
Atractylodin Produces Antinociceptive Effect through a Long-Lasting TRPA1 Channel Activation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
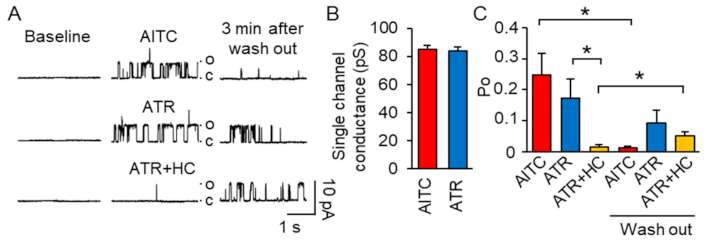
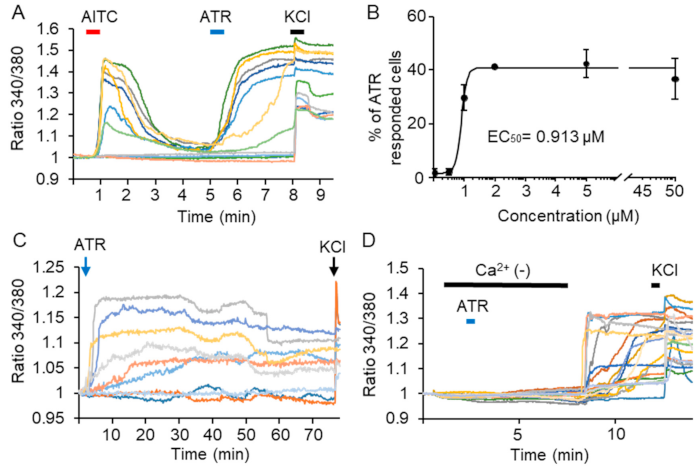
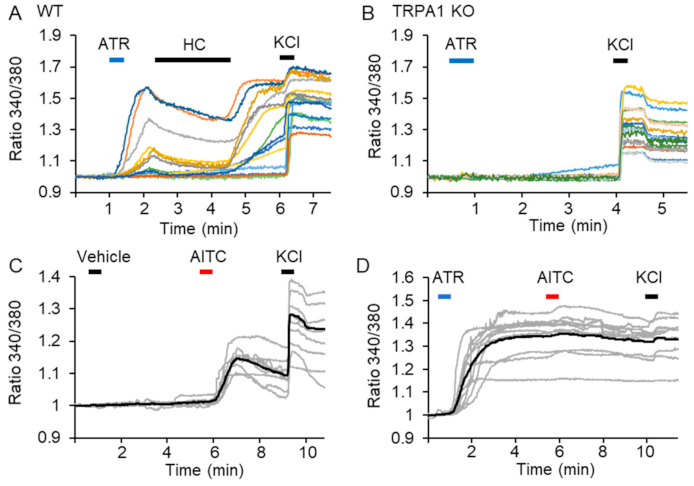
2.1. ATR Induces a Long-Lasting TRPA1 Channel Activation
2.2. Intraplantar Injection of ATR Induces Moderate But Prolonged Nociceptive Behaviors
2.3. Systemic Application of ATR Attenuates AITC-Induced Nociceptive Behaviors Dose-Dependently
2.4. Co-Application of ATR and QX-314 Increases Noxious Heat Threshold
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Primary Culture of the DRG Neurons
4.3. HEK293 Cell Culture and Transfection
4.4. Calcium Imaging Analysis
4.5. Single-Channel Patch-Clamp Analysis
4.6. Assessment of Nociceptive Behaviors
4.7. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATR | Atractylodin |
| AL | Atractylodes lancea De Candolle |
| AITC | Allyl isothiocyanate |
| ANOVA | Analysis of variance |
| CPZ | Capsazepine |
| DRG | Dorsal root ganglion |
| HC | HC-030031 |
| TRPA1 | Transient receptor potential ankyrin-1 |
| TRPV1 | Transient receptor potential vanilloid-1 |
References
- Zhang, W.J.; Zhao, Z.Y.; Chang, L.K.; Cao, Y.; Wang, S.; Kang, C.Z.; Wang, H.Y.; Zhou, L.; Huang, L.Q.; Guo, L.P. Atractylodis Rhizoma: A review of its traditional uses, phytochemistry, pharmacology, toxicology and quality control. J. Ethnopharmacol. 2021, 266, 113415. [Google Scholar] [CrossRef]
- Yu, C.; Xiong, Y.; Chen, D.; Li, Y.; Xu, B.; Lin, Y.; Tang, Z.; Jiang, C.; Wang, L. Ameliorative effects of atractylodin on intestinal inflammation and co-occurring dysmotility in both constipation and diarrhea prominent rats. Korean J. Physiol. Pharmacol. 2017, 21, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nakai, Y.; Kido, T.; Hashimoto, K.; Kase, Y.; Sakakibara, I.; Higuchi, M.; Sasaki, H. Effect of the rhizomes of Atractylodes lancea and its constituents on the delay of gastric emptying. J. Ethnopharmacol. 2003, 84, 51–55. [Google Scholar] [CrossRef]
- Arai, Y.C.; Shimo, K.; Inoue, M.; Sakurai, H.; Ohmichi, Y.; Matsubara, T.; Nishihara, M.; Kawai, T.; Ikemoto, T.; Inoue, S. Integration of a Kampo Medicine, Nijutsuto, and Western Medical Treatment in the Treatment of Long-term Frozen Shoulder Refractory to Western Medical Treatment: A Case Series. J. Evid.-Based Complement. Altern. Med. 2015, 20, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Koike, K.; Yamamoto, Y.; Suzuki, N.; Yamazaki, R.; Yoshikawa, C.; Takano, F.; Sugiura, K.; Inoue, M. Efficacy of porcine placental extract on shoulder stiffness in climacteric women. Climacteric 2013, 16, 447–452. [Google Scholar] [CrossRef]
- Sul, J.U.; Kim, M.K.; Leem, J.; Jo, H.G.; Yoon, S.H.; Kim, J.; Lee, E.J.; Yoo, J.E.; Park, S.J.; Kim, Y.I.; et al. Efficacy and safety of gyejigachulbutang (Gui-Zhi-Jia-Shu-Fu-Tang, Keishikajutsubuto, TJ-18) for knee pain in patients with degenerative knee osteoarthritis: A randomized, placebo-controlled, patient and assessor blinded clinical trial. Trials 2019, 20, 140. [Google Scholar] [CrossRef] [PubMed]
