
Thapsigargin-Stimulated LAD2 Human Mast Cell Line Is a Potent Cellular Adjuvant for the Maturation of Monocyte-Derived Dendritic Cells for Adoptive Cellular Immunotherapy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
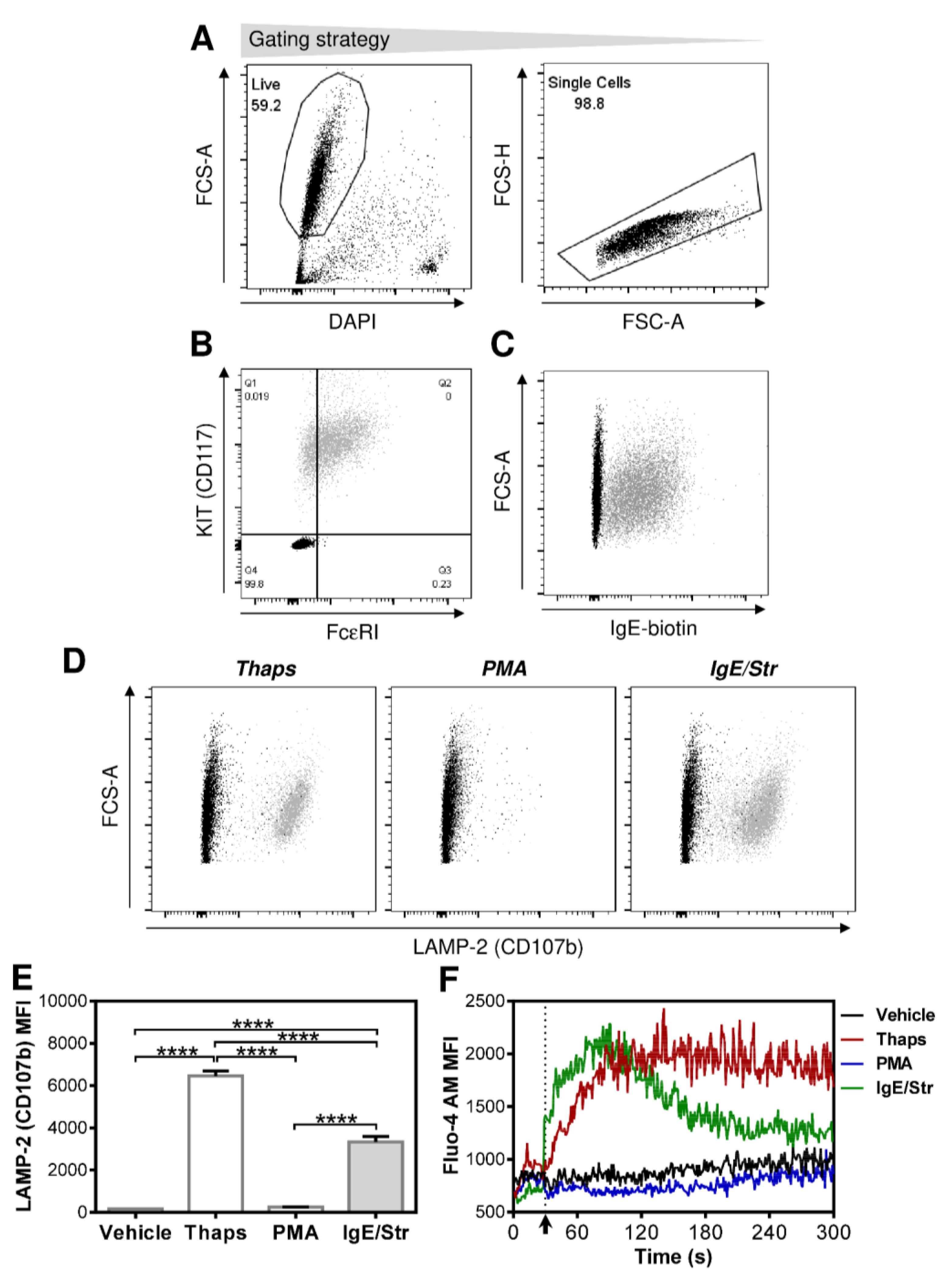
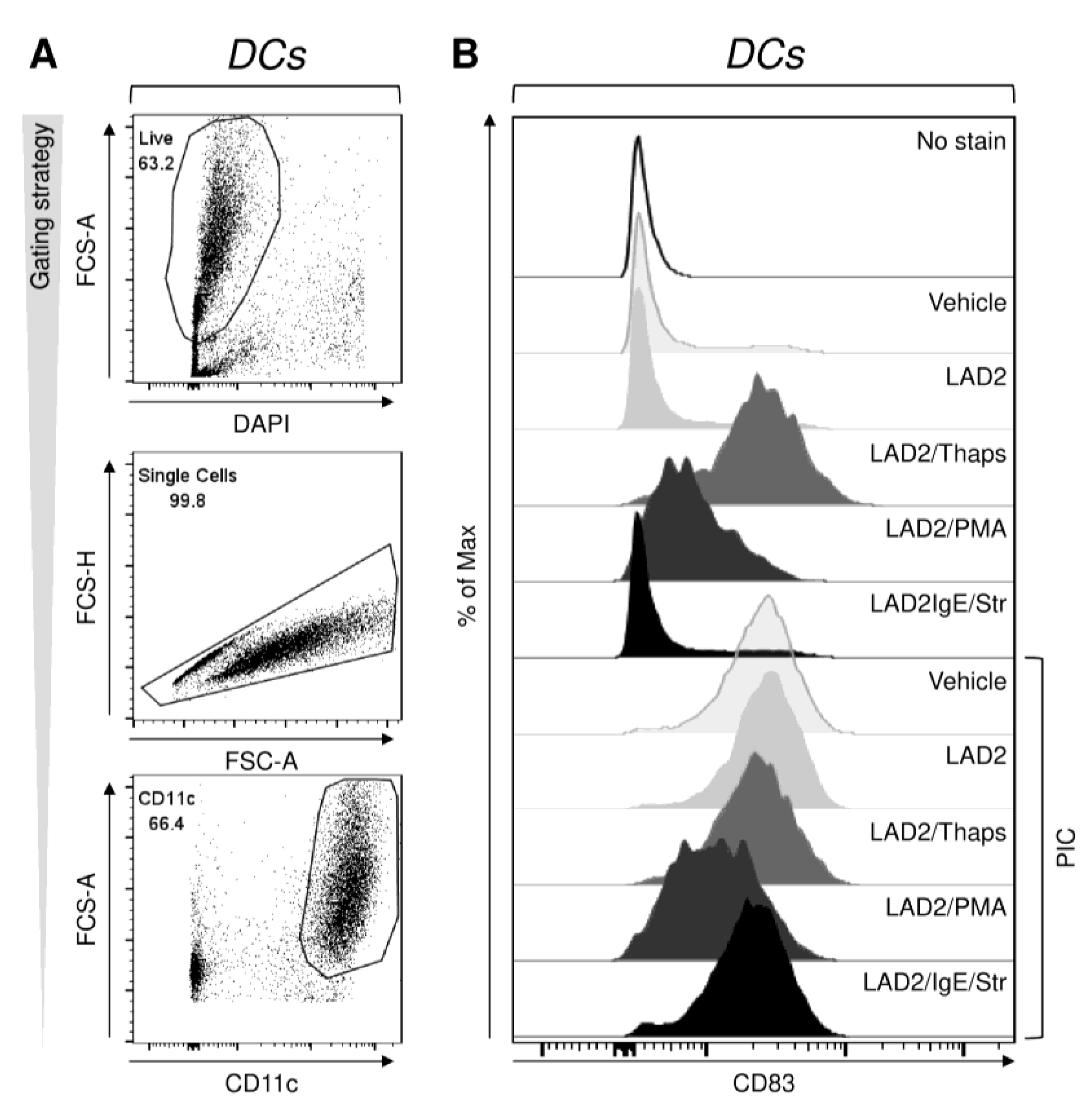
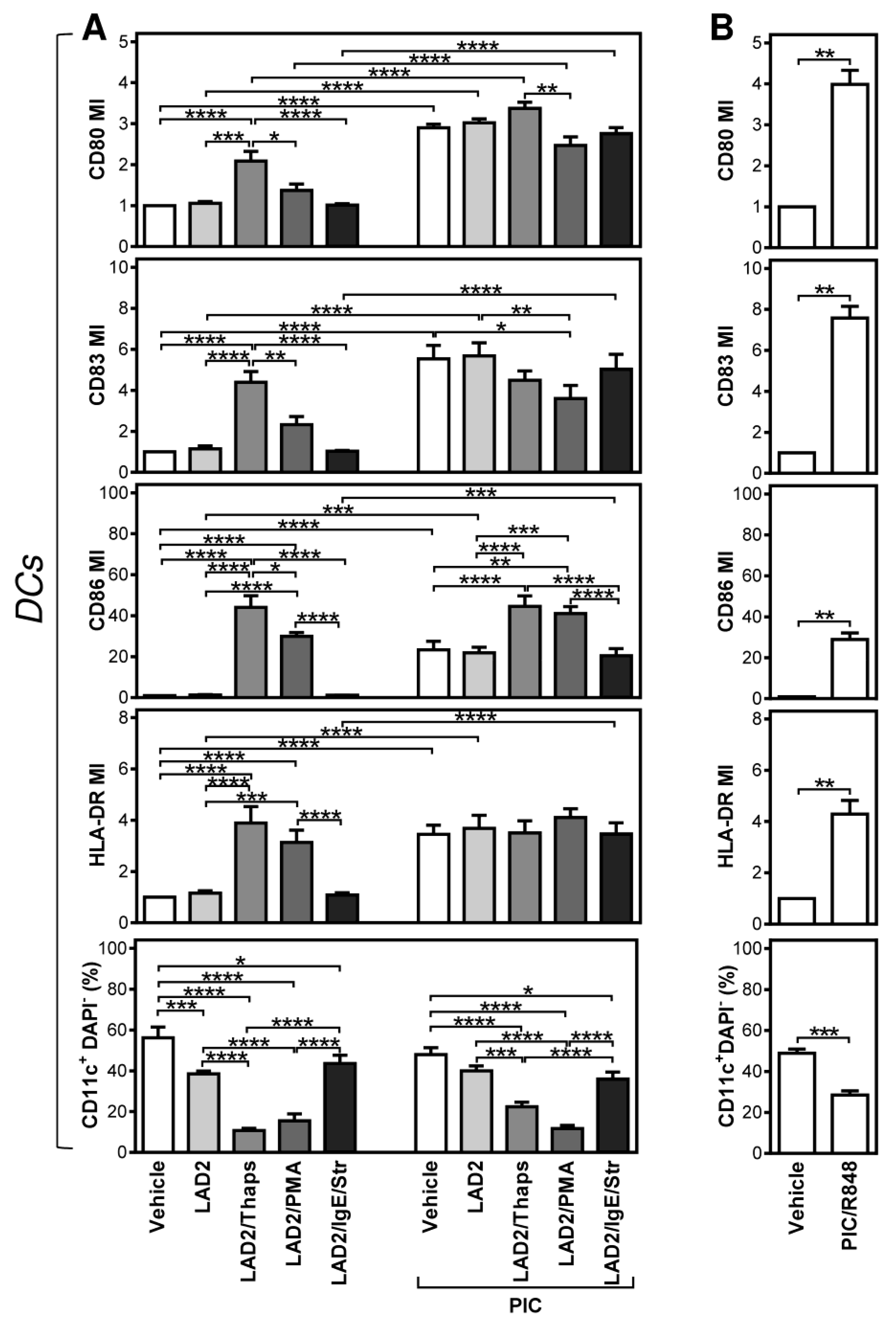
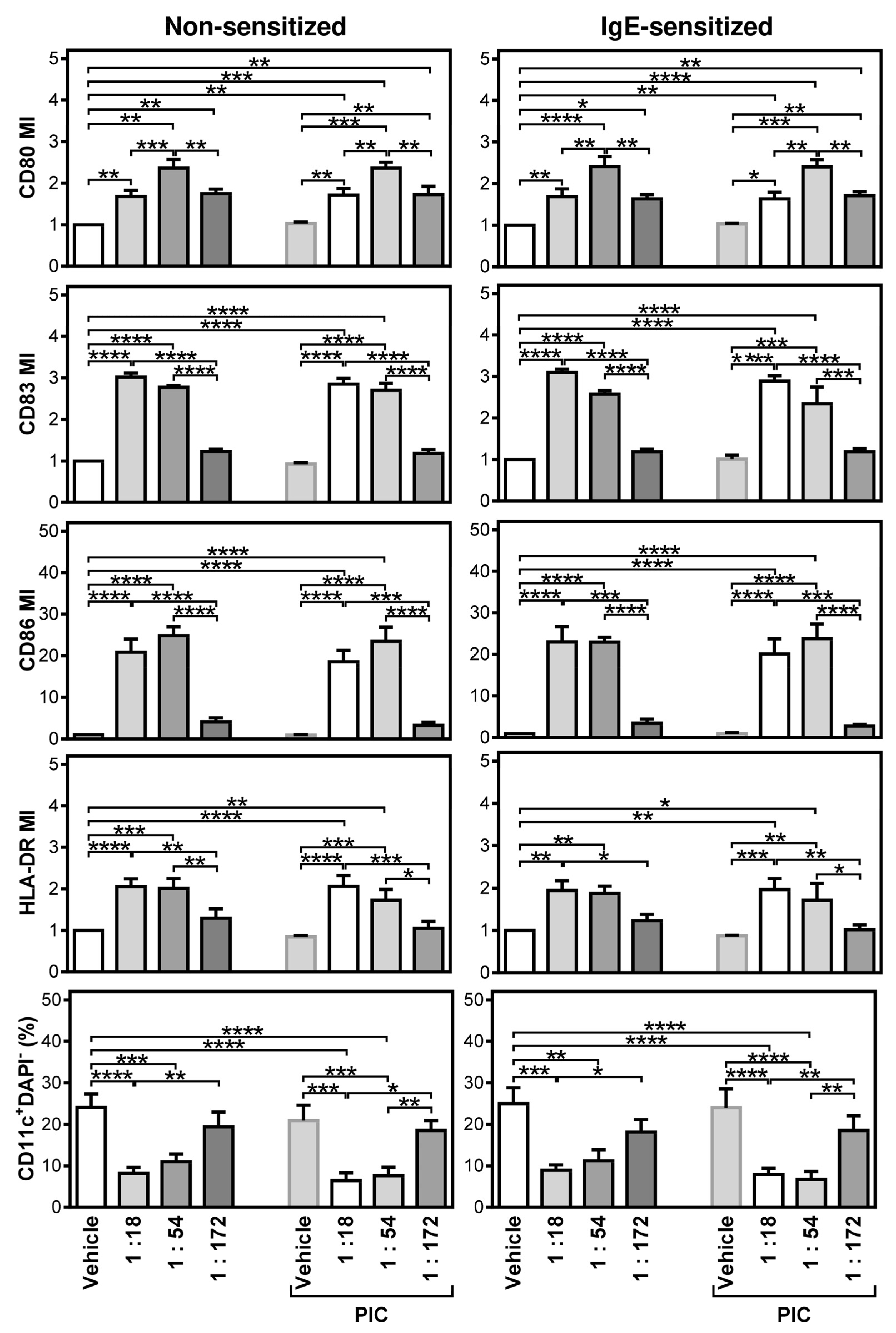
2.1. Differentially Stimulated LAD2 MCs Induce and Modulate DC Maturation
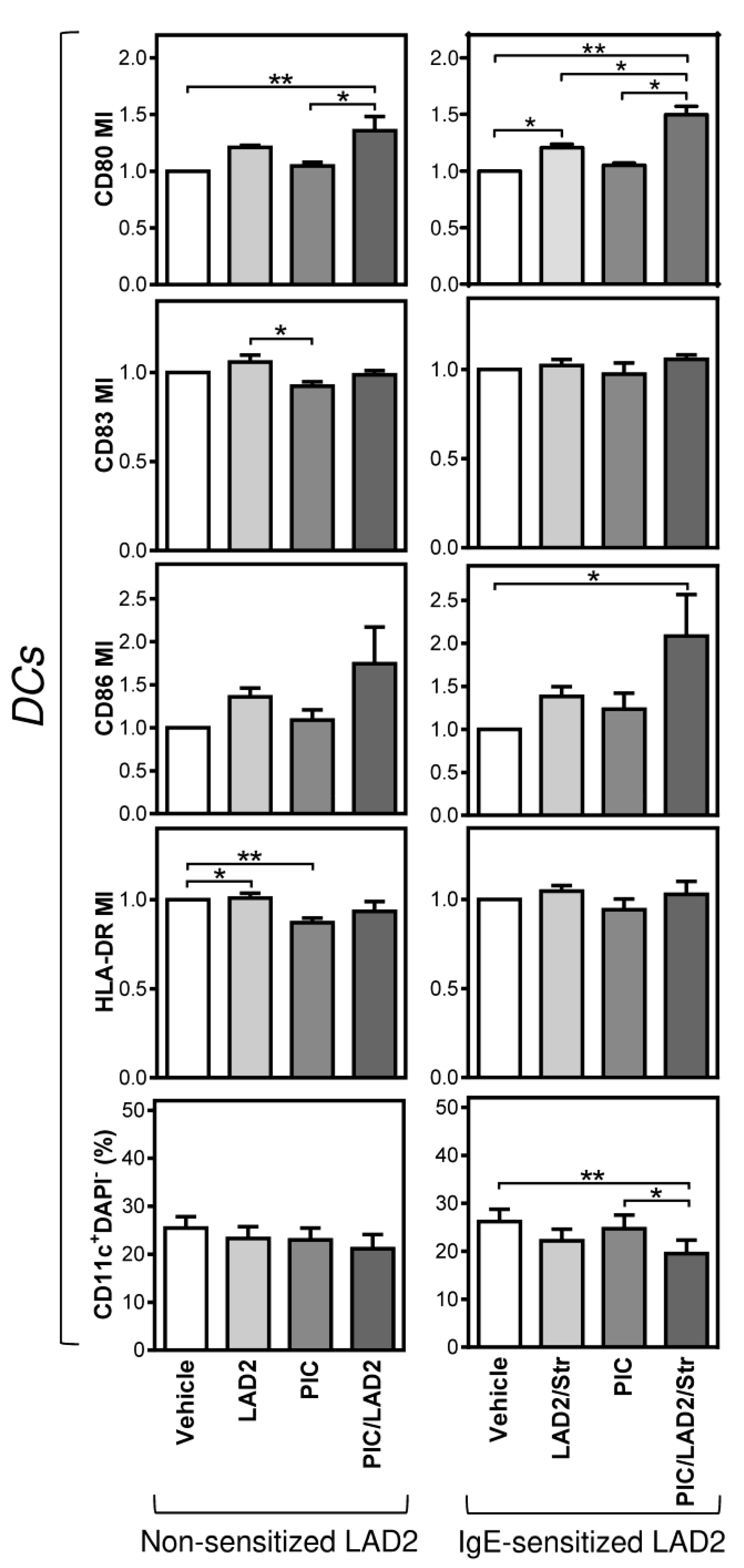
2.2. FcεRI-Stimulated LAD2 MCs Have Limited Potential to Promote Maturation of Maturation-Refractory iDCs
2.3. The Thapsigargin-Stimulated LAD2 MCs Induce Maturation in the Maturation-Refractory iDCs
3. Discussion
4. Materials and Methods
4.1. Specimens
4.2. LAD2 MC Stimulation and Analyses
4.3. Preparation of iDCs
4.4. DC Maturation and Analyses
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Gilfillan, A.M.; Austin, S.J.; Metcalfe, D.D. Mast Cell Biology: Introduction and Overview. Adv. Exp. Med. Biol. 2011, 716, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Valent, P.; Akin, C.; Nedoszytko, B.; Bonadonna, P.; Hartmann, K.; Niedoszytko, M.; Brockow, K.; Siebenhaar, F.; Triggiani, M.; Arock, M.; et al. Diagnosis, Classification and Management of Mast Cell Activation Syndromes (MCAS) in the Era of Personalized Medicine. Int. J. Mol. Sci. 2020, 21, 9030. [Google Scholar] [CrossRef] [PubMed]
- Komi, D.E.A.; Redegeld, F.A. Role of Mast Cells in Shaping the Tumor Microenvironment. Clin. Rev. Allergy Immunol. 2020, 58, 313–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varricchi, G.; Galdiero, M.R.; Loffredo, S.; Marone, G.; Iannone, R.; Marone, G.; Granata, F. Are Mast Cells MASTers in Cancer? Front. Immunol. 2017, 8, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derakhshani, A.; Vahidian, F.; Alihasanzadeh, M.; Mokhtarzadeh, A.; Lotfi Nezhad, P.; Baradaran, B. Mast Cells: A Double-edged Sword in Cancer. Immunol. Lett. 2019, 209, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Carroll-Portillo, A.; Cannon, J.L.; te Riet, J.; Holmes, A.; Kawakami, Y.; Kawakami, T.; Cambi, A.; Lidke, D.S. Mast Cells and Dendritic Cells Form Synapses That Facilitate Antigen Transfer for T Cell Activation. J. Cell. Biol. 2015, 210, 851–864. [Google Scholar] [CrossRef]
