
Fibrinolysis: A Primordial System Linked to the Immune Response


Abstract
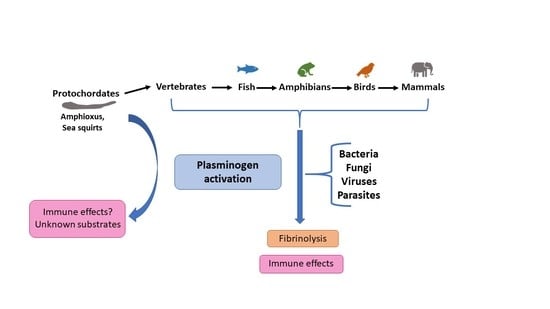
1. Introduction
2. Plasminogen Activation: A Universal System with a Broad Repertoire
3. Plasminogen, Fibrinolysis and Immune Function
4. Microorganisms, Plasmin Formation and Fibrinolysis
5. Phylogenetic Links with Plasminogen Activation
6. Conclusions, Clinical Implications and Future Potential
Author Contributions
Funding
Conflicts of Interest
References
- Draxler, D.F.; Sashindranath, M.; Medcalf, R.L. Plasmin: A Modulator of Immune Function. Semin. Thromb. Hemost. 2017, 43, 143–153. [Google Scholar] [CrossRef]
- Green, J.R. Note on the Action of Sodium Chloride in dissolving Fibrin. J. Physiol. 1887, 8, 372–377. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dastre, A. Fibrinolyse dans le sang. Arch. Physiol. 1893, 5, 661. [Google Scholar]
- Bier, O.E. Action anticoagulante et fibrinolytique de l’extract des glands salivaires d’une chauve-souris hematophage (desmodus rufus). C.R. Soc Biol. (Paris) 1932, 110, 129–131. [Google Scholar]
- Aoi, F. On the fibrolysis of the staphylococcus. Kitasato Arch. Exptl. Med. 1932, 9, 171–201. [Google Scholar]
- Lack, C.H. Staphylokinase: An activator of plasma Protease. Nature 1948, 559–560. [Google Scholar] [CrossRef]
- Tillett, W.S.; Garner, R.L. The Fibrinolytic Activity of Hemolytic Streptococci. J. Exp. Med. 1933, 58, 485–502. [Google Scholar] [CrossRef]
- Milstone, H. A factor in normal human blood which particiaptes in streptococcal fibrinolysis. J. Immunol. 1941, 42, 109–116. [Google Scholar]
- Macfarlane, R.G.; Pilling, J. Observations on fibrinolysis; plasminogen, plasmin, and antiplasmin content of human blood. Lancet 1946, 2, 562–565. [Google Scholar] [CrossRef]
- Astrup, T.; Permin, P.M. Fibrinolysis in the animal organism. Nature 1947, 159, 681. [Google Scholar] [CrossRef] [PubMed]
- Sobel, G.W.; Mohler, S.R.; Jones, N.W.; Dowdy, A.B.C.; Guest, M.M. Urokinase: An activator of plasma profibrinolysin extracted from urine. Am. J. Physiol. 1952, 171, 768–769. [Google Scholar]
- Parry, M.A.; Zhang, X.C.; Bode, I. Molecular mechanisms of plasminogen activation: Bacterial cofactors provide clues. Trends Biochem. Sci. 2000, 25, 53–59. [Google Scholar] [CrossRef]
- Tillett, W.S.; Sherry, S. The effect in patients of streptococcal fibrinolysin and streptococcal desoxyribonuclease on fibrinous, purulent, and sanguinous pleural exudations. J. Clin. Investig. 1949, 28, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Lijnen, H.R.; Van Hoef, B.; De Cock, F.; Okada, K.; Ueshima, S.; Matsuo, O.; Collen, D. On the mechanism of fibrin-specific plasminogen activation by staphylokinase. J. Biol. Chem. 1991, 266, 11826–11832. [Google Scholar] [CrossRef]
- Okamoto, S.O.U. A new potent antifibrinolytic substance and its effects on blood of animals. Keio J. Med. 1962, 11, 105–115. [Google Scholar] [CrossRef]
- Jorg, M.; Binder, B.R. Kinetic analysis of plasminogen activation by purified plasma kallikrein. Thromb. Res. 1985, 39, 323–331. [Google Scholar] [CrossRef]