- Noue, M.I.; No, Y.O.; Izukami, H.M. Suppressive Effect of Dai-bofu-to on Collagen-Induced Arthritis. Biol. Pharm. Bull. 2004, 27, 857–862. [Google Scholar]
- Ono, Y.; Ogihara, Y.; Saijo, S.; Iwakura, Y.; Inoue, M. Evaluation of Kampo medicines used to treat rheumatoid arthritis in collagen-induced arthritic and pX transgenic mice. Mod. Rheumatol. 2003, 13, 50–56. [Google Scholar] [CrossRef]
- Satoh, K.; Takano, S.; Kobayashi, T. Keishikajutsubuto (Guizhi-shu-fu-tang) treatment for refractory accumulation of synovial fluid in a patient with pustulotic arthro-osteitis. Fukushima J. Med. Sci. 2007, 53, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Zhao, Y.H.; Xu, J.Y.; Yu, X.Z.; Hu, Y.X.; Zhao, Z.Q. Atractylodin Induces Myosin Light Chain Phosphorylation and Promotes Gastric Emptying through Ghrelin Receptor. Evid.-based Complement. Altern. Med. 2017, 2017, 2186798. [Google Scholar] [CrossRef] [Green Version]
- Chuang, C.H.; Cheng, Y.C.; Lin, S.C.; Lehman, C.W.; Wang, S.P.; Chen, D.Y.; Tsai, S.W.; Lin, C.C. Atractylodin Suppresses Dendritic Cell Maturation and Ameliorates Collagen-Induced Arthritis in a Mouse Model. J. Agric. Food Chem. 2019, 67, 6773–6784. [Google Scholar] [CrossRef]
- Kotawong, K.; Chaijaroenkul, W.; Muhamad, P.; Na-Bangchang, K. Cytotoxic activities and effects of atractylodin and β-eudesmol on the cell cycle arrest and apoptosis on cholangiocarcinoma cell line. J. Pharmacol. Sci. 2018, 136, 51–56. [Google Scholar] [CrossRef]
- Lyu, Z.; Ji, X.; Chen, G.; An, B. Atractylodin ameliorates lipopolysaccharide and D-galactosamine-induced acute liver failure via the suppression of inflammation and oxidative stress. Int. Immunopharmacol. 2019, 72, 348–357. [Google Scholar] [CrossRef]
- Kádková, A.; Synytsya, V.; Krusek, J.; Zímová, L.; Vlachová, V. Molecular basis of TRPA1 regulation in nociceptive neurons. A review. Physiol. Res. 2017, 66, 425–439. [Google Scholar] [CrossRef]
- Meents, J.E.; Ciotu, C.I.; Fischer, M.J.M. Trpa1: A molecular view. J. Neurophysiol. 2019, 121, 427–443. [Google Scholar] [CrossRef]
- Macpherson, L.J.; Dubin, A.E.; Evans, M.J.; Marr, F.; Schultz, P.G.; Cravatt, B.F.; Patapoutian, A. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. Nature 2007, 445, 541–545. [Google Scholar] [CrossRef]
- Meents, J.E.; Fischer, M.J.M.; McNaughton, P.A. Agonist-induced sensitisation of the irritant receptor ion channel TRPA1. J. Physiol. 2016, 594, 6643–6660. [Google Scholar] [CrossRef]
- Talavera, K.; Startek, J.B.; Alvarez-Collazo, J.; Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Naert, R.; Nilius, B. Mammalian Transient Receptor Potential TRPA1 Channels: From Structure to Disease. Physiol. Rev. 2020, 100, 725–803. [Google Scholar] [CrossRef]
