
Emerging Role of Phospholipase-Derived Cleavage Products in Regulating Eosinophil Activity: Focus on Lysophospholipids, Polyunsaturated Fatty Acids and Eicosanoids




Abstract
:1. Introduction
2. Phospholipases Play a Key Role in Allergies
2.1. Phospholipase A2 (PLA2) and Eosinophils
2.1.1. Secreted Phospholipases A2
2.1.2. Cytosolic Phospholipases A2
2.1.3. Platelet Activating Factor Acetylhydrolase (PAF-AH)
2.1.4. Phospholipase A2 Cleavage Products: Lysophospholipids
2.1.5. Phospholipase A2 Cleavage Products: Free Fatty Acids
3. Eicosanoids and Eosinophils
3.1. Specialized Pro-Resolving Mediators: Resolvins, Protectins, and Lipoxins
3.2. Leukotrienes
3.3. Prostaglandins
3.3.1. Prostaglandin D2
3.3.2. Prostaglandin E2
3.3.3. Prostaglandin I2-Prostacyclin
3.4. 5-Oxo-8,11,14-Eicosatrienoic Acid (5-Oxo-ETE)
4. Phospholipase C (PLC) Cleavage Products: Diacylglycerol (DAG) and Inositol 1,4,5-Trisphosphate (IP3) and Endocannabinoids
5. Phospholipase D Cleavage Product: Lysophosphatidic Acid (LPA)
6. Non-Enzymatic Lipid Peroxidation
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wen, T.; Rothenberg, M.E.; Cells, R.E. The Regulatory Function of Eosinophils. Microbiol. Spectr. 2016, 4, 1–12. [Google Scholar]
- Lee, J.J.; Jacobsen, E.A.; McGarry, M.P.; Schleimer, R.P.; Lee, N.A. Eosinophils in Health and Disease: The LIAR Hypothesis. Clin. Exp. Allergy 2010, 40, 563–575. [Google Scholar] [CrossRef] [Green Version]
- Rothenberg, M.E.; Hogan, S.P. The Eosinophil. Annu. Rev. Immunol. 2006, 24, 147–174. [Google Scholar] [CrossRef] [PubMed]
- Fulkerson, P.C.; Rothenberg, M.E. Targeting Eosinophils in Allergy, Inflammation and Beyond. Nat. Rev. Drug Discov. 2013, 12, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kariyawasam, H.H.; Robinson, D.S. The Role of Eosinophils in Airway Tissue Remodelling in Asthma. Curr. Opin. Immunol. 2007, 19, 681–686. [Google Scholar] [CrossRef]
- Acharya, K.R.; Ackerman, S.J. Eosinophil Granule Proteins: Form and Function. J. Biol. Chem. 2014, 289, 17406–17415. [Google Scholar] [CrossRef] [Green Version]
- Legrand, F.; Klion, A.D. Biologic Therapies Targeting Eosinophils: Current Status and Future Prospects. J. Allergy Clin. Immunol. Pract. 2015, 3, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busse, W.W.; Bleecker, E.R.; FitzGerald, J.M.; Ferguson, G.T.; Barker, P.; Sproule, S.; Olsson, R.F.; Martin, U.J.; Goldman, M.; Yañez, A.; et al. Long-Term Safety and Efficacy of Benralizumab in Patients with Severe, Uncontrolled Asthma: 1-Year Results from the BORA Phase 3 Extension Trial. Lancet Respir. Med. 2019, 7, 46–59. [Google Scholar] [CrossRef]
- Dellon, E.S.; Peterson, K.A.; Murray, J.A.; Falk, G.W.; Gonsalves, N.; Chehade, M.; Genta, R.M.; Leung, J.; Khoury, P.; Klion, A.D.; et al. Anti–Siglec-8 Antibody for Eosinophilic Gastritis and Duodenitis. N. Engl. J. Med. 2020, 383, 1624–1634. [Google Scholar] [CrossRef] [PubMed]
- Reichman, H.; Itan, M.; Rozenberg, P.; Yarmolovski, T.; Brazowski, E.; Varol, C.; Gluck, N.; Shapira, S.; Arber, N.; Qimron, U.; et al. Activated Eosinophils Exert Antitumorigenic Activities in Colorectal Cancer. Cancer Immunol. Res. 2019, 7, 388–400. [Google Scholar] [CrossRef] [Green Version]
- Kienzl, M.; Hasenoehrl, C.; Valadez-Cosmes, P.; Maitz, K.; Sarsembayeva, A.; Sturm, E.; Heinemann, A.; Kargl, J.; Schicho, R. IL-33 Reduces Tumor Growth in Models of Colorectal Cancer with the Help of Eosinophils. Oncoimmunology 2020, 9, 1776059. [Google Scholar] [CrossRef]
- Munitz, A.; Hogan, S.P. Alarming Eosinophils to Combat Tumors. Nat. Immunol. 2019, 20, 250–252. [Google Scholar] [CrossRef]
- Grisaru-Tal, S.; Itan, M.; Grass, D.G.; Torres-Roca, J.; Eschrich, S.A.; Gordon, Y.; Dolitzky, A.; Hazut, I.; Avlas, S.; Jacobsen, E.A.; et al. Primary Tumors from Mucosal Barrier Organs Drive Unique Eosinophil Infiltration Patterns and Clinical Associations. Oncoimmunology 2021, 10, 1859732. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Y.; Wang, M.; Wang, S.; Jeong, J.-M.; Xu, L.; Wen, Y.; Emontzpohl, C.; Atkins, C.L.; Duong, K.; et al. Eosinophils Attenuate Hepatic Ischemia-Reperfusion Injury in Mice through ST2-Dependent IL-13 Production. Sci. Transl. Med. 2021, 13, eabb6576. [Google Scholar] [CrossRef]
- Liu, J.; Yang, C.; Liu, T.; Deng, Z.; Fang, W.; Zhang, X.; Li, J.; Huang, Q.; Liu, C.; Wang, Y.; et al. Eosinophils Improve Cardiac Function after Myocardial Infarction. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Ding, R.; Yang, Z.; Huang, D.; Wang, Y.; Li, X.; Zhou, X.; Yan, L.; Lu, W.; Zhang, Z. Identification of Parameters in Routine Blood and Coagulation Tests Related to the Severity of COVID-19. Int. J. Med. Sci. 2021, 18, 1207–1215. [Google Scholar] [CrossRef]
- Nair, A.P.; Soliman, A.; Al Masalamani, M.A.; De Sanctis, V.; Nashwan, A.J.; Sasi, S.; Ali, E.A.; Hassan, O.A.; Iqbal, F.M.; Yassin, M.A. Clinical Outcome of Eosinophilia in Patients with Covid-19: A Controlled Study. Acta Biomed. 2020, 91, 1–10. [Google Scholar]
- Cazzaniga, M.; Fumagalli, L.; D’Angelo, L.; Cerino, M.; Bonfanti, G.; Fumagalli, R.; Schiavo, G.; Lorini, C.; Lainu, E.; Terragni, S.; et al. Eosinopenia Is a Reliable Marker of Severe Disease and Unfavorable Outcome in Patients with COVID-19 Pneumonia. Int. J. Clin. Pract. 2021, e14047. [Google Scholar] [CrossRef]
- Outh, R.; Boutin, C.; Gueudet, P.; Suzuki, M.; Saada, M.; Aumaître, H. Eosinopenia <100/ΜL as a Marker of Active COVID-19: An Observational Prospective Study. J. Microbiol. Immunol. Infect. 2021, 54, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Georgakopoulou, V.E.; Garmpis, N.; Damaskos, C.; Valsami, S.; Dimitroulis, D.; Diamantis, E.; Farmaki, P.; Papageorgiou, C.V.; Makrodimitri, S.; Gravvanis, N.; et al. The Impact of Peripheral Eosinophil Counts and Eosinophil to Lymphocyte Ratio (ELR) in the Clinical Course of COVID-19 Patients: A Retrospective Study. In Vivo 2021, 35, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Ferastraoaru, D.; Hudes, G.; Jerschow, E.; Jariwala, S.; Karagic, M.; de Vos, G.; Rosenstreich, D.; Ramesh, M. Eosinophilia in Asthma Patients Is Protective Against Severe COVID-19 Illness. J. Allergy Clin. Immunol. Pract. 2021, 9, 1152–1162. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Yousefi, S.; Germic, N.; Arnold, I.C.; Haczku, A.; Karaulov, A.V.; Simon, D.; Rosenberg, H.F. The Cellular Functions of Eosinophils: Collegium Internationale Allergologicum (CIA) Update 2020. Int. Arch. Allergy Immunol. 2020, 181, 11–23. [Google Scholar] [CrossRef]
- Sokolowska, M.; Stefanska, J.; Wodz-Naskiewicz, K.; Cieslak, M.; Pawliczak, R. Cytosolic Phospholipase A2 Group IVA Is Overexpressed in Patients with Persistent Asthma and Regulated by the Promoter Microsatellites. J. Allergy Clin. Immunol. 2010, 125, 1393–1395. [Google Scholar] [CrossRef]
- Bowton, D.L.; Seeds, M.C.; Fasano, M.B.; Goldsmith, B.; Bass, D.A. Phospholipase A2 and Arachidonate Increase in Bronchoalveolar Lavage Fluid after Inhaled Antigen Challenge in Asthmatics. Am. J. Respir. Crit. Care Med. 1997, 155, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Blom, M.; Tool, A.T.J.; Wever, P.C.; Wolbink, G.J.; Brouwer, M.C.; Calafat, J.; Egesten, A.; Knol, E.F.; Hack, C.E.; Roos, D.; et al. Human Eosinophils Express, Relative to Other Circulating Leukocytes, Large Amounts of Secretory 14-KD Phospholipase A2. Blood 1998, 91, 3037–3043. [Google Scholar]
- Miyata, J.; Fukunaga, K.; Kawashima, Y.; Watanabe, T.; Saitoh, A.; Hirosaki, T.; Araki, Y.; Kikawada, T.; Betsuyaku, T.; Ohara, O.; et al. Dysregulated Fatty Acid Metabolism in Nasal Polyp-Derived Eosinophils from Patients with Chronic Rhinosinusitis. Allergy Eur. J. Allergy Clin. Immunol. 2019, 74, 1113–1124. [Google Scholar] [CrossRef]
- Rodriguez-Perez, N.; Schiavi, E.; Frei, R.; Ferstl, R.; Wawrzyniak, P.; Smolinska, S.; Sokolowska, M.; Sievi, N.A.; Kohler, M.; Schmid-Grendelmeier, P.; et al. Altered Fatty Acid Metabolism and Reduced Stearoyl-Coenzyme a Desaturase Activity in Asthma. Allergy Eur. J. Allergy Clin. Immunol. 2017, 72, 1744–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, J.; Fukunaga, K.; Kawashima, Y.; Ohara, O.; Kawana, A.; Asano, K.; Arita, M. Dysregulated Metabolism of Polyunsaturated Fatty Acids in Eosinophilic Allergic Diseases. Prostaglandins Other Lipid Mediat. 2020, 150, 106477. [Google Scholar] [CrossRef]
