxidative stress in long-lived GH/IGF-deficient Ames dwarf mice. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1882–H1894. [Google Scholar] [CrossRef] [Green Version]
- Puche, J.E.; Castilla-Cortázar, I. Human conditions of insulin-like growth factor-I (IGF-I) deficiency. J. Transl. Med. 2012, 10, 224. [Google Scholar] [CrossRef] [Green Version]
- Kamenický, P.; Mazziotti, G.; Lombès, M.; Giustina, A.; Chanson, P. Growth hormone, insulin-like growth factor-1, and the kidney: Pathophysiological and clinical implications. Endocr. Rev. 2014, 35, 234–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Garza, R.G.; Morales-Garza, L.A.; Martin-Estal, I.; Castilla-Cortazar, I. Insulin-Like Growth Factor-1 Deficiency and Cirrhosis Establishment. J. Clin. Med. Res. 2017, 9, 233–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ming, X.; Feng, Y.; Yang, C.; Wang, W.; Wang, P.; Deng, J. Radiation-induced heart disease in lung cancer radiotherapy: A dosimetric update. Medicine 2016, 95, e5051. [Google Scholar] [CrossRef]
- Valenciano, A.; Henríquez-Hernández, L.A.; Moreno, M.; Lloret, M.; Lara, P.C. Role of IGF-1 receptor in radiation response. Transl. Oncol. 2012, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatachalam, S.; Mettler, E.; Fottner, C.; Miederer, M.; Kaina, B.; Weber, M.M. The impact of the IGF-1 system of cancer cells on radiation response—An in vitro study. Clin. Transl. Radiat. Oncol. 2017, 7, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azimzadeh, O.; Sievert, W.; Sarioglu, H.; Merl-Pham, J.; Yentrapalli, R.; Bakshi, M.V.; Janik, D.; Ueffing, M.; Atkinson, M.J.; Multhoff, G.; et al. Integrative proteomics and targeted transcriptomics analyses in cardiac endothelial cells unravel mechanisms of long-term radiation-induced vascular dysfunction. J. Proteome Res. 2015, 14, 1203–1219. [Google Scholar] [CrossRef]
- Kenchegowda, D.; Legesse, B.; Hritzo, B.; Olsen, C.; Aghdam, S.; Kaur, A.; Culp, W.; Derrien-Colemyn, A.; Severson, G.; Moroni, M. Selective Insulin-like Growth Factor Resistance Associated with Heart Hemorrhages and Poor Prognosis in a Novel Preclinical Model of the Hematopoietic Acute Radiation Syndrome. Radiat. Res. 2018, 190, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Venkatesulu, B.P.; Mahadevan, L.S.; Aliru, M.L.; Yang, X.; Bodd, M.H.; Singh, P.K.; Yusuf, S.W.; Abe, J.I.; Krishnan, S. Radiation-Induced Endothelial Vascular Injury: A Review of Possible Mechanisms. JACC Basic Transl. Sci. 2018, 3, 563–572. [Google Scholar] [CrossRef]
- Legesse, B.; Kaur, A.; Kenchegowda, D.; Hritzo, B.; Culp, W.E.; Moroni, M. Neulasta Regimen for the Hematopoietic Acute Radiation Syndrome: Effects Beyond Neutrophil Recovery. Int. J. Radiat. Oncol. Biol. Phys. 2019, 103, 935–944. [Google Scholar] [CrossRef]
- Kobayashi, E.; Hishikawa, S.; Teratani, T.; Lefor, A.T. The pig as a model for translational research: Overview of porcine animal models at Jichi Medical University. Transpl. Res. 2012, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.L.; Hartvigsen, M.L.; Hedemann, M.S.; Lærke, H.N.; Hermansen, K.; Bach Knudsen, K.E. Similar metabolic responses in pigs and humans to breads with different contents and compositions of dietary fibers: A metabolomics study. Am. J. Clin. Nutr. 2014, 99, 941–949. [Google Scholar] [CrossRef] [PubMed]