- Foley, J.H. Plasmin(ogen) at the Nexus of Fibrinolysis, Inflammation, and Complement. Semin. Thromb. Hemost. 2017, 43, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Tomczyk, M.; Suzuki, Y.; Sano, H.; Brzoska, T.; Tanaka, H.; Urano, T. Bidirectional functions of thrombin on fibrinolysis: Evidence of thrombin-dependent enhancement of fibrinolysis provided by spontaneous plasma clot lysis. Thromb. Res. 2016, 143, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Lwaleed, B.A.; Greenfield, R.; Stewart, A.; Birch, B.; Cooper, A.J. Seminal clotting and fibrinolytic balance: A possible physiological role in the male reproductive system. Thromb. Haemost. 2004, 92, 752–766. [Google Scholar] [CrossRef] [PubMed]
- Basham, M.E.; Seeds, N.W. Plasminogen expression in the neonatal and adult mouse brain. J. Neurochem. 2001, 77, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Seiffert, D.; Fowler, B.J.; Jenkins, G.R.; Thinnes, T.C.; Loskutoff, D.J.; Parmer, R.J.; Miles, L.A. Plasminogen Has a Broad Extrahepatic Distribution. Thromb. Haemost. 2002, 87, 493–501. [Google Scholar] [PubMed]
- Del Giudice, P.T.; da Silva, B.F.; Lo Turco, E.G.; Fraietta, R.; Spaine, D.M.; Santos, L.F.; Pilau, E.J.; Gozzo, F.C.; Cedenho, A.P.; Bertolla, R.P. Changes in the seminal plasma proteome of adolescents before and after varicocelectomy. Fertil. Steril. 2013, 100, 667–672. [Google Scholar] [CrossRef]
- Kawashita, E.; Kanno, Y.; Asayama, H.; Okada, K.; Ueshima, S.; Matsuo, O.; Matsuno, H. Involvement of alpha2-antiplasmin in dendritic growth of hippocampal neurons. J. Neurochem. 2013, 126, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Medcalf, R.L. Fibrinolysis: From blood to the brain. J. Thromb. Haemost. 2017, 15, 2089–2098. [Google Scholar] [CrossRef]
- Yepes, M.; Lawrence, D.A. New functions for an old enzyme: Nonhemostatic roles for tissue-type plasminogen activator in the central nervous system. Exp. Biol. Med. (Maywood) 2004, 229, 1097–1104. [Google Scholar] [CrossRef]
- Samson, A.L.; Medcalf, R.L. Tissue-type plasminogen activator: A multifaceted modulator of neurotransmission and synaptic plasticity. Neuron 2006, 50, 673–678. [Google Scholar] [CrossRef]
- Draxler, D.F.; Medcalf, R.L. The fibrinolytic system-more than fibrinolysis? Transfus. Med. Rev. 2015, 29, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, L.; Lawrence, D.A.; Medcalf, R.L. tPA Modulation of the Blood-Brain Barrier: A Unifying Explanation for the Pleiotropic Effects of tPA in the CNS. Semin. Thromb. Hemost. 2017, 43, 154–168. [Google Scholar] [CrossRef]
- Medcalf, R.L.; Keragala, C.; Draxler, D.F. Fibrinolysis and the immune response in trauma. Semin. Thromb. Haemost. 2019, in press. [Google Scholar]
- Warnaar, N.; Mallett, S.V.; Klinck, J.R.; de Boer, M.T.; Rolando, N.; Burroughs, A.K.; Jamieson, N.V.; Rolles, K.; Porte, R.J. Aprotinin and the risk of thrombotic complications after liver transplantation: A retrospective analysis of 1492 patients. Liver Transpl. 2009, 15, 747–753. [Google Scholar] [CrossRef]
- Henry, D.A.; Carless, P.A.; Moxey, A.J.; O’Connell, D.; Stokes, B.J.; Fergusson, D.A.; Ker, K. Anti-fibrinolytic use for minimising perioperative allogeneic blood transfusion. Cochrane Database Syst. Rev. 2011, CD001886. [Google Scholar] [CrossRef]
- Li, Q.; Laumonnier, Y.; Syrovets, T.; Simmet, T. Plasmin triggers cytokine induction in human monocyte-derived macrophages. Arter. Thromb Vasc Biol 2007, 27, 1383–1389. [Google Scholar] [CrossRef]
- Draxler, D.F.; Yep, K.; Hanafi, G.; Winton, A.; Daglas, M.; Ho, H.; Sashindranath, M.; Wutzlhofer, L.M.; Forbes, A.; Goncalves, I.; et al. Tranexamic acid modulates the immune response and reduces postsurgical infection rates. Blood Adv. 2019, 3, 1598–1609. [Google Scholar] [CrossRef]