- Filkin, S.Y.; Lipkin, A.V.; Fedorov, A.N. Phospholipase Superfamily: Structure, Functions, and Biotechnological Applications. Biochemistry 2020, 85, 177–195. [Google Scholar] [CrossRef]
- Aloulou, A.; Rahier, R.; Arhab, Y.; Noiriel, A.; Abousalham, A. Phospholipases: An overview. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2018; Volume 1835, pp. 69–105. [Google Scholar]
- Pniewska, E.; Pawliczak, R. The Involvement of Phospholipases A2 in Asthma and Chronic Obstructive Pulmonary Disease. Mediat. Inflamm. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Burke, J.E.; Dennis, E.A. Phospholipase A 2 Structure/Function, Mechanism, and Signaling. J. Lipid Res. 2009, 50, S237. [Google Scholar] [CrossRef] [Green Version]
- Dennis, E.A. Introduction to Thematic Review Series: Phospholipases: Central Role in Lipid Signaling and Disease. J. Lipid Res. 2015, 56, 1245–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.R.; Strek, M.E.; Kulp, G.V.P.; Spaethe, S.M.; Burch, R.A.; Neeley, S.P.; Leff, A.R. Regulation of Human Eosinophil Degranulation and Activation by Endogenous Phospholipase A2. J. Clin. Investig. 1993, 91, 2118–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triggiani, M.; Giannattasio, G.; Calabrese, C.; Loffredo, S.; Granata, F.; Fiorello, A.; Santini, M.; Gelb, M.H.; Marone, G. Lung Mast Cells Are a Source of Secreted Phospholipases A2. J. Allergy Clin. Immunol. 2009, 124, 558–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kertys, M.; Grendar, M.; Kosutova, P.; Mokra, D.; Mokry, J. Plasma Based Targeted Metabolomic Analysis Reveals Alterations of Phosphatidylcholines and Oxidative Stress Markers in Guinea Pig Model of Allergic Asthma. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165572. [Google Scholar] [CrossRef]
- Nolin, J.D.; Lai, Y.; Ogden, H.L.; Manicone, A.M.; Murphy, R.C.; An, D.; Frevert, C.W.; Ghomashchi, F.; Naika, G.S.; Gelb, M.H.; et al. Secreted PLA2 Group X Orchestrates Innate and Adaptive Immune Responses to Inhaled Allergen. JCI Insight 2017, 2, e94929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogden, H.L.; Lai, Y.; Nolin, J.D.; An, D.; Frevert, C.W.; Gelb, M.H.; Altemeier, W.A.; Hallstrand, T.S. Secreted Phospholipase A 2 Group X Acts as an Adjuvant for Type 2 Inflammation, Leading to an Allergen-Specific Immune Response in the Lung. J. Immunol. 2020, 204, 3097–3107. [Google Scholar] [CrossRef]
- Henderson, W.R.; Chi, E.Y.; Bollinger, J.G.; Tien, Y.T.; Ye, X.; Castelli, L.; Rubtsov, Y.P.; Singer, A.G.; Chiang, G.K.S.; Nevalainen, T.; et al. Importance of Group X-Secreted Phospholipase A2 in Allergen-Induced Airway Inflammation and Remodeling in a Mouse Asthma Model. J. Exp. Med. 2007, 204, 865–877. [Google Scholar] [CrossRef]
- Hallstrand, T.S.; Lai, Y.; Hooper, K.A.; Oslund, R.C.; Altemeier, W.A.; Matute-Bello, G.; Gelb, M.H. Endogenous Secreted Phospholipase A2 Group X Regulates Cysteinyl Leukotrienes Synthesis by Human Eosinophils. J. Allergy Clin. Immunol. 2016, 137, 268–277.e8. [Google Scholar] [CrossRef]
- Lai, Y.; Oslund, R.C.; Bollinger, J.G.; Henderson, W.R.; Santana, L.F.; Altemeier, W.A.; Gelb, M.H.; Hallstrand, T.S. Eosinophil Cysteinyl Leukotriene Synthesis Mediated by Exogenous Secreted Phospholipase A2 Group X. J. Biol. Chem. 2010, 285, 41491–41500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, W.R.; Oslund, R.C.; Bollinger, J.G.; Ye, X.; Tien, Y.T.; Xue, J.; Gelb, M.H. Blockade of Human Group X Secreted Phospholipase A 2 (GX-SPLA 2)-Induced Airway Inflammation and Hyperresponsiveness in a Mouse Asthma Model by a Selective GX-SPLA 2 Inhibitor. J. Biol. Chem. 2011, 286, 28049–28055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, N.M.; Meliton, A.Y.; Arm, J.P.; Bonventre, J.V.; Cho, W.; Leff, A.R. Deletion of Secretory Group V Phospholipase A2 Attenuates Cell Migration and Airway Hyperresponsiveness in Immunosensitized Mice. J. Immunol. 2007, 179, 4800–4807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannattasio, G.; Fujioka, D.; Xing, W.; Katz, H.R.; Boyce, J.A.; Balestrieri, B. Group V Secretory Phospholipase A2 Reveals Its Role in House Dust Mite-Induced Allergic Pulmonary Inflammation by Regulation of Dendritic Cell Function. J. Immunol. 2010, 185, 4430–4438. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, M.; Samuchiwal, S.K.; Quehenberger, O.; Boyce, J.A.; Balestrieri, B. Macrophages Regulate Lung ILC2 Activation via Pla2g5-Dependent Mechanisms. Mucosal Immunol. 2018, 11, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, N.M.; Kim, Y.J.; Meliton, A.Y.; Kim, K.P.; Han, S.K.; Boetticher, E.; O’Leary, E.; Myou, S.; Zhu, X.; Bonventre, J.V.; et al. Human Group V Phospholipase A2 Induces Group IVA Phospholipase A2-Independent Cysteinyl Leukotriene Synthesis in Human Eosinophils. J. Biol. Chem. 2003, 278, 38813–38820. [Google Scholar] [CrossRef] [Green Version]
- Myou, S.; Sano, H.; Fujimura, M.; Zhu, X.; Kurashima, K.; Kita, T.; Nakao, S.; Nonomura, A.; Shioya, T.; Kim, K.P.; et al. Blockade of Eosinophil Migration and Airway Hyperresponsiveness by CPLA2-Inhibition. Nat. Immunol. 2001, 2, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Yanes, D.A.; Mosser-Goldfarb, J.L. Emerging Therapies for Atopic Dermatitis: The Prostaglandin/Leukotriene Pathway. J. Am. Acad. Dermatol. 2018, 78, S71–S75. [Google Scholar] [CrossRef]
- Kokotou, M.G.; Galiatsatou, G.; Magrioti, V.; Koutoulogenis, G.; Barbayianni, E.; Limnios, D.; Mouchlis, V.D.; Satpathy, B.; Navratil, A.; Dennis, E.A.; et al. 2-Oxoesters: A Novel Class of Potent and Selective Inhibitors of Cytosolic Group IVA Phospholipase A2. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef]
- Koutoulogenis, G.S.; Kokotou, M.G.; Hayashi, D.; Mouchlis, V.D.; Dennis, E.A.; Kokotos, G. 2-Oxoester Phospholipase A2 Inhibitors with Enhanced Metabolic Stability. Biomolecules 2020, 10, 491. [Google Scholar] [CrossRef] [Green Version]
- Henderson, W.R.; Lu, J.; Poole, K.M.; Dietsch, G.N.; Chi, E.Y. Recombinant Human Platelet-Activating Factor-Acetylhydrolase Inhibits Airway Inflammation and Hyperreactivity in Mouse Asthma Model. J. Immunol. 2000, 164, 3360–3367. [Google Scholar] [CrossRef] [Green Version]
- Henig, N.R.; Aitken, M.L.; Liu, M.C.; Yu, A.S.; Henderson, W.R. Effect of Recombinant Human Platelet-Activating Factor-Acetylhydrolase on Allergen-Induced Asthmatic Responses. Am. J. Respir. Crit. Care Med. 2000, 162, 523–527. [Google Scholar] [CrossRef]
- Maiti, R.; Rahman, J.; Jaida, J.; Allala, U.; Palani, A. Rupatadine and Levocetirizine for Seasonal Allergic Rhinitis: A Comparative Study of Efficacy and Safety. Arch. Otolaryngol. Head Neck Surg. 2010, 136, 796–800. [Google Scholar] [CrossRef] [Green Version]
- Metz, M.; Maurer, M. Rupatadine for the Treatment of Allergic Rhinitis and Urticaria. Expert Rev. Clin. Immunol. 2011, 7, 15–20. [Google Scholar] [CrossRef]
- Knuplez, E.; Kienzl, M.; Trakaki, A.; Schicho, R.; Heinemann, A.; Sturm, E.M.; Marsche, G. The Anti-parasitic Drug Miltefosine Suppresses Human Eosinophil Activation and Ameliorates Murine Allergic Inflammation In Vivo. Br. J. Pharmacol. 2021, 178, 1234–1248. [Google Scholar] [CrossRef] [PubMed]
- Magerl, M.; Rother, M.; Bieber, T.; Biedermann, T.; Brasch, J.; Dominicus, R.; Hunzelmann, N.; Jakob, T.; Mahler, V.; Popp, G.; et al. Randomized, Double-Blind, Placebo-Controlled Study of Safety and Efficacy of Miltefosine in Antihistamine-Resistant Chronic Spontaneous Urticaria. J. Eur. Acad. Dermatol. Venereol. 2013, 27, e363–e369. [Google Scholar] [CrossRef]
- Sato, M.; Aoki-Saito, H.; Fukuda, H.; Ikeda, H.; Koga, Y.; Yatomi, M.; Tsurumaki, H.; Maeno, T.; Saito, T.; Nakakura, T.; et al. Resolvin E3 Attenuates Allergic Airway Inflammation via the Interleukin-23-Interleukin-17A Pathway. FASEB J. 2019, 33, 12750–12759. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, H.; Ikeda, H.; Muromoto, R.; Hirashima, K.; Ishimura, K.; Fujiwara, K.; Aoki-Saito, H.; Hisada, T.; Watanabe, M.; Ishihara, J.; et al. Synthesis of Resolvin E3, a Proresolving Lipid Mediator, and Its Deoxy Derivatives: Identification of 18-Deoxy-Resolvin E3 as a Potent Anti-Inflammatory Agent. J. Org. Chem. 2020, 85, 14190–14200. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Xie, Y.K.; Zhang, Z.J.; Wang, Z.; Xu, Z.Z.; Ji, R.R. GPR37 Regulates Macrophage Phagocytosis and Resolution of Inflammatory Pain. J. Clin. Investig. 2018, 128, 3568–3582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, B.D.; Kohli, P.; Gotlinger, K.; Haworth, O.; Hong, S.; Kazani, S.; Israel, E.; Haley, K.J.; Serhan, C.N. Protectin D1 Is Generated in Asthma and Dampens Airway Inflammation and Hyperresponsiveness. J. Immunol. 2007, 178, 496–502. [Google Scholar] [CrossRef] [Green Version]