- Dudeck, J.; Medyukhina, A.; Frobel, J.; Svensson, C.M.; Kotrba, J.; Gerlach, M.; Gradtke, A.C.; Schroder, B.; Speier, S.; Figge, M.T.; et al. Mast Cells Acquire MHCII from Dendritic Cells during Skin Inflammation. J. Exp. Med. 2017, 214, 3791–3811. [Google Scholar] [CrossRef] [PubMed]
- de Vries, V.C.; Pino-Lagos, K.; Nowak, E.C.; Bennett, K.A.; Oliva, C.; Noelle, R.J. Mast Cells Condition Dendritic Cells to Mediate Allograft Tolerance. Immunity 2011, 35, 550–561. [Google Scholar] [CrossRef] [Green Version]
- Dudeck, J.; Ghouse, S.M.; Lehmann, C.H.; Hoppe, A.; Schubert, N.; Nedospasov, S.A.; Dudziak, D.; Dudeck, A. Mast-Cell-Derived TNF Amplifies CD8+ Dendritic Cell Functionality and CD8+ T Cell Priming. Cell Rep. 2015, 13, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Dudeck, A.; Suender, C.A.; Kostka, S.L.; von Stebut, E.; Maurer, M. Mast Cells Promote Th1 and Th17 Responses by Modulating Dendritic Cell Maturation and Function. Eur. J. Immunol. 2011, 41, 1883–1893. [Google Scholar] [CrossRef]
- Wculek, S.K.; Cueto, F.J.; Mujal, A.M.; Melero, I.; Krummel, M.F.; Sancho, D. Dendritic Cells in Cancer Immunology and Immunotherapy. Nat. Rev. Immunol. 2020, 20, 7–24. [Google Scholar] [CrossRef]
- Huber, A.; Dammeijer, F.; Aerts, J.; Vroman, H. Current State of Dendritic Cell-Based Immunotherapy: Opportunities for in vitro Antigen Loading of Different DC Subsets? Front. Immunol. 2018, 9, 2804. [Google Scholar] [CrossRef]
- Calmeiro, J.; Carrascal, M.A.; Tavares, A.R.; Ferreira, D.A.; Gomes, C.; Falcao, A.; Cruz, M.T.; Neves, B.M. Dendritic Cell Vaccines for Cancer Immunotherapy: The Role of Human Conventional Type 1 Dendritic Cells. Pharmaceutics 2020, 12, 158. [Google Scholar] [CrossRef] [Green Version]
- Gardner, A.; de Mingo Pulido, A.; Ruffell, B. Dendritic Cells and Their Role in Immunotherapy. Front. Immunol. 2020, 11, 924. [Google Scholar] [CrossRef] [PubMed]
- Calmeiro, J.; Mendes, L.; Duarte, I.F.; Leitao, C.; Tavares, A.R.; Ferreira, D.A.; Gomes, C.; Serra, J.; Falcao, A.; Cruz, M.T.; et al. In-Depth Analysis of the Impact of Different Serum-Free Media on the Production of Clinical Grade Dendritic Cells for Cancer Immunotherapy. Front. Immunol. 2020, 11, 593363. [Google Scholar] [CrossRef]