- Miles, L.A.; Parmer, R.J. Plasminogen receptors: The first quarter century. Semin Thromb Hemost 2013, 39, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.M.; Lum, A.G.; Tran, C.; Shaw, M.W.; Gao, Z.; Flick, M.J.; Moutsopoulos, N.M.; Bugge, T.H.; Mullins, E.S. Plasmin-mediated fibrinolysis enables macrophage migration in a murine model of inflammation. Blood 2019, 134, 291–303. [Google Scholar] [CrossRef]
- Das, R.; Ganapathy, S.; Settle, M.; Plow, E.F. Plasminogen promotes macrophage phagocytosis in mice. Blood 2014, 124, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.A.; Ribeiro, A.L.C.; Costa, B.R.C.; Vago, J.P.; Lima, K.M.; Carneiro, F.S.; Ortiz, M.M.O.; Lima, G.L.N.; Carmo, A.A.F.; Rocha, R.M.; et al. Plasmin and plasminogen induce macrophage reprogramming and regulate key steps of inflammation resolution via annexin A1. Blood 2017, 129, 2896–2907. [Google Scholar] [CrossRef]
- Borg, R.J.; Samson, A.L.; Au, A.E.; Scholzen, A.; Fuchsberger, M.; Kong, Y.Y.; Freeman, R.; Mifsud, N.A.; Plebanski, M.; Medcalf, R.L. Dendritic Cell-Mediated Phagocytosis but Not Immune Activation Is Enhanced by Plasmin. PLoS ONE 2015, 10, e0131216. [Google Scholar] [CrossRef] [PubMed]
- Syrovets, T.; Lunov, O.; Simmet, T. Plasmin as a proinflammatory cell activator. J. Leukoc. Biol. 2012, 92, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Ayon-Nunez, D.A.; Fragoso, G.; Bobes, R.J.; Laclette, J.P. Plasminogen-binding proteins as an evasion mechanism of the host’s innate immunity in infectious diseases. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Grundel, A.; Friedrich, K.; Pfeiffer, M.; Jacobs, E.; Dumke, R. Subunits of the Pyruvate Dehydrogenase Cluster of Mycoplasma pneumoniae Are Surface-Displayed Proteins that Bind and Activate Human Plasminogen. PLoS ONE 2015, 10, e0126600. [Google Scholar] [CrossRef]
- Xolalpa, W.; Vallecillo, A.J.; Lara, M.; Mendoza-Hernandez, G.; Comini, M.; Spallek, R.; Singh, M.; Espitia, C. Identification of novel bacterial plasminogen-binding proteins in the human pathogen Mycobacterium tuberculosis. Proteomics 2007, 7, 3332–3341. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.L.; Atzingen, M.V.; Oliveira, R.; Mendes, R.S.; Domingos, R.F.; Vasconcellos, S.A.; Nascimento, A.L. Plasminogen binding proteins and plasmin generation on the surface of Leptospira spp.: The contribution to the bacteria-host interactions. J. Biomed. Biotechnol. 2012, 2012, 758513. [Google Scholar] [CrossRef] [PubMed]
- Peetermans, M.; Vanassche, T.; Liesenborghs, L.; Lijnen, R.H.; Verhamme, P. Bacterial pathogens activate plasminogen to breach tissue barriers and escape from innate immunity. Crit. Rev. Microbiol. 2016, 42, 866–882. [Google Scholar] [CrossRef]
- Sun, H.; Ringdahl, U.; Homeister, J.W.; Fay, W.P.; Engleberg, N.C.; Yang, A.Y.; Rozek, L.S.; Wang, X.; Sjobring, U.; Ginsburg, D. Plasminogen is a critical host pathogenicity factor for group A streptococcal infection. Science 2004, 305, 1283–1286. [Google Scholar] [CrossRef] [PubMed]
- Gladysheva, I.P.; Turner, R.B.; Sazonova, I.Y.; Liu, L.; Reed, G.L. Coevolutionary patterns in plasminogen activation. Proc. Natl. Acad. Sci. USA 2003, 100, 9168–9172. [Google Scholar] [CrossRef] [PubMed]