- Levy, B.D.; De Sanctis, G.T.; Devchand, P.R.; Kim, E.; Ackerman, K.; Schmidt, B.A.; Szczeklik, W.; Drazen, J.M.; Serhan, C.N. Multi-Pronged Inhibition of Airway Hyper-Responsiveness and Inflammation by Lipoxin A4. Nat. Med. 2002, 8, 1018–1023. [Google Scholar] [CrossRef]
- Kong, X.; Wu, S.H.; Zhang, L.; Chen, X.Q. Pilot Application of Lipoxin A4 Analog and Lipoxin A4 Receptor Agonist in Asthmatic Children with Acute Episodes. Exp. Ther. Med. 2017, 14, 2284–2290. [Google Scholar] [CrossRef]
- Wu, S.H.; Chen, X.Q.; Liu, B.; Wu, H.J.; Dong, L. Efficacy and Safety of 15(R/S)-Methyl-Lipoxin A4 in Topical Treatment of Infantile Eczema. Br. J. Dermatol. 2013, 168, 172–178. [Google Scholar] [CrossRef]
- Haruna, T.; Soga, M.; Morioka, Y.; Imura, K.; Furue, Y.; Yamamoto, M.; Hayakawa, J.; Deguchi, M.; Arimura, A.; Yasui, K. The Inhibitory Effect of S-777469, a Cannabinoid Type 2 Receptor Agonist, on Skin Inflammation in Mice. Pharmacology 2017, 99, 259–267. [Google Scholar] [CrossRef]
- Abohalaka, R.; Bozkurt, T.E.; Nemutlu, E.; Onder, S.C.; Sahin-Erdemli, I. The Effects of Fatty Acid Amide Hydrolase and Monoacylglycerol Lipase Inhibitor Treatments on Lipopolysaccharide-Induced Airway Inflammation in Mice. Pulm. Pharmacol. Ther. 2020, 62, 101920. [Google Scholar] [CrossRef] [PubMed]
- Larose, M.C.; Turcotte, C.; Chouinard, F.; Ferland, C.; Martin, C.; Provost, V.; Laviolette, M.; Flamand, N. Mechanisms of Human Eosinophil Migration Induced by the Combination of IL-5 and the Endocannabinoid 2-Arachidonoyl-Glycerol. J. Allergy Clin. Immunol. 2014, 133, 1480–1482.e3. [Google Scholar] [CrossRef]
- Park, G.Y.; Lee, Y.G.; Berdyshev, E.; Nyenhuis, S.; Du, J.; Fu, P.; Gorshkova, I.A.; Li, Y.; Chung, S.; Karpurapu, M.; et al. Autotaxin Production of Lysophosphatidic Acid Mediates Allergic Asthmatic Inflammation. Am. J. Respir. Crit. Care Med. 2013, 188, 928–940. [Google Scholar] [CrossRef] [PubMed]
- Jendzjowsky, N.G.; Roy, A.; Barioni, N.O.; Kelly, M.M.; Green, F.H.Y.; Wyatt, C.N.; Pye, R.L.; Tenorio-Lopes, L.; Wilson, R.J.A. Preventing Acute Asthmatic Symptoms by Targeting a Neuronal Mechanism Involving Carotid Body Lysophosphatidic Acid Receptors. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, T.N.; Oczypok, E.A.; Dutz, R.E.; Donnell, M.L.; Myerburg, M.M.; Oury, T.D. The Receptor for Advanced Glycation End Products Is a Critical Mediator of Type 2 Cytokine Signaling in the Lungs. J. Allergy Clin. Immunol. 2019, 144, 796–808.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchette, D.; Preuss, C. V Zileuton; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Choi, J.; Azmat, C.E. Leukotriene Receptor Antagonists; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Aritake, K.; Kado, Y.; Inoue, T.; Miyano, M.; Urade, Y. Structural and Functional Characterization of HQL-79, an Orally Selective Inhibitor of Human Hematopoietic Prostaglandin D Synthase. J. Biol. Chem. 2006, 281, 15277–15286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoyagi, H.; Kajiwara, D.; Tsunekuni, K.; Tanaka, K.; Miyoshi, K.; Hirasawa, N. Potential Synergistic Effects of Novel Hematopoietic Prostaglandin D Synthase Inhibitor TAS-205 and Different Types of Anti-Allergic Medicine on Nasal Obstruction in a Guinea Pig Model of Experimental Allergic Rhinitis. Eur. J. Pharmacol. 2020, 875, 173030. [Google Scholar] [CrossRef] [PubMed]
- Brightling, C.E.; Gaga, M.; Inoue, H.; Li, J.; Maspero, J.; Wenzel, S.; Maitra, S.; Lawrence, D.; Brockhaus, F.; Lehmann, T.; et al. Effectiveness of Fevipiprant in Reducing Exacerbations in Patients with Severe Asthma (LUSTER-1 and LUSTER-2): Two Phase 3 Randomised Controlled Trials. Lancet Respir. Med. 2021, 9, 43–56. [Google Scholar] [CrossRef]
- Okubo, K.; Hashiguchi, K.; Takeda, T.; Baba, K.; Kitagoh, H.; Miho, H.; Tomomatsu, H.; Yamaguchi, S.; Odani, M.; Yamamotoya, H. A Randomized Controlled Phase II Clinical Trial Comparing ONO-4053, a Novel DP1 Antagonist, with a Leukotriene Receptor Antagonist Pranlukast in Patients with Seasonal Allergic Rhinitis. Allergy Eur. J. Allergy Clin. Immunol. 2017, 72, 1565–1575. [Google Scholar] [CrossRef]
- Sturm, E.M.; Schratl, P.; Schuligoi, R.; Konya, V.; Sturm, G.J.; Lippe, I.T.; Peskar, B.A.; Heinemann, A. Prostaglandin E2 Inhibits Eosinophil Trafficking through E-Prostanoid 2 Receptors. J. Immunol. 2008, 181, 7273–7283. [Google Scholar] [CrossRef] [Green Version]
- Serra-Pages, M.; Torres, R.; Plaza, J.; Herrerias, A.; Costa-Farré, C.; Marco, A.; Jiménez, M.; Maurer, M.; Picado, C.; de Mora, F. Activation of the Prostaglandin E2 Receptor EP2 Prevents House Dust Mite-Induced Airway Hyperresponsiveness and Inflammation by Restraining Mast Cells’ Activity. Clin. Exp. Allergy 2015, 45, 1590–1600. [Google Scholar] [CrossRef]
- Idzko, M.; Hammad, H.; Van Nimwegen, M.; Kool, M.; Vos, N.; Hoogsteden, H.C.; Lambrecht, B.N. Inhaled Iloprost Suppresses the Cardinal Features of Asthma via Inhibition of Airway Dendritic Cell Function. J. Clin. Investig. 2007, 117, 464–472. [Google Scholar] [CrossRef]
- Sturm, E.M.; Schuligoi, R.; Konya, V.; Sturm, G.J.; Heinemann, A. Inhibitory Effect of Prostaglandin I2 on Bone Marrow Kinetics of Eosinophils in the Guinea Pig. J. Leukoc. Biol. 2011, 90, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.A.; Cossette, C.; Chourey, S.; Ye, Q.; Reddy, C.N.; Rokach, J.; Powell, W.S. Inhibition of Allergen-Induced Dermal Eosinophilia by an Oxoeicosanoid Receptor Antagonist in Non-Human Primates. Br. J. Pharmacol. 2020, 177, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Adler, D.H.; Cogan, J.D.; Phillips, J.A.; Schnetz-Boutaud, N.; Milne, G.L.; Iverson, T.; Stein, J.A.; Brenner, D.A.; Morrow, J.D.; Boutaud, O.; et al. Inherited Human CPLA2α Deficiency Is Associated with Impaired Eicosanoid Biosynthesis, Small Intestinal Ulceration, and Platelet Dysfunction. J. Clin. Investig. 2008, 118, 2121–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallstrand, T.S.; Chi, E.Y.; Singer, A.G.; Gelb, M.H.; Henderson, W.R. Secreted Phospholipase A2 Group X Overexpression in Asthma and Bronchial Hyperresponsiveness. Am. J. Respir. Crit. Care Med. 2007, 176, 1072–1078. [Google Scholar] [CrossRef] [Green Version]
- Urasaki, T.; Ninomiya, H.; Takasaki, J.; Kawauchi, Y.; Nagasawa, T.; Masuho, Y. Cytosolic Phospholipase A2, Increased and Activated in the Eosinophils of Patients with Hypereosinophilic Syndrome in Vivo, Is Involved in the Augmented Release of Leukotriene C4. Inflamm. Res. 1999, 48, 36–40. [Google Scholar] [CrossRef]
- Clark, J.D.; Williams, C.M.M. Utility of cytosolic phospholipase A2α (cPLA2α) inhibitors in the treatment of asthma. In New Drugs and Targets for Asthma and COPD; S. Karger AG: Basel, Switzerland, 2010; Volume 39, pp. 207–212. ISBN 9783805595674. [Google Scholar]
- Nikolaou, A.; Kokotou, M.G.; Vasilakaki, S.; Kokotos, G. Small-Molecule Inhibitors as Potential Therapeutics and as Tools to Understand the Role of Phospholipases A2. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Hewson, C.A.; Patel, S.; Calzetta, L.; Campwala, H.; Havard, S.; Luscombe, E.; Clarke, P.A.; Peachell, P.T.; Matera, M.G.; Cazzola, M.; et al. Preclinical Evaluation of an Inhibitor of Cytosolic Phospholipase A 2α for the Treatment of Asthma. J. Pharmacol. Exp. Ther. 2012, 340, 656–665. [Google Scholar] [CrossRef] [Green Version]
- Karasawa, K.; Harada, A.; Satoh, N.; Inoue, K.; Setaka, M. Plasma Platelet Activating Factor-Acetylhydrolase (PAF-AH). Prog. Lipid Res. 2003, 42, 93–114. [Google Scholar] [CrossRef]
- Snyder, F. Platelet-Activating Factor: The Biosynthetic and Catabolic Enzymes. Biochem. J. 1995, 305, 689–705. [Google Scholar] [CrossRef] [Green Version]
- Gill, P.; Jindal, N.L.; Jagdis, A.; Vadas, P. Platelets in the Immune Response: Revisiting Platelet-Activating Factor in Anaphylaxis. J. Allergy Clin. Immunol. 2015, 135, 1424–1432. [Google Scholar] [CrossRef] [Green Version]