- Lovgren, T.; Sarhan, D.; Truxova, I.; Choudhary, B.; Maas, R.; Melief, J.; Nystrom, M.; Edback, U.; Vermeij, R.; Scurti, G.; et al. Enhanced Stimulation of Human Tumor-specific T Cells by Dendritic Cells Matured in the Presence of Interferon-gamma and Multiple Toll-like Receptor Agonists. Cancer Immunol. Immunother. CII 2017, 66, 1333–1344. [Google Scholar] [CrossRef] [Green Version]
- Ten Brinke, A.; Karsten, M.L.; Dieker, M.C.; Zwaginga, J.J.; van Ham, S.M. The Clinical Grade Maturation Cocktail Monophosphoryl Lipid A Plus IFNgamma Generates Monocyte-derived Dendritic Cells with the Capacity to Migrate and Induce Th1 Polarization. Vaccine 2007, 25, 7145–7152. [Google Scholar] [CrossRef] [PubMed]
- Cechim, G.; Chies, J.A.B. In Vitro Generation of Human Monocyte-derived Dendritic Cells Methodological Aspects in a Comprehensive Review. An. Acad. Bras. Cienc. 2019, 91. [Google Scholar] [CrossRef]
- Kirshenbaum, A.S.; Akin, C.; Wu, Y.; Rottem, M.; Goff, J.P.; Beaven, M.A.; Rao, V.K.; Metcalfe, D.D. Characterization of Novel Stem Cell Factor Responsive Human Mast Cell Lines LAD 1 and 2 Established from a Patient with Mast Cell Sarcoma/Leukemia; Activation Following Aggregation of FcεRI or FcγRI. Leuk. Res. 2003, 27, 677–682. [Google Scholar] [CrossRef]
- Rouas, R.; Lewalle, P.; El Ouriaghli, F.; Nowak, B.; Duvillier, H.; Martiat, P. Poly(I:C) Used for Human Dendritic Cell Maturation Preserves Their Ability to Secondarily Secrete Bioactive IL-12. Int. Immunol. 2004, 16, 767–773. [Google Scholar] [CrossRef] [Green Version]
- Fucikova, J.; Rozkova, D.; Ulcova, H.; Budinsky, V.; Sochorova, K.; Pokorna, K.; Bartunkova, J.; Spisek, R. Poly I: C-activated Dendritic Cells That Were Generated in CellGro for Use in Cancer Immunotherapy Trials. J. Transl. Med. 2011, 9, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, G.A.; Varhol, R.J. Kinetics of Thapsigargin-Ca2+-ATPase (Sarcoplasmic reticulum) Interaction Reveals a Two-step Binding Mechanism and Picomolar Inhibition. J. Biol. Chem. 1995, 270, 11731–11734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kijima, Y.; Ogunbunmi, E.; Fleischer, S. Drug Action of Thapsigargin on the Ca2+ Pump Protein of Sarcoplasmic reticulum. J. Biol. Chem. 1991, 266, 22912–22918. [Google Scholar] [CrossRef]