- Eddy, J.L.; Schroeder, J.A.; Zimbler, D.L.; Caulfield, A.J.; Lathem, W.W. Proteolysis of plasminogen activator inhibitor-1 by Yersinia pestis remodulates the host environment to promote virulence. J. Thromb. Haemost. 2016, 14, 1833–1843. [Google Scholar] [CrossRef]
- Sodeinde, O.A.; Subrahmanyam, Y.V.; Stark, K.; Quan, T.; Bao, Y.; Goguen, J.D. A surface protease and the invasive character of plague. Science 1992, 258, 1004–1007. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.K.; Crane, S.D.; Pechous, R.D. A Dual Role for the Plasminogen Activator Protease During the Preinflammatory Phase of Primary Pneumonic Plague. J. Infect. Dis. 2020, 222, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Knaust, A.; Weber, M.V.; Hammerschmidt, S.; Bergmann, S.; Frosch, M.; Kurzai, O. Cytosolic proteins contribute to surface plasminogen recruitment of Neisseria meningitidis. J. Bacteriol. 2007, 189, 3246–3255. [Google Scholar] [CrossRef]
- Crane, D.D.; Warner, S.L.; Bosio, C.M. A novel role for plasmin-mediated degradation of opsonizing antibody in the evasion of host immunity by virulent, but not attenuated, Francisella tularensis. J. Immunol. 2009, 183, 4593–4600. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, G.H.; Atzingen, M.V.; de Souza, G.O.; Vasconcellos, S.A.; Nascimento, A. Leptospira interrogans Lsa23 protein recruits plasminogen, factor H and C4BP from normal human serum and mediates C3b and C4b degradation. Microbiology (Reading) 2016, 162, 295–308. [Google Scholar] [CrossRef]
- Crowe, J.D.; Sievwright, I.K.; Auld, G.C.; Moore, N.R.; Gow, N.A.; Booth, N.A. Candida albicans binds human plasminogen: Identification of eight plasminogen-binding proteins. Mol. Microbiol. 2003, 47, 1637–1651. [Google Scholar] [CrossRef]
- Funk, J.; Schaarschmidt, B.; Slesiona, S.; Hallstrom, T.; Horn, U.; Brock, M. The glycolytic enzyme enolase represents a plasminogen-binding protein on the surface of a wide variety of medically important fungal species. Int. J. Med. Microbiol. 2016, 306, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Stie, J.; Fox, D. Blood-brain barrier invasion by Cryptococcus neoformans is enhanced by functional interactions with plasmin. Microbiology (Reading) 2012, 158, 240–258. [Google Scholar] [CrossRef]
- Vanegas, G.; Quinones, W.; Carrasco-Lopez, C.; Concepcion, J.L.; Albericio, F.; Avilan, L. Enolase as a plasminogen binding protein in Leishmania mexicana. Parasitol. Res. 2007, 101, 1511–1516. [Google Scholar] [CrossRef]
- Roggwiller, E.; Fricaud, A.C.; Blisnick, T.; Braun-Breton, C. Host urokinase-type plasminogen activator participates in the release of malaria merozoites from infected erythrocytes. Mol. Biochem. Parasitol. 1997, 86, 49–59. [Google Scholar] [CrossRef]
- Alves, E.S.T.L.; Radtke, A.; Balaban, A.; Pascini, T.V.; Pala, Z.R.; Roth, A.; Alvarenga, P.H.; Jeong, Y.J.; Olivas, J.; Ghosh, A.K.; et al. The fibrinolytic system enables the onset of Plasmodium infection in the mosquito vector and the mammalian host. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Mantuano, E.; Brifault, C.; Lam, M.S.; Azmoon, P.; Gilder, A.S.; Gonias, S.L. LDL receptor-related protein-1 regulates NFkappaB and microRNA-155 in macrophages to control the inflammatory response. Proc. Natl. Acad. Sci. USA 2016, 113, 1369–1374. [Google Scholar] [CrossRef]