- Dyer, K.D.; Percopo, C.M.; Xie, Z.; Yang, Z.; Kim, J.D.; Davoine, F.; Lacy, P.; Druey, K.M.; Moqbel, R.; Rosenberg, H.F. Mouse and Human Eosinophils Degranulate in Response to Platelet-Activating Factor (PAF) and LysoPAF via a PAF-Receptor–Independent Mechanism: Evidence for a Novel Receptor. J. Immunol. 2010, 184, 6327–6334. [Google Scholar] [CrossRef]
- Kroegel, C.; Matthys, H. Platelet-Activating Factor-Induced Human Eosinophil Activation. Generation and Release of Cyclo-Oxygenase Metabolites in Human Blood Eosinophils from Asthmatics. Immunology 1993, 78, 279–285. [Google Scholar] [PubMed]
- Bankers-Fulbright, J.L.; Kephart, G.M.; Bartemes, K.R.; Kita, H.; O’Grady, S.M. Platelet-Activating Factor Stimulates Cytoplasmic Alkalinization and Granule Acidification in Human Eosinophils. J. Cell Sci. 2004, 117, 5749–5757. [Google Scholar] [CrossRef] [Green Version]
- Wardlaw, A.J.; Moqbel, R.; Cromwell, O.; Kay, A.B. Platelet-Activating Factor. A Potent Chemotactic and Chemokinetic Factor for Human Eosinophils. J. Clin. Investig. 1986, 78, 1701–1706. [Google Scholar] [CrossRef] [PubMed]
- Mullol, J. Positioning of Antihistamines in the Allergic Rhinitis and Its Impact on Asthma (ARIA) Guidelines. Clin. Exp. Allergy Rev. 2012, 12, 17–26. [Google Scholar] [CrossRef]
- Mullol, J.; Izquierdo, I.; Okubo, K.; Canonica, G.W.; Bousquet, J.; Valero, A. Clinically Relevant Effect of Rupatadine 20 Mg and 10 Mg in Seasonal Allergic Rhinitis: A Pooled Responder Analysis. Clin. Transl. Allergy 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Santamaría, E.; Izquierdo, I.; Valle, M.; Vermeulen, J.; Potter, P. Rupatadine Oral Solution for 2–5-Year-Old Children with Allergic Rhinitis: A Safety, Open-Label, Prospective Study. J. Asthma Allergy 2018, 11, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, P.; Maspero, J.F.; Vermeulen, J.; Barkai, L.; Németh, I.; Baillieau, R.A.; Garde, J.M.; Giralt, J.; Doménech, A.; Izquierdo, I.; et al. Rupatadine Oral Solution in Children with Persistent Allergic Rhinitis: A Randomized, Double-Blind, Placebo-Controlled Study. Pediatr. Allergy Immunol. 2013, 24, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Nieto, A.; Nieto, M.; Mazón, Á. The Clinical Evidence of Second-Generation H1-Antihistamines in the Treatment of Allergic Rhinitis and Urticaria in Children over 2 Years with a Special Focus on Rupatadine. Expert Opin. Pharmacother. 2020, 22, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Roberts, G.; Xatzipsalti, M.; Borrego, L.M.; Custovic, A.; Halken, S.; Hellings, P.W.; Papadopoulos, N.G.; Rotiroti, G.; Scadding, G.; Timmermans, F.; et al. Paediatric Rhinitis: Position Paper of the European Academy of Allergy and Clinical Immunology. Allergy 2013, 68, 1102–1116. [Google Scholar] [CrossRef] [PubMed]
- Church, M.K.; Maurer, M.; Simons, F.E.R.; Bindslev-Jensen, C.; Van Cauwenberge, P.; Bousquet, J.; Holgate, S.T.; Zuberbier, T. Risk of First-Generation H1-Antihistamines: A GA2LEN Position Paper. Allergy Eur. J. Allergy Clin. Immunol. 2010, 65, 459–466. [Google Scholar] [CrossRef]
- Knuplez, E.; Curcic, S.; Theiler, A.; Bärnthaler, T.; Trakaki, A.; Trieb, M.; Holzer, M.; Heinemann, A.; Zimmermann, R.; Sturm, E.M.; et al. Lysophosphatidylcholines Inhibit Human Eosinophil Activation and Suppress Eosinophil Migration in Vivo. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158686. [Google Scholar] [CrossRef]
- Hwang, S.M.; Kim, H.J.; Kim, S.M.; Jung, Y.; Park, S.W.; Chung, I.Y. Lysophosphatidylserine Receptor P2Y10: A G Protein-Coupled Receptor That Mediates Eosinophil Degranulation. Clin. Exp. Allergy 2018, 48, 990–999. [Google Scholar] [CrossRef] [PubMed]
- Frei, R.B.; Luschnig, P.; Parzmair, G.P.; Peinhaupt, M.; Schranz, S.; Fauland, A.; Wheelock, C.E.; Heinemann, A.; Sturm, E.M. Cannabinoid Receptor 2 Augments Eosinophil Responsiveness and Aggravates Allergen-Induced Pulmonary Inflammation in Mice. Allergy Eur. J. Allergy Clin. Immunol. 2016, 71, 944–956. [Google Scholar] [CrossRef]
- Czech, W.; Barbisch, M.; Tenscher, K.; Schöpf, E.; Schröder, J.M.; Norgauer, J. Chemotactic 5-Oxo-Eicosatetraenoic Acids Induce Oxygen Radical Production, Ca2+-Mobilization, and Actin Reorganization in Human Eosinophils via a Pertussis Toxin-Sensitive G-Protein. J. Investig. Dermatol. 1997, 108, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Schwenk, U.; Morita, E.; Engel, R.; Schröder, J.M. Identification of 5-Oxo-15-Hydroxy-6,8,11,13-Eicosatetraenoic Acid as a Novel and Potent Human Eosinophil Chemotactic Eicosanoid. J. Biol. Chem. 1992, 267, 12482–12488. [Google Scholar] [CrossRef]
- Sturm, G.J.; Schuligoi, R.; Sturm, E.M.; Royer, J.F.; Lang-Loidolt, D.; Stammberger, H.; Amann, R.; Peskar, B.A.; Heinemann, A. 5-Oxo-6,8,11,14-Eicosatetraenoic Acid Is a Potent Chemoattractant for Human Basophils. J. Allergy Clin. Immunol. 2005, 116, 1014–1019. [Google Scholar] [CrossRef]
- Muro, S.; Hamid, Q.; Olivenstein, R.; Taha, R.; Rokach, J.; Powell, W.S. 5-Oxo-6,8,11,14-Eicosatetraenoic Acid Induces the Infiltration of Granulocytes into Human Skin. J. Allergy Clin. Immunol. 2003, 112, 768–774. [Google Scholar] [CrossRef]
- Lin, L.; Chen, Z.; Tang, X.; Dai, F.; Wei, J.; Sun, G. 5-Oxo-ETE from Nasal Epithelial Cells Upregulates Eosinophil Cation Protein by Eosinophils in Nasal Polyps in Vitro. Int. Arch. Allergy Immunol. 2018, 177, 107–115. [Google Scholar] [CrossRef]
- Fregonese, L.; Silvestri, M.; Sabatini, F.; Rossi, G.A. Cysteinyl Leukotrienes Induce Human Eosinophil Locomotion and Adhesion Molecule Expression via a CysLT1 Receptor-Mediated Mechanism. Clin. Exp. Allergy 2002, 32, 745–750. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Nagata, M.; Kikuchi, I.; Sakamoto, Y. Leukotriene D4 and Eosinophil Transendothelial Migration, Superoxide Generation, and Degranulation via Β2 Integrin. Ann. Allergy Asthma Immunol. 2004, 93, 594–600. [Google Scholar] [CrossRef]
- Neves, J.S.; Radke, A.L.; Weller, P.F. Cysteinyl Leukotrienes Acting via Granule Membrane-Expressed Receptors Elicit Secretion from within Cell-Free Human Eosinophil Granules. J. Allergy Clin. Immunol. 2010, 125, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Tager, A.M.; Dufour, J.H.; Goodarzi, K.; Bercury, S.D.; Von Andrian, U.H.; Luster, A.D. BLTR Mediates Leukotriene B4-Induced Chemotaxis and Adhesion and Plays a Dominant Role in Eosinophil Accumulation in a Murine Model of Peritonitis. J. Exp. Med. 2000, 192, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Idzko, M.; Laut, M.; Panther, E.; Sorichter, S.; Durk, T.; Fluhr, J.W.; Herouy, Y.; Mockenhaupt, M.; Myrtek, D.; Elsner, P.; et al. Lysophosphatidic Acid Induces Chemotaxis, Oxygen Radical Production, CD11b Up-Regulation, Ca2+ Mobilization, and Actin Reorganization in Human Eosinophils via Pertussis Toxin-Sensitive G Proteins. J. Immunol. 2004, 172, 4480–4485. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, T.; Yamashita, M.; Ohata, H.; Momose, K. Lysophosphatidic Acid Enhances in Vivo Infiltration and Activation of Guinea Pig Eosinophils and Neutrophils via a Rho/Rho-Associated Protein Kinase-Mediated Pathway. J. Pharmacol. Sci. 2003, 91, 8–14. [Google Scholar] [CrossRef]
- Zhu, X.; Learoyd, J.; Butt, S.; Zhu, L.; Usatyuk, P.V.; Natarajan, V.; Munoz, N.M.; Leff, A.R. Regulation of Eosinophil Adhesion by Lysophosphatidylcholine via a Non-Store-Operated Ca2+ Channel. Am. J. Respir. Cell Mol. Biol. 2007, 36, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, O.; Kume, H.; Kondo, M.; Ito, Y.; Ito, M.; Yamaki, K. Role of Lysophosphatidylcholine in Eosinophil Infiltration and Resistance in Airways. Clin. Exp. Pharmacol. Physiol. 2004, 31, 179–184. [Google Scholar] [CrossRef]
- Kim, H.J.; Sim, M.S.; Lee, D.H.; Kim, C.; Choi, Y.; Park, H.; Chung, I.Y. Lysophosphatidylserine Induces Eosinophil Extracellular Trap Formation and Degranulation: Implications in Severe Asthma. Allergy 2020, 75, 3159–3170. [Google Scholar] [CrossRef]
- Gervais, F.G.; Cruz, R.P.G.; Chateauneuf, A.; Gale, S.; Sawyer, N.; Nantel, F.; Metters, K.M.; O’Neill, G.P. Selective Modulation of Chemokinesis, Degranulation, and Apoptosis in Eosinophils through the PGD2 Receptors CRTH2 and DP. J. Allergy Clin. Immunol. 2001, 108, 982–988. [Google Scholar] [CrossRef] [PubMed]