- Matthan, N.R.; Solano-Aguilar, G.; Meng, H.; Lamon-Fava, S.; Goldbaum, A.; Walker, M.E.; Jang, S.; Lakshman, S.; Molokin, A.; Xie, Y.; et al. The Ossabaw Pig Is a Suitable Translational Model to Evaluate Dietary Patterns and Coronary Artery Disease Risk. J. Nutr. 2018, 148, 542–551. [Google Scholar] [CrossRef]
- Moroni, M.; Coolbaugh, T.V.; Lombardini, E.; Mitchell, J.M.; Moccia, K.D.; Shelton, L.J.; Nagy, V.; Whitnall, M.H. Hematopoietic Radiation Syndrome in the Gottingen Minipig. Radiat. Res. 2011, 176, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Moroni, M.; Lombardini, E.; Salber, R.; Kazemzedeh, M.; Nagy, V.; Olsen, C.; Whitnall, M.H. Hematological Changes as Prognostic Indicators of Survival: Similarities Between Gottingen Minipigs, Humans, and Other Large Animal Models. PLoS ONE 2011, 6, e25210. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, X.M.; Meinkoth, J.; Pittman, R.N. Akt regulates cell survival and apoptosis at a postmitochondrial level. J. Cell Biol. 2000, 151, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Gallo, S.; Vitacolonna, A.; Bonzano, A.; Comoglio, P.; Crepaldi, T. ERK: A Key Player in the Pathophysiology of Cardiac Hypertrophy. Int. J. Mol. Sci. 2019, 20, 2164. [Google Scholar] [CrossRef] [Green Version]
- González-Guerra, J.L.; Castilla-Cortazar, I.; Aguirre, G.A.; Muñoz, Ú.; Martín-Estal, I.; Ávila-Gallego, E.; Granado, M.; Puche, J.E.; García-Villalón, Á.L. Partial IGF-1 deficiency is sufficient to reduce heart contractibility, angiotensin II sensibility, and alter gene expression of structural and functional cardiac proteins. PLoS ONE 2017, 12, e0181760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- el Azzouzi, H.; Leptidis, S.; Bourajjaj, M.; Armand, A.-S.; van der Nagel, R.; van Bilsen, M.; Da Costa Martins, P.A.; De Windt, L.J. Peroxisome proliferator-activated receptor (PPAR) gene profiling uncovers insulin-like growth factor-1 as a PPARalpha target gene in cardioprotection. J. Biol. Chem. 2011, 286, 14598–14607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kersten, S. Peroxisome proliferator activated receptors and lipoprotein metabolism. PPAR Res. 2008, 2008, 132960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pautsch, A.; Zoephel, A.; Ahorn, H.; Spevak, W.; Hauptmann, R.; Nar, H. Crystal structure of bisphosphorylated IGF-1 receptor kinase: Insight into domain movements upon kinase activation. Structure 2001, 9, 955–965. [Google Scholar] [CrossRef] [Green Version]
- Preston, D.L.; Shimizu, Y.; Pierce, D.A.; Suyama, A.; Mabuchi, K. Studies of mortality of atomic bomb survivors. Report 13: Solid cancer and noncancer disease mortality: 1950–1997. Radiat. Res. 2003, 160, 381–407. [Google Scholar] [CrossRef]
- Stoppel, G.; Eich, H.-T.; Matuschek, C.; Kortmann, R.-D.; Meyer, F.; Martinsson, U.; Nilsson, K.; Kristensen, I.; Vordermark, D.; Willich, N.; et al. Lung toxicity after radiation in childhood: Results of the International Project on Prospective Analysis of Radiotoxicity in Childhood and Adolescence. Radiother. Oncol. J. Eur. Soc. Ther. Radiol. Oncol. 2017, 125, 286–292. [Google Scholar] [CrossRef]