- Lytton, J.; Westlin, M.; Hanley, M.R. Thapsigargin Inhibits the Sarcoplasmic or Endoplasmic reticulum Ca-ATPase Family of Calcium Pumps. J. Biol. Chem. 1991, 266, 17067–17071. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P.; Francis, G.; Becker, K. Phorbol Esters: Structure, Biological Activity, and Toxicity in Animals. Int. J. Toxicol. 2007, 26, 279–288. [Google Scholar] [CrossRef]
- Smrz, D.; Cruse, G.; Beaven, M.A.; Kirshenbaum, A.; Metcalfe, D.D.; Gilfillan, A.M. Rictor Negatively Regulates High-Affinity Receptors for IgE-Induced Mast Cell Degranulation. J. Immunol. 2014, 193, 5924–5932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruse, G.; Gilfillan, A.M.; Smrz, D. Flow Cytometry-based Monitoring of Mast Cell Activation. Methods Mol. Biol. 2015, 1220, 365–379. [Google Scholar] [CrossRef]
- Cruse, G.; Beaven, M.A.; Ashmole, I.; Bradding, P.; Gilfillan, A.M.; Metcalfe, D.D. A Truncated Splice-variant of the FcepsilonRIbeta Receptor Subunit is Critical for Microtubule Formation and Degranulation in Mast Cells. Immunity 2013, 38, 906–917. [Google Scholar] [CrossRef] [Green Version]
- Fucikova, J.; Podrazil, M.; Jarolim, L.; Bilkova, P.; Hensler, M.; Becht, E.; Gasova, Z.; Klouckova, J.; Kayserova, J.; Horvath, R.; et al. Phase I/II Trial of Dendritic Cell-based Active Cellular Immunotherapy with DCVAC/PCa in Patients with Rising PSA after Primary Prostatectomy or Salvage Radiotherapy for the Treatment of Prostate Cancer. Cancer Immunol. Immunother. CII 2018, 67, 89–100. [Google Scholar] [CrossRef]
- Podrazil, M.; Horvath, R.; Becht, E.; Rozkova, D.; Bilkova, P.; Sochorova, K.; Hromadkova, H.; Kayserova, J.; Vavrova, K.; Lastovicka, J.; et al. Phase I/II Clinical Trial of Dendritic-cell Based Immunotherapy (DCVAC/PCa) Combined with Chemotherapy in Patients with Metastatic, Castration-resistant Prostate Cancer. Oncotarget 2015, 6, 18192–18205. [Google Scholar] [CrossRef] [Green Version]