- Mantuano, E.; Azmoon, P.; Brifault, C.; Banki, M.A.; Gilder, A.S.; Campana, W.M.; Gonias, S.L. Tissue-type plasminogen activator regulates macrophage activation and innate immunity. Blood 2017, 130, 1364–1374. [Google Scholar] [CrossRef] [PubMed]
- Das, L.; Banki, M.A.; Azmoon, P.; Pizzo, D.; Gonias, S.L. Enzymatically Inactive Tissue-Type Plasminogen Activator Reverses Disease Progression in the Dextran Sulfate Sodium Mouse Model of Inflammatory Bowel Disease. Am. J. Pathol. 2021. [Google Scholar] [CrossRef]
- Thaler, B.; Baik, N.; Hohensinner, P.J.; Baumgartner, J.; Panzenbock, A.; Stojkovic, S.; Demyanets, S.; Huk, I.; Rega-Kaun, G.; Kaun, C.; et al. Differential expression of Plg-RKT and its effects on migration of proinflammatory monocyte and macrophage subsets. Blood 2019, 134, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Ganapathy, S.; Mahabeleshwar, G.H.; Drumm, C.; Febbraio, M.; Jain, M.K.; Plow, E.F. Macrophage gene expression and foam cell formation are regulated by plasminogen. Circulation 2013, 127, 1209–1218, e1201–e1216. [Google Scholar] [CrossRef]
- Vago, J.P.; Sugimoto, M.A.; Lima, K.M.; Negreiros-Lima, G.L.; Baik, N.; Teixeira, M.M.; Perretti, M.; Parmer, R.J.; Miles, L.A.; Sousa, L.P. Plasminogen and the Plasminogen Receptor, Plg-RKT, Regulate Macrophage Phenotypic, and Functional Changes. Front. Immunol. 2019, 10, 1458. [Google Scholar] [CrossRef] [PubMed]
- Syrovets, T.; Tippler, B.; Rieks, M.; Simmet, T. Plasmin is a potent and specific chemoattractant for human peripheral monocytes acting via a cyclic guanosine monophosphate-dependent pathway. Blood 1997, 89, 4574–4583. [Google Scholar] [CrossRef] [PubMed]
- Weide, I.; Tippler, B.; Syrovets, T.; Simmet, T. Plasmin is a specific stimulus of the 5-lipoxygenase pathway of human peripheral monocytes. Thromb. Haemost. 1996, 76, 561–568. [Google Scholar] [CrossRef]
- Burysek, L.; Syrovets, T.; Simmet, T. The serine protease plasmin triggers expression of MCP-1 and CD40 in human primary monocytes via activation of p38 MAPK and janus kinase (JAK)/STAT signaling pathways. J. Biol. Chem. 2002, 277, 33509–33517. [Google Scholar] [CrossRef]
- Li, X.; Syrovets, T.; Genze, F.; Pitterle, K.; Oberhuber, A.; Orend, K.H.; Simmet, T. Plasmin triggers chemotaxis of monocyte-derived dendritic cells through an Akt2-dependent pathway and promotes a T-helper type-1 response. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Uhl, B.; Zuchtriegel, G.; Puhr-Westerheide, D.; Praetner, M.; Rehberg, M.; Fabritius, M.; Hessenauer, M.; Holzer, M.; Khandoga, A.; Furst, R.; et al. Tissue plasminogen activator promotes postischemic neutrophil recruitment via its proteolytic and nonproteolytic properties. Arter. Thromb. Vasc. Biol. 2014, 34, 1495–1504. [Google Scholar] [CrossRef]
- Li, Q.; Ke, F.; Zhang, W.; Shen, X.; Xu, Q.; Wang, H.; Yu, X.Z.; Leng, Q.; Wang, H. Plasmin plays an essential role in amplification of psoriasiform skin inflammation in mice. PLoS ONE 2011, 6, e16483. [Google Scholar] [CrossRef]
- Syrovets, T.; Jendrach, M.; Rohwedder, A.; Schule, A.; Simmet, T. Plasmin-induced expression of cytokines and tissue factor in human monocytes involves AP-1 and IKKbeta-mediated NF-kappaB activation. Blood 2001, 97, 3941–3950. [Google Scholar] [CrossRef]
- Chang, W.C.; Shi, G.Y.; Chow, Y.H.; Chang, L.C.; Hau, J.S.; Lin, M.T.; Jen, C.J.; Wing, L.Y.; Wu, H.L. Human plasmin induces a receptor-mediated arachidonate release coupled with G proteins in endothelial cells. Am. J. Physiol. 1993, 264, C271–C281. [Google Scholar] [CrossRef]