- Simone, C.B., II. Thoracic Radiation Normal Tissue Injury. Semin. Radiat. Oncol. 2017, 27, 370–377. [Google Scholar] [CrossRef]
- Lawrence, J.H.; Tennant, R. The comparative effects of neutrons and x-rays on the whole body. J. Exp. Med. 1937, 66, 667–688. [Google Scholar] [CrossRef]
- Vasilenko, I.; Osipov, V.A.; Turova, V.I.; Istomina, A.G. Delayed effects of acute radiation injuries by radioactive carbon (14C-glycine). Radiobiologiia 1985, 25, 815–819. [Google Scholar] [PubMed]
- Chiang, C.S.; Hong, J.H.; Stalder, A.; Sun, J.R.; Withers, H.R.; McBride, W.H. Delayed molecular responses to brain irradiation. Int. J. Radiat. Biol. 1997, 72, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Shankar, B.; Premachandran, S.; Bharambe, S.D.; Sundaresan, P.; Sainis, K.B. Modification of immune response by low dose ionizing radiation: Role of apoptosis. Immunol. Lett. 1999, 68, 237–245. [Google Scholar] [CrossRef]
- Martin, C.; Martin, S.; Viret, R.; Denis, J.; Mirguet, F.; Diserbo, M.; Multon, E.; Lamproglou, I. Low dose of the gamma acute radiation syndrome (1.5 Gy) does not significantly alter either cognitive behavior or dopaminergic and serotoninergic metabolism. Cell Mol. Biol. 2001, 47, 459–465. [Google Scholar]
- Epperly, M.W.; Dixon, T.; Wang, H.; Schlesselman, J.; Franicola, D.; Greenberger, J.S. Modulation of Radiation-Induced Life Shortening by Systemic Intravenous MnSOD-Plasmid Liposome Gene Therapy. Radiat. Res. 2008, 170, 437–443. [Google Scholar] [CrossRef]
- Chua, H.L.; Plett, P.A.; Fisher, A.; Sampson, C.H.; Vemula, S.; Feng, H.; Sellamuthu, R.; Wu, T.; MacVittie, T.J.; Orschell, C.M. Lifelong Residual bone Marrow Damage in Murine Survivors of the Hematopoietic Acute Radiation Syndrome (H-ARS): A Compilation of Studies Comprising the Indiana University Experience. Health Phys. 2019, 116, 546–557. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yu, J.; Jones, J.W.; Carter, C.L.; Jackson, I.L.; Vujaskovic, Z.; MacVittie, T.J.; Kane, M.A. Acute Proteomic Changes in the Lung After WTLI in a Mouse Model: Identification of Potential Initiating Events for Delayed Effects of Acute Radiation Exposure. Health Phys. 2019, 116, 503–515. [Google Scholar] [CrossRef]
- Micewicz, E.D.; Iwamoto, K.S.; Ratikan, J.A.; Nguyen, C.; Xie, M.W.; Cheng, G.; Boxx, G.M.; Deriu, E.; Damoiseaux, R.D.; Whitelegge, J.P.; et al. The Aftermath of Surviving Acute Radiation Hematopoietic Syndrome and its Mitigation. Radiat. Res. 2019, 191, 323–334. [Google Scholar] [CrossRef]
- Unthank, J.L.; Miller, S.J.; Quickery, A.K.; Ferguson, E.L.; Wang, M.; Sampson, C.H.; Chua, H.L.; DiStasi, M.R.; Feng, H.; Fisher, A.; et al. Delayed Effects of Acute Radiation Exposure in a Murine Model of the H-ARS: Multiple-Organ Injury Consequent to <10 Gy Total Body Irradiation. Health Phys. 2015, 109, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Seol, M.A.; Jung, U.; Eom, H.S.; Kim, S.H.; Park, H.R.; Jo, S.K. Prolonged expression of senescence markers in mice exposed to gamma-irradiation. J. Vet. Sci. 2012, 13, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.K.; Holmes-Hampton, G.P.; Kumar, V.P.; Biswas, S.; Wuddie, K.; Stone, S.; Aschenake, Z.; Wilkins, W.L.; Fam, C.M.; Cox, G.N.; et al. Delayed effects of acute whole body lethal radiation exposure in mice pre-treated with BBT-059. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacVittie, T.J.; Bennett, A.; Booth, C.; Garofalo, M.; Tudor, G.; Ward, A.; Shea-Donohue, T.; Gelfond, D.; McFarland, E.; Jackson, W., III; et al. The prolonged gastrointestinal syndrome in rhesus macaques: The relationship between gastrointestinal, hematopoietic, and delayed multi-organ sequelae following acute, potentially lethal, partial-body irradiation. Health Phys. 2012, 103, 427–453. [Google Scholar] [CrossRef] [Green Version]
- Booth, C.; Tudor, G.; Tonge, N.; Shea-Donohue, T.; MacVittie, T.J. Evidence of delayed gastrointestinal syndrome in high-dose irradiated mice. Health Phys. 2012, 103, 400–410. [Google Scholar] [CrossRef] [Green Version]
- Booth, C.; Tudor, G.L.; Katz, B.P.; MacVittie, T.J. The Delayed Effects of Acute Radiation Syndrome: Evidence of Long-Term Functional Changes in the Clonogenic Cells of the Small Intestine. Health Phys. 2015, 109, 399–413. [Google Scholar] [CrossRef] [Green Version]
- Cohen, E.P.; Hankey, K.G.; Farese, A.M.; Parker, G.A.; Jones, J.W.; Kane, M.A.; Bennett, A.; MacVittie, T.J. Radiation Nephropathy in a Nonhuman Primate Model of Partial-body Irradiation with Minimal Bone Marrow Sparing-Part 1: Acute and Chronic Kidney Injury and the Influence of Neupogen. Health Phys. 2019, 116, 401–408. [Google Scholar] [CrossRef]
- Garofalo, M.; Bennett, A.; Farese, A.M.; Harper, J.; Ward, A.; Taylor-Howell, C.; Cui, W.; Gibbs, A.; Lasio, G.; Jackson, W.; et al. The delayed pulmonary syndrome following acute high-dose irradiation: A rhesus macaque model. Health Phys. 2014, 106, 56–72. [Google Scholar] [CrossRef]
- Medhora, M.; Gao, F.; Gasperetti, T.; Narayanan, J.; Khan, A.H.; Jacobs, E.R.; Fish, B.L. Delayed Effects of Acute Radiation Exposure (Deare) in Juvenile and Old Rats: Mitigation by Lisinopril. Health Phys. 2019, 116, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Mathias, D.; Mitchel, R.E.J.; Barclay, M.; Wyatt, H.; Bugden, M.; Priest, N.D.; Whitman, S.C.; Scholz, M.; Hildebrandt, G.; Kamprad, M.; et al. Correction: Low-Dose Irradiation Affects Expression of Inflammatory Markers in the Heart of ApoE -/- Mice. PLoS ONE 2016, 11, e0157616. [Google Scholar] [CrossRef] [PubMed]
- Adamek, A.; Kasprzak, A. Insulin-Like Growth Factor (IGF) System in Liver Diseases. Int. J. Mol. Sci. 2018, 19, 1308. [Google Scholar] [CrossRef] [Green Version]
- Yakar, S.; Liu, J.-L.; Stannard, B.; Butler, A.; Accili, D.; Sauer, B.; LeRoith, D. Normal growth and development in the absence of hepatic insulin-like growth factor I. Proc. Natl. Acad. Sci. USA 1999, 96, 7324–7329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, M.; Rosen, C.J. The IGF-I regulatory system and its impact on skeletal and energy homeostasis. J. Cell. Biochem. 2010, 111, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locatelli, V.; Bianchi, V.E. Effect of GH/IGF-1 on Bone Metabolism and Osteoporsosis. Int. J. Endocrinol. 2014, 2014, 235060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, R.; García-Fernández, M.; Díaz-Sánchez, M.; Puche, J.E.; Delgado, G.; Conchillo, M.; Muntané, J.; Castilla-Cortázar, I. Mitochondrial protection by low doses of insulin-like growth factor-Iin experimental cirrhosis. World J. Gastroenterol. 2008, 14, 2731–2739. [Google Scholar] [CrossRef]