- Hirai, H.; Tanaka, K.; Yoshie, O.; Ogawa, K.; Kenmotsu, K.; Takamori, Y.; Ichimasa, M.; Sugamura, K.; Nakamura, M.; Takano, S.; et al. Prostaglandin D2 Selectively Induces Chemotaxis in T Helper Type 2 Cells, Eosinophils, and Basophils via Seven-Transmembrane Receptor CRTH2. J. Exp. Med. 2001, 193, 255–261. [Google Scholar] [CrossRef]
- Schratl, P.; Sturm, E.M.; Royer, J.F.; Sturm, G.J.; Lippe, I.T.; Peskar, B.A.; Heinemann, A. Hierarchy of Eosinophil Chemoattractants: Role of P38 Mitogen-Activated Protein Kinase. Eur. J. Immunol. 2006, 36, 2401–2409. [Google Scholar] [CrossRef]
- Heinemann, A.; Schuligoi, R.; Sabroe, I.; Hartnell, A.; Peskar, B.A. Δ12-Prostaglandin J2, a Plasma Metabolite of Prostaglandin D2, Causes Eosinophil Mobilization from the Bone Marrow and Primes Eosinophils for Chemotaxis. J. Immunol. 2003, 170, 4752–4758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peinhaupt, M.; Roula, D.; Theiler, A.; Sedej, M.; Schicho, R.; Marsche, G.; Sturm, E.M.; Sabroe, I.; Rothenberg, M.E.; Heinemann, A. DP1 Receptor Signaling Prevents the Onset of Intrinsic Apoptosis in Eosinophils and Functions as a Transcriptional Modulator. J. Leukoc. Biol. 2018, 104, 159–171. [Google Scholar] [CrossRef] [Green Version]
- Mesquita-Santos, F.P.; Bakker-Abreu, I.; Luna-Gomes, T.; Bozza, P.T.; Diaz, B.L.; Bandeira-Melo, C. Co-Operative Signalling through DP1 and DP2 Prostanoid Receptors Is Required to Enhance Leukotriene C4 Synthesis Induced by Prostaglandin D2 in Eosinophils. Br. J. Pharmacol. 2011, 162, 1674–1685. [Google Scholar] [CrossRef] [Green Version]
- Luschnig-Schratl, P.; Sturm, E.M.; Konya, V.; Philipose, S.; Marsche, G.; Fröhlich, E.; Samberger, C.; Lang-Loidolt, D.; Gattenlöhner, S.; Lippe, I.T.; et al. EP4 Receptor Stimulation Down-Regulates Human Eosinophil Function. Cell. Mol. Life Sci. 2011, 68, 3573–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, C.D.; Misso, N.L.A.; Watkins, D.N.; Thompson, P.J. PGE2 and Dibutyryl Cyclic Adenosine Monophosphate Prolong Eosinophil Survival In Vitro. J. Allergy Clin. Immunol. 1999, 104, 153–162. [Google Scholar] [CrossRef]
- Daffern, P.J.; Jagels, M.A.; Saad, J.J.; Fischer, W.; Hugli, T.E. Upper Airway Epithelial Cells Support Eosinophil Survival In Vitro through Production of GM-CSF and Prostaglandin E2: Regulation by Glucocorticoids and TNF-α. Allergy Asthma Proc. 1999, 20, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Gauvreau, G.M.; Watson, R.M.; O’Byrne, P.M. Protective Effects of Inhaled PGE2 on Allergen-Induced Airway Responses and Airway Inflammation. Am. J. Respir. Crit. Care Med. 1999, 159, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Konya, V.; Sturm, E.M.; Schratl, P.; Beubler, E.; Marsche, G.; Schuligoi, R.; Lippe, I.T.; Peskar, B.A.; Heinemann, A.; Austria, G. Endothelium-Derived Prostaglandin I 2 Controls the Migration of Eosinophils. J. Allergy Clin. Immunol. 2010, 125, 1105–1113. [Google Scholar] [CrossRef]
- Miyata, J.; Fukunaga, K.; Iwamoto, R.; Isobe, Y.; Niimi, K.; Takamiya, R.; Takihara, T.; Tomomatsu, K.; Suzuki, Y.; Oguma, T.; et al. Dysregulated Synthesis of Protectin D1 in Eosinophils from Patients with Severe Asthma. J. Allergy Clin. Immunol. 2013, 131, 353–360.e2. [Google Scholar] [CrossRef]
- Rabini, R.A.; Galassi, R.; Fumelli, P.; Dousset, N.; Solera, M.L.; Valdiguie, P.; Curatola, G.; Ferretti, G.; Taus, M.; Mazzanti, L. Reduced Na+-K+-ATPase Activity and Plasma Lysophosphatidylcholine Concentrations in Diabetic Patients. Diabetes 1994, 43, 915–919. [Google Scholar] [CrossRef]
- Mcmahon, H.T.; Boucrot, E. Membrane Curvature at a Glance. J. Cell Sci. 2015, 128, 1065–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, A.J.; Moores, G.R.; Steele, J. A Conductimetric Study of Erythrocyte Lysis by Lysolecithin and Linoleic Acid. Eur. J. Biochem. 1974, 48, 277–286. [Google Scholar] [CrossRef]
- Yoder, M.; Zhuge, Y.; Yuan, Y.; Holian, O.; Kuo, S.; van Breemen, R.; Thomas, L.L.; Lum, H. Bioactive Lysophosphatidylcholine 16:0 and 18:0 Are Elevated in Lungs of Asthmatic Subjects. Allergy. Asthma Immunol. Res. 2014, 6, 61. [Google Scholar] [CrossRef] [Green Version]
- Ried, J.S.; Baurecht, H.; Stückler, F.; Krumsiek, J.; Gieger, C.; Heinrich, J.; Kabesch, M.; Prehn, C.; Peters, A.; Rodriguez, E.; et al. Integrative Genetic and Metabolite Profiling Analysis Suggests Altered Phosphatidylcholine Metabolism in Asthma. Allergy Eur. J. Allergy Clin. Immunol. 2013, 68, 629–636. [Google Scholar] [CrossRef]
- Knuplez, E.; Marsche, G. An Updated Review of Pro-and Anti-Inflammatory Properties of Plasma Lysophosphatidylcholines in the Vascular System. Int. J. Mol. Sci. 2020, 21, 4501. [Google Scholar] [CrossRef]
- Marathe, G.K.; Silva, A.R.; de Castro Faria Neto, H.C.; Tjoelker, L.W.; Prescott, S.M.; Zimmerman, G.A.; McIntyre, T.M. Lysophosphatidylcholine and Lyso-PAF Display PAF-like Activity Derived from Contaminating Phospholipids. J. Lipid Res. 2001, 42, 1430–1437. [Google Scholar] [CrossRef]
- Bansal, P.; Gaur, S.N.; Arora, N. Lysophosphatidylcholine Plays Critical Role in Allergic Airway Disease Manifestation. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Aoki, J.; Taira, A.; Takanezawa, Y.; Kishi, Y.; Hama, K.; Kishimoto, T.; Mizuno, K.; Saku, K.; Taguchi, R.; Arai, H. Serum Lysophosphatidic Acid Is Produced through Diverse Phospholipase Pathways. J. Biol. Chem. 2002, 277, 48737–48744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamanti, K.; Cavalli, M.; Pan, G.; Pereira, M.J.; Kumar, C.; Skrtic, S.; Grabherr, M.; Risérus, U.; Eriksson, J.W.; Komorowski, J.; et al. Intra- and Inter-Individual Metabolic Profiling Highlights Carnitine and Lysophosphatidylcholine Pathways as Key Molecular Defects in Type 2 Diabetes. Sci. Rep. 2019, 9, 9653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwatia, M.A.; Doyle, C.B.; Cho, W.; Enhorning, G.; Ackerman, S.J. Combined Activities of Secretory Phospholipases and Eosinophil Lysophospholipases Induce Pulmonary Surfactant Dysfunction by Phospholipid Hydrolysis. J. Allergy Clin. Immunol. 2007, 119, 838–847. [Google Scholar] [CrossRef]
- Weller, P.F.; Wang, H.; Melo, R.C.N. The Charcot–Leyden Crystal Protein Revisited—A Lysopalmitoylphospholipase and More. J. Leukoc. Biol. 2020, 108, 105–112. [Google Scholar] [CrossRef]
- Weller, P.F.; Goetzl, E.J.; Austen, K.F. Identification of Human Eosinophil Lysophospholipase as the Constituent of Charcot-Leyden Crystals. Proc. Natl. Acad. Sci. USA 1980, 77, 7440–7443. [Google Scholar] [CrossRef] [Green Version]
- Nyenhuis, S.M.; Alumkal, P.; Du, J.; Maybruck, B.T.; Vinicky, M.; Ackerman, S.J. Charcot-Leyden Crystal Protein/Galectin-10 Is a Surrogate Biomarker of Eosinophilic Airway Inflammation in Asthma. Biomark. Med. 2019, 13, 715–724. [Google Scholar] [CrossRef]
- Persson, E.K.; Verstraete, K.; Heyndrickx, I.; Gevaert, E.; Aegerter, H.; Percier, J.M.; Deswarte, K.; Verschueren, K.H.G.; Dansercoer, A.; Gras, D.; et al. Protein Crystallization Promotes Type 2 Immunity and Is Reversible by Antibody Treatment. Science 2019, 364, eaaw4295. [Google Scholar] [CrossRef]
- Cunningham, T.J.; Yao, L.; Lucena, A. Product Inhibition of Secreted Phospholipase A2 May Explain Lysophosphatidylcholines’ Unexpected Therapeutic Properties. J. Inflamm. 2008, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Hanna, V.S.; Hafez, E.A.A. Synopsis of Arachidonic Acid Metabolism: A Review. J. Adv. Res. 2018, 11, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Elagizi, A.; Lavie, C.J.; O’keefe, E.; Marshall, K.; O’keefe, J.H.; Milani, R.V. An Update on Omega-3 Polyunsaturated Fatty Acids and Cardiovascular Health. Nutrients 2021, 13, 204. [Google Scholar] [CrossRef]
- Suzumura, A.; Terao, R.; Kaneko, H. Protective Effects and Molecular Signaling of N-3 Fatty Acids on Oxidative Stress and Inflammation in Retinal Diseases. Antioxidants 2020, 9, 920. [Google Scholar] [CrossRef]
- Hutchinson, A.N.; Tingö, L.; Brummer, R.J. The Potential Effects of Probiotics and ω-3 Fatty Acids on Chronic Low-Grade Inflammation. Nutrients 2020, 12, 2402. [Google Scholar] [CrossRef] [PubMed]