- Ton, H.T.; Phan, T.X.; Abramyan, A.M.; Shi, L.; Ahern, G.P. Identification of a putative binding site critical for general anesthetic activation of TRPA1. Proc. Natl. Acad. Sci. USA 2017, 114, 3762–3767. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Reese, R.; Vu, S.; Rougé, L.; Shields, S.D.; Kakiuchi-Kiyota, S.; Chen, H.; Johnson, K.; Shi, Y.P.; Chernov-Rogan, T.; et al. A Non-covalent Ligand Reveals Biased Agonism of the TRPA1 Ion Channel. Neuron 2020, 109, 273–284. [Google Scholar] [CrossRef]
- Chen, J.; Kang, D.; Xu, J.; Lake, M.; Hogan, J.O.; Sun, C.; Walter, K.; Yao, B.; Kim, D. Species differences and molecular determinant of TRPA1 cold sensitivity. Nat. Commun. 2013, 4, 3501. [Google Scholar] [CrossRef]
- Kistner, K.; Siklosi, N.; Babes, A.; Khalil, M.; Selescu, T.; Zimmermann, K.; Wirtz, S.; Becker, C.; Neurath, M.F.; Reeh, P.W.; et al. Systemic desensitization through TRPA1 channels by capsazepine and mustard oil—A novel strategy against inflammation and pain. Sci. Rep. 2016, 6, 28621. [Google Scholar] [CrossRef] [Green Version]
- Binshtok, A.M.; Bean, B.P.; Woolf, C.J. Inhibition of nociceptors by TRPV1-mediated entry of impermeant sodium channel blockers. Nature 2007, 449, 607–610. [Google Scholar] [CrossRef]
- Brenneis, C.; Kistner, K.; Puopolo, M.; Segal, D.; Roberson, D.; Sisignano, M.; Labocha, S.; Ferreirós, N.; Strominger, A.; Cobos, E.J.; et al. Phenotyping the function of TRPV1-expressing sensory neurons by targeted axonal silencing. J. Neurosci. 2013, 33, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Kim, D.; Bianchi, B.R.; Cavanaugh, E.J.; Faltynek, C.R.; Kym, P.R.; Reilly, R.M. Pore dilation occurs in TRPA1 but not in TRPM8 channels. Mol. Pain 2009, 5, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Hiura, A. Comparison of the transport of QX-314 through TRPA1, TRPM8, and TRPV1 channels. J. Pain Res. 2013, 6, 223. [Google Scholar] [CrossRef] [Green Version]
- Peterlin, Z.; Chesler, A.; Firestein, S. A Painful Trp Can Be a Bonding Experience. Neuron 2007, 53, 635–638. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Lin King, J.V.; Paulsen, C.E.; Cheng, Y.; Julius, D. Mechanisms governing irritant-evoked activation and calcium modulation of TRPA1. bioRxiv 2019. [Google Scholar] [CrossRef]
- Nakamura, Y.; Une, Y.; Miyano, K.; Abe, H.; Hisaoka, K.; Morioka, N.; Nakata, Y. Activation of transient receptor potential ankyrin 1 evokes nociception through substance P release from primary sensory neurons. J. Neurochem. 2012, 120, 1036–1047. [Google Scholar] [CrossRef]
- Gentry, C.; Stoakley, N.; Andersson, D.A.; Bevan, S. The roles of iPLA2, TRPM8 and TRPA1 in chemically induced cold hypersensitivity. Mol. Pain 2010, 6, 1744–8069. [Google Scholar] [CrossRef] [Green Version]
- Salazar, H.; Llorente, I.; Jara-Oseguera, A.; García-Villegas, R.; Munari, M.; Gordon, S.E.; Islas, L.D.; Rosenbaum, T. A single N-terminal cysteine in TRPV1 determines activation by pungent compounds from onion and garlic. Nat. Neurosci. 2008, 11, 255–261. [Google Scholar] [CrossRef]