- Jin, P.; Han, T.H.; Ren, J.; Saunders, S.; Wang, E.; Marincola, F.M.; Stroncek, D.F. Molecular Signatures of Maturing Dendritic Cells: Implications for Testing the Quality of Dendritic Cell Therapies. J. Transl. Med. 2010, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pufnock, J.S.; Cigal, M.; Rolczynski, L.S.; Andersen-Nissen, E.; Wolfl, M.; McElrath, M.J.; Greenberg, P.D. Priming CD8+ T Cells with Dendritic Cells Matured Using TLR4 and TLR7/8 Ligands Together Enhances Generation of CD8+ T Cells Retaining CD28. Blood 2011, 117, 6542–6551. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Yang, D.; Trivett, A.; Meyer, T.J.; Oppenheim, J.J. HMGN1 and R848 Synergistically Activate Dendritic Cells Using Multiple Signaling Pathways. Front. Immunol. 2018, 9, 2982. [Google Scholar] [CrossRef]
- Nie, Y.; Yang, D.; Trivett, A.; Han, Z.; Xin, H.; Chen, X.; Oppenheim, J.J. Development of a Curative Therapeutic Vaccine (TheraVac) for the Treatment of Large Established Tumors. Sci. Rep. 2017, 7, 14186. [Google Scholar] [CrossRef] [Green Version]
- Loudovaris, M.; Hansen, M.; Suen, Y.; Lee, S.M.; Casing, P.; Bender, J.G. Differential Effects of Autologous Serum on CD34+ or Monocyte-derived Dendritic Cells. J. Hematotherapy Stem Cell Res. 2001, 10, 569–578. [Google Scholar] [CrossRef]
- Ratta, M.; Fagnoni, F.; Curti, A.; Vescovini, R.; Sansoni, P.; Oliviero, B.; Fogli, M.; Ferri, E.; Della Cuna, G.R.; Tura, S.; et al. Dendritic Cells are Functionally Defective in Multiple Myeloma: The Role of Interleukin-6. Blood 2002, 100, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.H.; Lee, H.J.; Lee, Y.K.; Yang, D.H.; Kim, H.J.; Rhee, J.H.; Emmrich, F.; Lee, J.J. A Phase I Clinical Study of Autologous Dendritic Cell Therapy in Patients with Relapsed or Refractory Multiple Myeloma. Oncotarget 2017, 8, 41538–41548. [Google Scholar] [CrossRef]
- Moon, T.C.; Befus, A.D.; Kulka, M. Mast Cell Mediators: Their Differential Release and the Secretory Pathways Involved. Front. Immunol. 2014, 5, 569. [Google Scholar] [CrossRef] [Green Version]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast Cells as Sources of Cytokines, Chemokines, and Growth Factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Zhao, Y.; Wang, X.; Chen, N.; Mao, F.; Teng, Y.; Wang, T.; Peng, L.; Zhang, J.; Cheng, P.; et al. Increased Intratumoral Mast Cells Foster Immune Suppression and Gastric Cancer Progression through TNF-alpha-PD-L1 Pathway. J. Immunother. Cancer 2019, 7, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somasundaram, R.; Connelly, T.; Choi, R.; Choi, H.; Samarkina, A.; Li, L.; Gregorio, E.; Chen, Y.; Thakur, R.; Abdel-Mohsen, M.; et al. Tumor-infiltrating Mast Cells are Associated with Resistance to Anti-PD-1 Therapy. Nat. Commun. 2021, 12, 346. [Google Scholar] [CrossRef]