- Touvron, M.; Escoubet, B.; Mericskay, M.; Angelini, A.; Lamotte, L.; Santini, M.P.; Rosenthal, N.; Daegelen, D.; Tuil, D.; Decaux, J.-F. Locally expressed IGF1 propeptide improves mouse heart function in induced dilated cardiomyopathy by blocking myocardial fibrosis and SRF-dependent CTGF induction. Dis. Models Mech. 2012, 5, 481–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seawright, J.W.; Sridharan, V.; Landes, R.D.; Cao, M.; Singh, P.; Koturbash, I.; Mao, X.-W.; Miousse, I.R.; Singh, S.P.; Nelson, G.A.; et al. Effects of low-dose oxygen ions and protons on cardiac function and structure in male C57BL/6J mice. Life Sci. Space Res. 2019, 20, 72–84. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose (Gy) | Heart, Left Ventricle and Atrium | Lung, Right Diaphragmatic Lobe | Left Kidney |
|---|---|---|---|
| 2 | arterial pigmentation | hemorrhage, mild fibrosis | nephropathy, mild fibrosis |
| 2 | arterial pigmentation | minimal fibrosis | nephropathy, mild fibrosis |
| 2 | no visible lesions | no visible lesions | mild fibrosis |
| 1.9 | arterial pigmentation | edema, congestion | mild fibrosis |
| 1.9 | no visible lesions | minimal fibrosis | minimal fibrosis |
| 1.9 | no visible lesions | no visible lesions | mild fibrosis |
| 1.9 | no visible lesions | no visible lesions | minimal fibrosis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hritzo, B.; Aghdam, S.Y.; Legesse, B.; Kaur, A.; Cao, M.; Boerma, M.; Chakraborty, N.; Dimitrov, G.; Gautam, A.; Hammamieh, R.; et al. Late Health Effects of Partial Body Irradiation Injury in a Minipig Model Are Associated with Changes in Systemic and Cardiac IGF-1 Signaling. Int. J. Mol. Sci. 2021, 22, 3286. https://doi.org/10.3390/ijms22063286
Hritzo B, Aghdam SY, Legesse B, Kaur A, Cao M, Boerma M, Chakraborty N, Dimitrov G, Gautam A, Hammamieh R, et al. Late Health Effects of Partial Body Irradiation Injury in a Minipig Model Are Associated with Changes in Systemic and Cardiac IGF-1 Signaling. International Journal of Molecular Sciences. 2021; 22(6):3286. https://doi.org/10.3390/ijms22063286
Chicago/Turabian StyleHritzo, Bernadette, Saeed Y. Aghdam, Betre Legesse, Amandeep Kaur, Maohua Cao, Marjan Boerma, Nabarun Chakraborty, George Dimitrov, Aarti Gautam, Rasha Hammamieh, and et al. 2021. "Late Health Effects of Partial Body Irradiation Injury in a Minipig Model Are Associated with Changes in Systemic and Cardiac IGF-1 Signaling" International Journal of Molecular Sciences 22, no. 6: 3286. https://doi.org/10.3390/ijms22063286
APA StyleHritzo, B., Aghdam, S. Y., Legesse, B., Kaur, A., Cao, M., Boerma, M., Chakraborty, N., Dimitrov, G., Gautam, A., Hammamieh, R., Wilkins, W., Tsioplaya, A., Holmes-Hampton, G. P., & Moroni, M. (2021). Late Health Effects of Partial Body Irradiation Injury in a Minipig Model Are Associated with Changes in Systemic and Cardiac IGF-1 Signaling. International Journal of Molecular Sciences, 22(6), 3286. https://doi.org/10.3390/ijms22063286