- Raisinghani, M.; Zhong, L.; Jeffry, J.A.; Bishnoi, M.; Pabbidi, R.M.; Pimentel, F.; Cao, D.S.; Evans, M.S.; Premkumar, L.S. Activation characteristics of transient receptor potential ankyrin 1 and its role in nociception. Am. J. Physiol. Cell Physiol. 2011, 301, C587–C600. [Google Scholar] [CrossRef] [Green Version]
- Karashima, Y.; Damann, N.; Prenen, J.; Talavera, K.; Segal, A.; Voets, T.; Nilius, B. Bimodal action of menthol on the transient receptor potential channel TRPA1. J. Neurosci. 2007, 27, 9874–9884. [Google Scholar] [CrossRef]
- Xu, H.; Blair, N.T.; Clapham, D.E. Camphor activates and strongly desensitizes the transient receptor potential vanilloid subtype 1 channel in a vanilloid-independent mechanism. J. Neurosci. 2005, 25, 8924–8937. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with Aδ/C-fibers and colocalization with Trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef]
- Ongun, S.; Sarkisian, A.; McKemy, D.D. Selective cold pain inhibition by targeted block of TRPM8-expressing neurons with quaternary lidocaine derivative QX-314. Commun. Biol. 2018, 1, 53. [Google Scholar] [CrossRef]
- Bautista, D.M.; Jordt, S.E.; Nikai, T.; Tsuruda, P.R.; Read, A.J.; Poblete, J.; Yamoah, E.N.; Basbaum, A.I.; Julius, D. TRPA1 Mediates the Inflammatory Actions of Environmental Irritants and Proalgesic Agents. Cell 2006, 124, 1269–1282. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kobayashi, K.; Kogure, Y.; Yamanaka, H.; Yamamoto, S.; Yagi, H.; Noguchi, K.; Dai, Y. Negative regulation of TRPA1 by AMPK in primary sensory neurons as a potential mechanism of painful diabetic neuropathy. Diabetes 2018, 67, 98–109. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanda, H.; Yang, Y.; Duan, S.; Kogure, Y.; Wang, S.; Iwaoka, E.; Ishikawa, M.; Takeda, S.; Sonoda, H.; Mizuta, K.; et al. Atractylodin Produces Antinociceptive Effect through a Long-Lasting TRPA1 Channel Activation. Int. J. Mol. Sci. 2021, 22, 3614. https://doi.org/10.3390/ijms22073614
Kanda H, Yang Y, Duan S, Kogure Y, Wang S, Iwaoka E, Ishikawa M, Takeda S, Sonoda H, Mizuta K, et al. Atractylodin Produces Antinociceptive Effect through a Long-Lasting TRPA1 Channel Activation. International Journal of Molecular Sciences. 2021; 22(7):3614. https://doi.org/10.3390/ijms22073614
Chicago/Turabian StyleKanda, Hirosato, Yanjing Yang, Shaoqi Duan, Yoko Kogure, Shenglan Wang, Emiko Iwaoka, Miku Ishikawa, Saki Takeda, Hidemi Sonoda, Kyoka Mizuta, and et al. 2021. "Atractylodin Produces Antinociceptive Effect through a Long-Lasting TRPA1 Channel Activation" International Journal of Molecular Sciences 22, no. 7: 3614. https://doi.org/10.3390/ijms22073614
APA StyleKanda, H., Yang, Y., Duan, S., Kogure, Y., Wang, S., Iwaoka, E., Ishikawa, M., Takeda, S., Sonoda, H., Mizuta, K., Aoki, S., Yamamoto, S., Noguchi, K., & Dai, Y. (2021). Atractylodin Produces Antinociceptive Effect through a Long-Lasting TRPA1 Channel Activation. International Journal of Molecular Sciences, 22(7), 3614. https://doi.org/10.3390/ijms22073614