- Fujiyoshi, T.; Hirano, K.; Hirano, M.; Nishimura, J.; Takahashi, S.; Kanaide, H. Plasmin induces endothelium-dependent nitric oxide-mediated relaxation in the porcine coronary artery. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 949–954. [Google Scholar] [CrossRef]
- Bauman, K.A.; Wettlaufer, S.H.; Okunishi, K.; Vannella, K.M.; Stoolman, J.S.; Huang, S.K.; Courey, A.J.; White, E.S.; Hogaboam, C.M.; Simon, R.H.; et al. The antifibrotic effects of plasminogen activation occur via prostaglandin E2 synthesis in humans and mice. J. Clin. Invest. 2010, 120, 1950–1960. [Google Scholar] [CrossRef]
- Zhao, X.J.; Larkin, T.M.; Lauver, M.A.; Ahmad, S.; Ducruet, A.F. Tissue plasminogen activator mediates deleterious complement cascade activation in stroke. PLoS ONE 2017, 12, e0180822. [Google Scholar] [CrossRef]
- Quinton, T.M.; Kim, S.; Derian, C.K.; Jin, J.; Kunapuli, S.P. Plasmin-mediated activation of platelets occurs by cleavage of protease-activated receptor 4. J. Biol. Chem. 2004, 279, 18434–18439. [Google Scholar] [CrossRef]
- Syrovets, T.; Simmet, T. Novel aspects and new roles for the serine protease plasmin. Cell Mol. Life Sci. 2004, 61, 873–885. [Google Scholar] [CrossRef]
- Lijnen, H.R. Matrix metalloproteinases and cellular fibrinolytic activity. Biochemistry (Mosc.) 2002, 67, 92–98. [Google Scholar] [CrossRef]
- Pang, P.T.; Teng, H.K.; Zaitsev, E.; Woo, N.T.; Sakata, K.; Zhen, S.; Teng, K.K.; Yung, W.H.; Hempstead, B.L.; Lu, B. Cleavage of proBDNF by tPA/plasmin is essential for long-term hippocampal plasticity. Science 2004, 306, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Yee, J.A.; Yan, L.; Dominguez, J.C.; Allan, E.H.; Martin, T.J. Plasminogen-dependent activation of latent transforming growth factor beta (TGF beta) by growing cultures of osteoblast-like cells. J. Cell Physiol. 1993, 157, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The molecular evolution of genes and proteins: A tale of two serines. Nature 1988, 334, 528–530. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, R.F. Step-by-step evolution of vertebrate blood coagulation. Cold Spring Harb. Symp. Quant. Biol. 2009, 74, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Chana-Munoz, A.; Jendroszek, A.; Sonnichsen, M.; Wang, T.; Ploug, M.; Jensen, J.K.; Andreasen, P.A.; Bendixen, C.; Panitz, F. Origin and diversification of the plasminogen activation system among chordates. BMC Evol. Biol. 2019, 19, 27. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.J.; Zhang, S.C. Demonstration of plasminogen-like protein in amphioxus with implications for the origin of vertebrate liver. Acta Zool. 2006, 87, 141–145. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, S. A kringle-containing protease with plasminogen-like activity in the basal chordate Branchiostoma belcheri. Biosci. Rep. 2009, 29, 385–395. [Google Scholar] [CrossRef]
- Doolittle, R.F. The protochordate Ciona intestinalis has a protein like full-length vertebrate fibrinogen. J. Innate Immun. 2012, 4, 219–222. [Google Scholar] [CrossRef]

{kind=link}
| Target Effects | Properties of Plasmin(ogen) | References |
|---|---|---|
| Proinflammatory Macrophage and Monocyte effects | Interacts with macrophage migration and activation via plasminogen receptors. Promotes cytokine production in macrophages. | [32,35,62] |