- Rajput, A.B.; Turbin, D.A.; Cheang, M.C.; Voduc, D.K.; Leung, S.; Gelmon, K.A.; Gilks, C.B.; Huntsman, D.G. Stromal Mast Cells in Invasive Breast Cancer are a Marker of Favourable Prognosis: A Study of 4,444 Cases. Breast Cancer Res. Treat. 2008, 107, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleischmann, A.; Schlomm, T.; Köllermann, J.; Sekulic, N.; Huland, H.; Mirlacher, M.; Sauter, G.; Simon, R.; Erbersdobler, A. Immunological Microenvironment in Prostate Cancer: High Mast Cell Densities are Associated with Favorable Tumor Characteristics and Good Prognosis. Prostate 2009, 69, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Dzopalic, T.; Rajkovic, I.; Dragicevic, A.; Colic, M. The Response of Human Dendritic Cells to Co-ligation of Pattern-recognition Receptors. Immunol. Res. 2012, 52, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.L. Endotoxin Detection and Control in Pharma, Limulus, and Mammalian Systems; Springer: Cham, Switzeland, 2019. [Google Scholar]
- Casella, C.R.; Mitchell, T.C. Putting Endotoxin to Work for Us: Monophosphoryl Lipid A as a Safe and Effective Vaccine Adjuvant. Cell. Mol. Life Sci. CMLS 2008, 65, 3231–3240. [Google Scholar] [CrossRef] [Green Version]
- Varricchi, G.; Rossi, F.W.; Galdiero, M.R.; Granata, F.; Criscuolo, G.; Spadaro, G.; de Paulis, A.; Marone, G. Physiological Roles of Mast Cells: Collegium Internationale Allergologicum Update 2019. Int. Arch. Allergy Immunol. 2019, 179, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Porebski, G.; Kwiecien, K.; Pawica, M.; Kwitniewski, M. Mas-Related G Protein-Coupled Receptor-X2 (MRGPRX2) in Drug Hypersensitivity Reactions. Front. Immunol. 2018, 9, 3027. [Google Scholar] [CrossRef]
- Willows, S.; Kulka, M. Harnessing the Power of Mast Cells in unconventional Immunotherapy Strategies and Vaccine Adjuvants. Cells 2020, 9, 2713. [Google Scholar] [CrossRef]