- Balić, A.; Vlašić, D.; Žužul, K.; Marinović, B.; Mokos, Z.B. Omega-3 versus Omega-6 Polyunsaturated Fatty Acids in the Prevention and Treatment of Inflammatory Skin Diseases. Int. J. Mol. Sci. 2020, 21, 741. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowska, B.; Maślińska, M. The Place of Omega-3 and Omega-6 Acids in Supplementary Treatment of Inflammatory Joint Diseases. Reumatologia 2020, 58, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Mechanisms of Action of (n-3) Fatty Acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Zou, H.; Yuan, C.; Hong, Y.H.; Kuklev, D.V.; Smith, W.L. Different Fatty Acids Compete with Arachidonic Acid for Binding to the Allosteric or Catalytic Subunits of Cyclooxygenases to Regulate Prostanoid Synthesis. J. Biol. Chem. 2016, 291, 4069–4078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuchiwal, S.K.; Boyce, J.A. Role of Lipid Mediators and Control of Lymphocyte Responses in Type 2 Immunopathology. J. Allergy Clin. Immunol. 2018, 141, 1182–1190. [Google Scholar] [CrossRef] [Green Version]
- Harizi, H.; Corcuff, J.B.; Gualde, N. Arachidonic-Acid-Derived Eicosanoids: Roles in Biology and Immunopathology. Trends Mol. Med. 2008, 14, 461–469. [Google Scholar] [CrossRef]
- Dennis, E.A.; Norris, P.C. Eicosanoid Storm in Infection and Inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Arita, M. Eosinophil Polyunsaturated Fatty Acid Metabolism and Its Potential Control of Inflammation and Allergy. Allergol. Int. 2016, 65, S2–S5. [Google Scholar] [CrossRef] [Green Version]
- Serhan, C.N.; Dalli, J.; Colas, R.A.; Winkler, J.W.; Chiang, N. Protectins and Maresins: New pro-Resolving Families of Mediators in Acute Inflammation and Resolution Bioactive Metabolome. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2015, 1851, 397–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnamoorthy, N.; Abdulnour, R.E.E.; Walker, K.H.; Engstrom, B.D.; Levy, B.D. Specialized Proresolving Mediators in Innate and Adaptive Immune Responses in Airway Diseases. Physiol. Rev. 2018, 98, 1335–1370. [Google Scholar] [CrossRef] [PubMed]
- Isobe, Y.; Arita, M.; Matsueda, S.; Iwamoto, R.; Fujihara, T.; Nakanishi, H.; Taguchi, R.; Masuda, K.; Sasaki, K.; Urabe, D.; et al. Identification and Structure Determination of Novel Anti-Inflammatory Mediator Resolvin E3, 17,18-Dihydroxyeicosapentaenoic Acid. J. Biol. Chem. 2012, 287, 10525–10534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çelik, G.E.; Erkekol, F.O.; Misirligil, Z.; Melli, M. Lipoxin A4 Levels in Asthma: Relation with Disease Severity and Aspirin Sensitivity. Clin. Exp. Allergy 2007, 37, 1494–1501. [Google Scholar] [PubMed]
- Sanak, M.; Levy, B.D.; Clish, C.B.; Chiang, N.; Gronert, K.; Mastalerz, L.; Serhan, C.N.; Szczeklik, A. Aspirin-Tolerant Asthmatics Generate More Lipoxins than Aspirin- Intolerant Asthmatics. Eur. Respir. J. 2000, 16, 44–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.H.; Yin, P.L.; Zhang, Y.M.; Tao, H.X. Reversed Changes of Lipoxin A4 and Leukotrienes in Children with Asthma in Different Severity Degree. Pediatr. Pulmonol. 2010, 45, 333–340. [Google Scholar] [CrossRef]
- Planagumà, A.; Kazani, S.; Marigowda, G.; Haworth, O.; Mariani, T.J.; Israel, E.; Bleecker, E.R.; Curran-Everett, D.; Erzurum, S.C.; Calhoun, W.J.; et al. Airway Lipoxin A4 Generation and Lipoxin A4 Receptor Expression Are Decreased in Severe Asthma. Am. J. Respir. Crit. Care Med. 2008, 178, 574–582. [Google Scholar] [CrossRef] [Green Version]
- Miyata, J.; Fukunaga, K.; Kawashima, Y.; Ohara, O.; Arita, M. Cysteinyl Leukotriene Metabolism of Human Eosinophils in Allergic Disease. Allergol. Int. 2020, 69, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, T.; Nakamura, M.; Shimizu, T. Leukotriene Receptors as Potential Therapeutic Targets. J. Clin. Investig. 2018, 128, 2691–2701. [Google Scholar] [CrossRef]
- Thompson-Souza, G.A.; Gropillo, I.; Neves, J.S. Cysteinyl Leukotrienes in Eosinophil Biology: Functional Roles and Therapeutic Perspectives in Eosinophilic Disorders. Front. Med. 2017, 4, 106. [Google Scholar] [CrossRef] [Green Version]
- Steinke, J.W.; Negri, J.; Payne, S.C.; Borish, L. Biological Effects of Leukotriene E4 on Eosinophils. Prostaglandins Leukot. Essent. Fat. Acids 2014, 91, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Bandeira-Melo, C.; Weller, P.F. Eosinophils and Cysteinyl Leukotrienes. Prostaglandins Leukot. Essent. Fat. Acids 2003, 69, 135–143. [Google Scholar] [CrossRef]
- Chen, K.; Yu, Z.; Yang, J.; Li, H. Expression of Cysteinyl Leukotriene Receptor GPR17 in Eosinophilic and Non-Eosinophilic Chronic Rhinosinusitis with Nasal Polyps. Asian Pacific J. Allergy Immunol. 2018, 36, 93–100. [Google Scholar]
- Liu, M.; Yokomizo, T. The Role of Leukotrienes in Allergic Diseases. Allergol. Int. 2015, 64, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Kamohara, M.; Takasaki, J.; Matsumoto, M.; Saito, T.; Ohishi, T.; Ishii, H.; Furuichi, K. Molecular Cloning and Characterization of Another Leukotriene B4 Receptor. J. Biol. Chem. 2000, 275, 27000–27004. [Google Scholar] [CrossRef]
- Serhan, C.N.; Prescott, S.M. The Scent of a Phagocyte: Advances on Leukotriene B4 Receptors. J. Exp. Med. 2000, 192, F5–F8. [Google Scholar] [CrossRef] [Green Version]
- Yokomizo, T.; Izumi, T.; Chang, K.; Takuwa, Y.; Shimizu, T. A G-Protein-Coupled Receptor for Leukotriene B4 That Mediates Chemotaxis. Nature 1997, 387, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Pal, K.; Feng, X.; Steinke, J.W.; Burdick, M.D.; Shim, Y.M.; Sung, S.S.; Gerald Teague, W.; Borish, L. Leukotriene A4 Hydrolase Activation and Leukotriene B4 Production by Eosinophils in Severe Asthma. Am. J. Respir. Cell Mol. Biol. 2019, 60, 413–419. [Google Scholar] [CrossRef]
- Jo-Watanabe, A.; Okuno, T.; Yokomizo, T. The Role of Leukotrienes as Potential Therapeutic Targets in Allergic Disorders. Int. J. Mol. Sci. 2019, 20, 3580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas Scow, D.; Luttermoser, G.K.; Scott Dickerson, K. Leukotriene Inhibitors in the Treatment of Allergy and Asthma. Am. Fam. Physician 2007, 75, 65–70. [Google Scholar]
- Ujino, M.; Sugimoto, N.; Koizumi, Y.; Ro, S.; Kojima, Y.; Asae, K.H.; Yamashita, N.; Ohta, K.; Nagase, H. Leukotriene Receptor Antagonist Attenuated Airway Inflammation and Hyperresponsiveness in a Double-Stranded RNA-Induced Asthma Exacerbation Model. Allergol. Int. 2017, 66, S21–S26. [Google Scholar] [CrossRef]
- Turner, C.R.; Breslow, R.; Conklyn, M.J.; Andresen, C.J.; Patterson, D.K.; Lopez-Anaya, A.; Owens, B.; Lee, P.; Watson, J.W.; Showell, H.J. In Vitro and in Vivo Effects of Leukotriene B4 Antagonism in a Primate Model of Asthma. J. Clin. Investig. 1996, 97, 381–387. [Google Scholar] [CrossRef]
- Evans, D.J.; Barnes, P.J.; Spaethe, S.M.; Van Alstyne, E.L.; Mitchell, M.I.; O’Connor, B.J. Effect of a Leukotriene B4 Receptor Antagonist, LY293111, on Allergen Induced Responses in Asthma. Thorax 1996, 51, 1178–1184. [Google Scholar] [CrossRef] [Green Version]
- Asanuma, F.; Kuwabara, K.; Arimura, A.; Furue, Y.; Fleisch, J.H.; Hori, Y. Effects of Leukotriene B4 Receptor Antagonist, LY293111Na, on Antigen-Induced Bronchial Hyperresponsiveness and Leukocyte Infiltration in Sensitized Guinea Pigs. Inflamm. Res. 2001, 50, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Gelfand, E.W. Importance of the Leukotriene B4-BLT1 and LTB4-BLT2 Pathways in Asthma. Semin. Immunol. 2017, 33, 44–51. [Google Scholar] [CrossRef]
- Kandhare, A.D.; Liu, Z.; Mukherjee, A.A.; Bodhankar, S.L. Therapeutic Potential of Morin in Ovalbumin-Induced Allergic Asthma Via Modulation of SUMF2/IL-13 and BLT2/NF-KB Signaling Pathway. Curr. Mol. Pharmacol. 2019, 12, 122–138. [Google Scholar] [CrossRef]
- Liu, M.; Shen, J.; Yuan, H.; Chen, F.; Song, H.; Qin, H.; Li, Y.; Xu, J.; Ye, Q.; Li, S.; et al. Leukotriene B4 Receptor 2 Regulates the Proliferation, Migration, and Barrier Integrity of Bronchial Epithelial Cells. J. Cell. Physiol. 2018, 233, 6117–6124. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, Y.; Fukuyama, S.; Okuno, T.; Sasaki, F.; Matsunobu, T.; Asai, Y.; Matsumoto, K.; Saeki, K.; Oike, M.; Sadamura, Y.; et al. Leukotriene B 4 Receptor BLT2 Negatively Regulates Allergic Airway Eosinophilia. FASEB J. 2013, 27, 3306–3314. [Google Scholar] [CrossRef] [PubMed]