| Directly alters gene expression in macrophages by binding plasminogen receptors and enhancing phagocytic capacity, efferocytosis and foam cell formation. | [37,63,64] | |
| Promote macrophage phagocytosis in mice | [36] | |
| Potent chemoattractant of monocytes, induces actin polymerisation. | [65] | |
| Activates 5-lipoxygenase pathway in monocytes and macrophages resulting in the synthesis of proinflammatory leukotrienes. | [66] | |
| Stimulates JAK/STAT signalling in monocytes resulting in MCP-1 release, further promoting monocyte recruitment. | [67] | |
| Increases phagocytic activity of DCs without causing activation. | [38] | |
| Dendritic Cell (DC) effects | This interaction is involved in the chemoattraction of dendritic cells, T- and B cells. Triggers chemotaxis of monocyte derived DCs and a T helper type-1 (Th1) phenotype in CD4+ T cells. | [68] |
| Indirectly promotes neutrophil recruitment by binding to mast cells and stimulating release of leukotrienes | [69] | |
| Induce expression of CCR6-activating chemokine CCL20 in dermis via induction of NF-kB. | [70] | |
| Other inflammatory effects | Stimulates NF-kB and AP-1, resulting in the production of tumor necrosis factor (TNF)-α, interleukin (IL)-1α, IL-1β, and tissue factor. | [71] |
| Activates phospholipase A2 in endothelial cells, releasing arachidonic acid and subsequent production of prostacyclin, enhanced nitric oxide (NO)-mediated vasorelaxation and chemotactic monocyte chemotactic protein (MCP)-1 release. | [39,72,73] | |
| In pulmonary epithelial cells, plasmin induces cyclooxygenase (COX)-2, resulting in the release of antifibrotic prostaglandin E-2 (PGE-2). | [74] | |
| Promotes complement activation. | [17,75] | |
| Binds to platelets via PAR-4 and dose-dependently activate or inhibit platelet activation and aggregation. Interacts with extracellular matrix, endothelial cells, smooth muscle. | [76,77] | |
| Plasmin can activate the Matrix Metalloproteinases, transforming growth factor (TGF)-β, and neurotrophic factors. | [78,79,80] | |
| Inhibition of DC maturation following phagocytosis thereby inducing a tolerogenic phenotype. Reduced migration of DCs to lymph nodes and increase release of TGF-β. Reduction in DC ability to induce allogeneic lymphocyte proliferation. | [38] | |
| Immuno-suppression DC effects | Suppression of proinflammatory cytokines in vivo, reversed by tranexamic acid. Inhibition of plasmin reduces post-surgical infection rates. | [33] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medcalf, R.L.; Keragala, C.B. Fibrinolysis: A Primordial System Linked to the Immune Response. Int. J. Mol. Sci. 2021, 22, 3406. https://doi.org/10.3390/ijms22073406
Medcalf RL, Keragala CB. Fibrinolysis: A Primordial System Linked to the Immune Response. International Journal of Molecular Sciences. 2021; 22(7):3406. https://doi.org/10.3390/ijms22073406
Chicago/Turabian StyleMedcalf, Robert L., and Charithani B. Keragala. 2021. "Fibrinolysis: A Primordial System Linked to the Immune Response" International Journal of Molecular Sciences 22, no. 7: 3406. https://doi.org/10.3390/ijms22073406
APA StyleMedcalf, R. L., & Keragala, C. B. (2021). Fibrinolysis: A Primordial System Linked to the Immune Response. International Journal of Molecular Sciences, 22(7), 3406. https://doi.org/10.3390/ijms22073406