- Metz, M.; Piliponsky, A.M.; Chen, C.C.; Lammel, V.; Abrink, M.; Pejler, G.; Tsai, M.; Galli, S.J. Mast Cells Can Enhance Resistance to Snake and Honeybee Venoms. Science 2006, 313, 526–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piconese, S.; Gri, G.; Tripodo, C.; Musio, S.; Gorzanelli, A.; Frossi, B.; Pedotti, R.; Pucillo, C.E.; Colombo, M.P. Mast Cells Counteract Regulatory T-cell Suppression through Interleukin-6 and OX40/OX40L Axis toward Th17-cell Differentiation. Blood 2009, 114, 2639–2648. [Google Scholar] [CrossRef]
- Foufelle, F.; Fromenty, B. Role of Endoplasmic Reticulum Stress in Drug-induced Toxicity. Pharmacol. Res. Perspect. 2016, 4, e00211. [Google Scholar] [CrossRef]
- Sehgal, P.; Szalai, P.; Olesen, C.; Praetorius, H.A.; Nissen, P.; Christensen, S.B.; Engedal, N.; Moller, J.V. Inhibition of the Sarco/Endoplasmic Reticulum (ER) Ca2+-ATPase by Thapsigargin Analogs Induces Cell Death via ER Ca2+ Depletion and the Unfolded Protein Response. J. Biol. Chem. 2017, 292, 19656–19673. [Google Scholar] [CrossRef] [Green Version]
- Katsoulis-Dimitriou, K.; Kotrba, J.; Voss, M.; Dudeck, J.; Dudeck, A. Mast Cell Functions Linking Innate Sensing to Adaptive Immunity. Cells 2020, 9, 2538. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Lantz, C.S.; Oettgen, H.C.; Katona, I.M.; Fleming, T.; Miyajima, I.; Kinet, J.P.; Galli, S.J. IgE Enhances Mouse Mast Cell Fc(epsilon)RI Expression in vitro and in vivo: Evidence for a Novel Amplification Mechanism in IgE-dependent Reactions. J. Exp. Med. 1997, 185, 663–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, M.; Sayama, K.; Yano, K.; Lantz, C.S.; Noben-Trauth, N.; Ra, C.; Costa, J.J.; Galli, S.J. IgE Enhances Fc Epsilon Receptor I Expression and IgE-dependent Release of Histamine and Lipid Mediators from Human Umbilical Cord Blood-derived Mast Cells: Synergistic Effect of IL-4 and IgE on Human Mast Cell Fc Epsilon Receptor I Expression and Mediator Release. J. Immunol. 1999, 162, 5455–5465. [Google Scholar]
- Goulding, L.V.; Yang, J.; Jiang, Z.; Zhang, H.; Lea, D.; Emes, R.D.; Dottorini, T.; Pu, J.; Liu, J.; Chang, K.C. Thapsigargin at Non-Cytotoxic Levels Induces a Potent Host Antiviral Response that Blocks Influenza A Virus Replication. Viruses 2020, 12, 1093. [Google Scholar] [CrossRef]
- Zhao, H.; Raines, L.N.; Huang, S.C. Molecular Chaperones: Molecular Assembly Line Brings Metabolism and Immunity in Shape. Metabolites 2020, 10, 394. [Google Scholar] [CrossRef]