- Feltenmark, S.; Gautam, N.; Brunnström, Å.; Griffiths, W.; Backman, L.; Edenius, C.; Lindbom, L.; Björkholm, M.; Claesson, H.E. Eoxins Are Proinflammatory Arachidonic Acid Metabolites Produced via the 15-Lipoxygenase-1 Pathway in Human Eosinophils and Mast Cells. Proc. Natl. Acad. Sci. USA 2008, 105, 680–685. [Google Scholar] [CrossRef] [Green Version]
- Claesson, H.E. On the Biosynthesis and Biological Role of Eoxins and 15-Lipoxygenase-1 in Airway Inflammation and Hodgkin Lymphoma. Prostaglandins Other Lipid Mediat. 2009, 89, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Brunnström, Å.; Tryselius, Y.; Feltenmark, S.; Andersson, E.; Leksell, H.; James, A.; Mannervik, B.; Dahlén, B.; Claesson, H.E. On the Biosynthesis of 15-HETE and Eoxin C4 by Human Airway Epithelial Cells. Prostaglandins Other Lipid Mediat. 2015, 121, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.W.; Balzar, S.; Westcott, J.Y.; Trudeau, J.B.; Sun, Y.; Conrad, D.J.; Wenzel, S.E. Expression and Activation of 15-Lipoxygenase Pathway in Severe Asthma: Relationship to Eosinophilic Phenotype and Collagen Deposition. Clin. Exp. Allergy 2002, 32, 1558–1565. [Google Scholar] [CrossRef]
- Laprise, C.; Sladek, R.; Ponton, A.; Bernier, M.C.; Hudson, T.J.; Laviolette, M. Functional Classes of Bronchial Mucosa Genes That Are Differentially Expressed in Asthma. BMC Genom. 2004, 5, 21. [Google Scholar] [CrossRef]
- Shannon, V.R.; Chanez, P.; Bousquet, J.; Holtzman, M.J. Histochemical Evidence for Induction of Arachidonate 15-Lipoxygenase in Airway Disease. Am. Rev. Respir. Dis. 1993, 147, 1024–1028. [Google Scholar] [CrossRef]
- Sachs-Olsen, C.; Sanak, M.; Lang, A.M.; Gielicz, A.; Mowinckel, P.; Lødrup Carlsen, K.C.; Carlsen, K.H.; Szczeklik, A. Eoxins: A New Inflammatory Pathway in Childhood Asthma. J. Allergy Clin. Immunol. 2010, 126, 859–867.e9. [Google Scholar] [CrossRef]
- Ono, E.; Mita, H.; Taniguchi, M.; Higashi, N.; Hasegawa, M.; Miyazaki, E.; Kumamoto, T.; Akiyama, K. Concentration of 14,15-Leukotriene C4 (Eoxin C4) in Bronchoalveolar Lavage Fluid. Clin. Exp. Allergy 2009, 39, 1348–1352. [Google Scholar] [CrossRef]
- Lee, K.; Lee, S.H.; Kim, T.H. The Biology of Prostaglandins and Their Role as a Target for Allergic Airway Disease Therapy. Int. J. Mol. Sci. 2020, 21, 1851. [Google Scholar] [CrossRef] [Green Version]
- Urade, Y.; Eguchi, N. Lipocalin-Type and Hematopoietic Prostaglandin D Synthases as a Novel Example of Functional Convergence. Prostaglandins Other Lipid Mediat. 2002, 68–69, 375–382. [Google Scholar] [CrossRef]
- Rittchen, S.; Heinemann, A. Therapeutic Potential of Hematopoietic Prostaglandin D2 Synthase in Allergic Inflammation. Cells 2019, 8, 619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peinhaupt, M.; Sturm, E.M.; Heinemann, A. Prostaglandins and Their Receptors in Eosinophil Function and as Therapeutic Targets. Front. Med. 2017, 4, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.Y.; Pillinger, M.H.; Abramson, S.B. Prostaglandin E2 Synthesis and Secretion: The Role of PGE2 Synthases. Clin. Immunol. 2006, 119, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Insuela, D.B.R.; Ferrero, M.R.; de Coutinho, D.S.; Martins, M.A.; Carvalho, V.F. Could Arachidonic Acid-Derived Pro-Resolving Mediators Be a New Therapeutic Strategy for Asthma Therapy? Front. Immunol. 2020, 11, 1. [Google Scholar] [CrossRef]
- Luna-Gomes, T.; Magalhães, K.G.; Mesquita-Santos, F.P.; Bakker-Abreu, I.; Samico, R.F.; Molinaro, R.; Calheiros, A.S.; Diaz, B.L.; Bozza, P.T.; Weller, P.F.; et al. Eosinophils as a Novel Cell Source of Prostaglandin D2: Autocrine Role in Allergic Inflammation. J. Immunol. 2011, 187, 6518–6526. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Ramsden, M.K.; Negri, J.; Baker, M.G.; Payne, S.C.; Borish, L.; Steinke, J.W. Eosinophil Production of Prostaglandin D2 in Patients with Aspirin-Exacerbated Respiratory Disease. J. Allergy Clin. Immunol. 2016, 138, 1089–1097.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okano, M.; Fujiwara, T.; Sugata, Y.; Gotoh, D.; Masaoka, Y.; Sogo, M.; Tanimoto, W.; Yamamoto, M.; Matsumoto, R.; Eguchi, N.; et al. Presence and Characterization of Prostaglandin D2–Related Molecules in Nasal Mucosa of Patients with Allergic Rhinitis. Am. J. Rhinol. 2006, 20, 342–348. [Google Scholar] [CrossRef]
- Suzuki, N.; Ko-Mitamura, E.P.; Inui, T.; Terada, T.; Dejima, K.; Nagata, N.; Urade, Y.; Kawata, R. Steroids Inhibit Eosinophil Accumulation and Downregulate Hematopoietic Chemotaxic Prostaglandin D2 Receptor in Aspirin-Exacerbated Respiratory Disease. Ear Nose Throat J. 2020, 20, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Sedej, M.; Schröder, R.; Bell, K.; Platzer, W.; Vukoja, A.; Kostenis, E.; Heinemann, A.; Waldhoer, M. D-Type Prostanoid Receptor Enhances the Signaling of Chemoattractant Receptor-Homologous Molecule Expressed on TH2 Cells. J. Allergy Clin. Immunol. 2012, 129, 492–500.e9. [Google Scholar] [CrossRef]
- Profita, M.; Sala, A.; Bonanno, A.; Riccobono, L.; Siena, L.; Melis, M.R.; Di Giorgi, R.; Mirabella, F.; Gjomarkaj, M.; Bonsignore, G.; et al. Increased Prostaglandin E2 Concentrations and Cyclooxygenase-2 Expression in Asthmatic Subjects with Sputum Eosinophilia. J. Allergy Clin. Immunol. 2003, 112, 709–716. [Google Scholar] [CrossRef]
- Sastre, B.; Fernández-Nieto, M.; Mollá, R.; López, E.; Lahoz, C.; Sastre, J.; del Pozo, V.; Quirce, S. Increased Prostaglandin E 2 Levels in the Airway of Patients with Eosinophilic Bronchitis. Allergy 2007, 63, 58–66. [Google Scholar]
- Bateman, E.D.; O’Brien, C.; Rugman, P.; Luke, S.; Ivanov, S.; Uddin, M. Efficacy and Safety of the CRTh2 Antagonist AZD1981 as Add-on Therapy to Inhaled Corticosteroids and Long-Acting Β2-Agonists in Patients with Atopic Asthma. Drug Des. Devel. Ther. 2018, 12, 1093–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuge, K.; Inazumi, T.; Shimamoto, A.; Sugimoto, Y. Molecular Mechanisms Underlying Prostaglandin E 2-Exacerbated Inflammation and Immune Diseases. Int. Immunol. 2019, 31, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, R.; Kawada, K.; Sakai, Y. Prostaglandin E2/EP Signaling in the Tumor Microenvironment of Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 6254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolbright, B.L.; Pilbeam, C.C.; Taylor, J.A. Prostaglandin E2 as a Therapeutic Target in Bladder Cancer: From Basic Science to Clinical Trials. Prostaglandins Other Lipid Mediat. 2020, 148, 106409. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.M.; Kleczko, E.K.; Nemenoff, R.A. Eicosanoids in Cancer: New Roles in Immunoregulation. Front. Pharmacol. 2020, 11, 595498. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Moodley, Y.P.; Thompson, P.J.; Misso, N.L. Prostaglandin E2 and Cysteinyl Leukotriene Concentrations in Sputum: Association with Asthma Severity and Eosinophilic Inflammation. Clin. Exp. Allergy 2010, 40, 85–93. [Google Scholar]
- Ricke-Hoch, M.; Stelling, E.; Lasswitz, L.; Kasten, M.; Zapatero-Belinchón, F.J.; Brogden, G.; Gerold, G.; Battmer, K.; Pietschmann, T.; Montiel, V.; et al. SARS-CoV-2-Induced Impaired Immune Response by Prostaglandin E2 Is Accelerated by Age, Male Sex and Air Pollution. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Dorris, S.L.; Peebles, R.S. PGI 2 as a Regulator of Inflammatory Diseases. Mediat. Inflamm. 2012, 2012, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Durchschein, F.; Eherer, A.; Grill, M.; Sturm, E.M.; Pommer, V.; Langner, C.; Högenauer, C.; Schicho, R. Involvement of EP2 and EP4 Receptors in Eosinophilic Esophagitis: A Pilot Study. Dig. Dis. Sci. 2019, 64, 2806–2814. [Google Scholar] [CrossRef] [Green Version]
- Konya, V.; Philipose, S.; Bálint, Z.; Olschewski, A.; Marsche, G.; Sturm, E.M.; Schicho, R.; Peskar, B.A.; Schuligoi, R.; Heinemann, A. Interaction of Eosinophils with Endothelial Cells Is Modulated by Prostaglandin EP4 Receptors. Eur. J. Immunol. 2011, 41, 2379–2389. [Google Scholar] [CrossRef]
- Plaza, J.; Torres, R.; Urbano, A.; Picado, C.; de Mora, F. In Vitro and In Vivo Validation of EP2-Receptor Agonism to Selectively Achieve Inhibition of Mast Cell Activity. Allergy. Asthma Immunol. Res. 2020, 12, 712. [Google Scholar] [CrossRef]