- Taborska, P.; Bartunkova, J.; Smrz, D. Simultaneous in vitro Generation of Human CD34+-derived Dendritic Cells and Mast Cells from Non-mobilized Peripheral Blood Mononuclear Cells. J. Immunol. Methods 2018, 458, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Tkaczyk, C.; Beaven, M.A.; Brachman, S.M.; Metcalfe, D.D.; Gilfillan, A.M. The Phospholipase Cg1-dependent Pathway of FceRI-mediated Mast Cell Activation is Regulated Independently of Phosphatidylinositol 3-kinase. J. Biol. Chem. 2003, 278, 48474–48484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taborska, P.; Stakheev, D.; Svobodova, H.; Strizova, Z.; Bartunkova, J.; Smrz, D. Acute Conditioning of Antigen-Expanded CD8+ T Cells via the GSK3beta-mTORC Axis Differentially Dictates Their Immediate and Distal Responses after Antigen Rechallenge. Cancers 2020, 12, 3766. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taborska, P.; Stakheev, D.; Bartunkova, J.; Smrz, D. Thapsigargin-Stimulated LAD2 Human Mast Cell Line Is a Potent Cellular Adjuvant for the Maturation of Monocyte-Derived Dendritic Cells for Adoptive Cellular Immunotherapy. Int. J. Mol. Sci. 2021, 22, 3978. https://doi.org/10.3390/ijms22083978
Taborska P, Stakheev D, Bartunkova J, Smrz D. Thapsigargin-Stimulated LAD2 Human Mast Cell Line Is a Potent Cellular Adjuvant for the Maturation of Monocyte-Derived Dendritic Cells for Adoptive Cellular Immunotherapy. International Journal of Molecular Sciences. 2021; 22(8):3978. https://doi.org/10.3390/ijms22083978
Chicago/Turabian StyleTaborska, Pavla, Dmitry Stakheev, Jirina Bartunkova, and Daniel Smrz. 2021. "Thapsigargin-Stimulated LAD2 Human Mast Cell Line Is a Potent Cellular Adjuvant for the Maturation of Monocyte-Derived Dendritic Cells for Adoptive Cellular Immunotherapy" International Journal of Molecular Sciences 22, no. 8: 3978. https://doi.org/10.3390/ijms22083978
APA StyleTaborska, P., Stakheev, D., Bartunkova, J., & Smrz, D. (2021). Thapsigargin-Stimulated LAD2 Human Mast Cell Line Is a Potent Cellular Adjuvant for the Maturation of Monocyte-Derived Dendritic Cells for Adoptive Cellular Immunotherapy. International Journal of Molecular Sciences, 22(8), 3978. https://doi.org/10.3390/ijms22083978