- Nakanishi, M.; Rosenberg, D.W. Multifaceted Roles of PGE2 in Inflammation and Cancer. Semin. Immunopathol. 2013, 35, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Babaev, V.R.; Chew, J.D.; Ding, L.; Davis, S.; Breyer, M.D.; Breyer, R.M.; Oates, J.A.; Fazio, S.; Linton, M.F. Macrophage EP4 Deficiency Increases Apoptosis and Suppresses Early Atherosclerosis. Cell Metab. 2008, 8, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Coulombe, F.; Jaworska, J.; Verway, M.; Tzelepis, F.; Massoud, A.; Gillard, J.; Wong, G.; Kobinger, G.; Xing, Z.; Couture, C.; et al. Targeted Prostaglandin E2 Inhibition Enhances Antiviral Immunity through Induction of Type I Interferon and Apoptosis in Macrophages. Immunity 2014, 40, 554–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergqvist, F.; Morgenstern, R.; Jakobsson, P.J. A Review on MPGES-1 Inhibitors: From Preclinical Studies to Clinical Applications. Prostaglandins Other Lipid Mediat. 2020, 147, 106383. [Google Scholar] [CrossRef]
- Sheibanie, A.F.; Khayrullina, T.; Safadi, F.F.; Ganea, D. Prostaglandin E2 Exacerbates Collagen-Induced Arthritis in Mice through the Inflammatory Interleukin-23/Interleukin-17 Axis. Arthritis Rheum. 2007, 56, 2608–2619. [Google Scholar] [CrossRef] [PubMed]
- Schölkens, B.A.; Bartmann, W.; Beck, G.; Lerch, U.; Konz, E.; Weithmann, U. Vasodilation and Inhibition of Platelet Aggregation by Prostacyclins with Modified ω-Side Chain. Prostaglandines Med. 1979, 3, 7–22. [Google Scholar] [CrossRef]
- Kuang, H.; Li, Q.; Yi, Q.; Lu, T. The Efficacy and Safety of Aerosolized Iloprost in Pulmonary Arterial Hypertension: A Systematic Review and Meta-Analysis. Am. J. Cardiovasc. Drugs 2019, 19, 393–401. [Google Scholar] [CrossRef]
- Johansson, P.I.; Bestle, M.; Søe-Jensen, P.; Kristiansen, K.T.; Stensballe, J.; Clausen, N.E.; Perner, A. The Effect of Prostacyclin (Iloprost) Infusion at a Dose of 1 Ng/Kg/Min for 72 Hours Compared to Placebo in Mechanically Ventilated Patients with COVID-19: A Structured Summary of a Study Protocol for a Randomized Controlled Trial. Trials 2020, 21, 746. [Google Scholar] [CrossRef] [PubMed]
- Monneret, G.; Gravel, S.; Diamond, M.; Rokach, J.; Powell, W.S. Prostaglandin D2 Is a Potent Chemoattractant for Human Eosinophils That Acts via a Novel DP Receptor. Blood 2001, 98, 1942–1948. [Google Scholar] [CrossRef]
- Powell, W.S.; Chung, D.; Gravel, S. 5-Oxo-6,8,11,14-Eicosatetraenoic Acid Is a Potent Stimulator of Human Eosinophil Migration. J. Immunol. 1995, 154, 4123 LP–4132 LP. [Google Scholar]
- Simons, B.; Ferrini, M.E.; Carvalho, S.; Bassett, D.J.P.; Jaffar, Z.; Roberts, K. PGI 2 Controls Pulmonary NK Cells That Prevent Airway Sensitization to House Dust Mite Allergen. J. Immunol. 2017, 198, 461–471. [Google Scholar] [CrossRef] [Green Version]
- Goetzl, E.J.; Goldman, D.W.; Naccache, P.H.; Sha’afi, R.I.; Pickett, W.C. Mediation of Leukocyte Components of Inflammatory Reactions by Lipoxygenase Products of Arachidonic Acid. Adv. Prostaglandin. Thromboxane. Leukot. Res. 1982, 9, 273–282. [Google Scholar] [PubMed]
- Powell, W.S.; Rokach, J.; Biophys, B.; Author, A. Biosynthesis, Biological Effects, and Receptors of Hydroxyeicosatetraenoic Acids (HETEs) and Oxoeicosatetraenoic Acids (Oxo-ETEs) Derived from Arachidonic Acid HHS Public Access Author Manuscript. Biochim. Biophys. Acta 2015, 1851, 340–355. [Google Scholar] [CrossRef] [Green Version]
- Kowal, K.; Gielicz, A.; Sanak, M. The Effect of Allergen-Induced Bronchoconstriction on Concentration of 5-Oxo-ETE in Exhaled Breath Condensate of House Dust Mite-Allergic Patients. Clin. Exp. Allergy 2017, 47, 1253–1262. [Google Scholar] [CrossRef]
- Jones, C.E.; Holden, S.; Tenaillon, L.; Bhatia, U.; Seuwen, K.; Tranter, P.; Turner, J.; Kettle, R.; Bouhelal, R.; Charlton, S.; et al. Expression and Characterization of a 5-Oxo-6 E,8 Z,11 Z,14 Z-Eicosatetraenoic Acid Receptor Highly Expressed on Human Eosinophils and Neutrophils. Mol. Pharmacol. 2003, 63, 471–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosoi, T.; Koguchi, Y.; Sugikawa, E.; Chikada, A.; Ogawa, K.; Tsuda, N.; Suto, N.; Tsunoda, S.; Taniguchi, T.; Ohnuki, T. Identification of a Novel Human Eicosanoid Receptor Coupled to Gi/O. J. Biol. Chem. 2002, 277, 31459–31465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, W.S.; Rokach, J. The Eosinophil Chemoattractant 5-Oxo-ETE and the OXE Receptor. Prog. Lipid Res. 2013, 52, 651–665. [Google Scholar] [CrossRef] [Green Version]
- Urasaki, T.; Takasaki, J.; Nagasawa, T.; Ninomiya, H. Pivotal Role of 5-Lipoxygenase in the Activation of Human Eosinophils: Platelet-Activating Factor and Interleukin-5 Induce CD69 on Eosinophils through the 5-Lipoxygenase Pathway. J. Leukoc. Biol. 2001, 69, 105–112. [Google Scholar] [PubMed]
- O’Flaherty, J.T.; Kuroki, M.; Nixon, A.B.; Wijkander, J.; Yee, E.; Lee, S.L.; Smitherman, P.K.; Wykle, R.L.; Daniel, L.W. 5-Oxo-Eicosatetraenoate Is a Broadly Active, Eosinophil-Selective Stimulus for Human Granulocytes. J. Immunol. 1996, 157, 336–342. [Google Scholar]
- Langlois, A.; Chouinard, F.; Flamand, N.; Ferland, C.; Rola-Pleszczynski, M.; Laviolette, M. Crucial Implication of Protein Kinase C (PKC)-, PKC-, ERK-1/2, and P38 MAPK in Migration of Human Asthmatic Eosinophils. J. Leukoc. Biol. 2009, 85, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Guilbert, M.; Ferland, C.; Bossé, M.; Flamand, N.; Lavigne, S.; Laviolette, M. 5-Oxo-6,8,11,14-Eicosatetraenoic Acid Induces Important Eosinophil Transmigration through Basement Membrane Components: Comparison of Normal and Asthmatic Eosinophils. Am. J. Respir. Cell Mol. Biol. 1999, 21, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Patel, K.; Davison, J.; Pittman, Q.; Sharkey, K. Cannabinoid CB2 Receptors in Health and Disease. Curr. Med. Chem. 2010, 17, 1394–1410. [Google Scholar] [CrossRef]
- Zoppi, S.; Madrigal, J.L.; Caso, J.R.; García-Gutiérrez, M.S.; Manzanares, J.; Leza, J.C.; García-Bueno, B. Regulatory Role of the Cannabinoid CB2 Receptor in Stress-Induced Neuroinflammation in Mice. Br. J. Pharmacol. 2014, 171, 2814–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, W.S.; Rokach, J. Targeting the OXE Receptor as a Potential Novel Therapy for Asthma. Biochem. Pharmacol. 2020, 179, 113930. [Google Scholar] [CrossRef]
- Kadamur, G.; Ross, E.M. Mammalian Phospholipase C. Annu. Rev. Physiol. 2013, 75, 127–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, M.J. The Inositol Trisphosphate/Calcium Signaling Pathway in Health and Disease. Physiol. Rev. 2016, 96, 1261–1296. [Google Scholar] [CrossRef] [Green Version]
- Kroegel, C.; Chilvers, E.R.; Giembycz, M.A.; Challiss, R.A.J.; Barnes, P.J. Platelet-Activating Factor Stimulates a Rapid Accumulation of Inositol (1,4,5)Trisphosphate in Guinea Pig Eosinophils: Relationship to Calcium Mobilization and Degranulation. J. Allergy Clin. Immunol. 1991, 88, 114–124. [Google Scholar] [CrossRef]
- Sano, A.; Zhu, X.; Sano, H.; Muñoz, N.M.; Boetticher, E.; Leff, A.R. Regulation of Eosinophil Function by Phosphatidylinositol-Specific PLC and Cytosolic PLA 2. Am. J. Physiol. Cell. Mol. Physiol. 2001, 281, L844–L851. [Google Scholar] [CrossRef] [Green Version]
- Nakao, M.; Sugaya, M.; Suga, H.; Kawaguchi, M.; Morimura, S.; Kai, H.; Ohmatsu, H.; Fujita, H.; Asano, Y.; Tada, Y.; et al. Serum Autotaxin Levels Correlate with Pruritus in Patients with Atopic Dermatitis. J. Investig. Dermatol. 2014, 134, 1745–1747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maher, T.M.; Kreuter, M.; Lederer, D.J.; Brown, K.K.; Wuyts, W.; Verbruggen, N.; Stutvoet, S.; Fieuw, A.; Ford, P.; Abi-Saab, W.; et al. Rationale, Design and Objectives of Two Phase III, Randomised, Placebo-Controlled Studies of GLPG1690, a Novel Autotaxin Inhibitor, in Idiopathic Pulmonary Fibrosis (ISABELA 1 and 2). BMJ Open Respir. Res. 2019, 6, e000422. [Google Scholar] [CrossRef] [PubMed]
- Reisenberg, M.; Singh, P.K.; Williams, G.; Doherty, P. The Diacylglycerol Lipases: Structure, Regulation and Roles in and beyond Endocannabinoid Signalling. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 3264–3275. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, C.; Archambault, A.S.; Dumais, É.; Martin, C.; Blanchet, M.R.; Bissonnette, E.; Ohashi, N.; Yamamoto, K.; Itoh, T.; Laviolette, M.; et al. Endocannabinoid Hydrolysis Inhibition Unmasks That Unsaturated Fatty Acids Induce a Robust Biosynthesis of 2-Arachidonoyl-Glycerol and Its Congeners in Human Myeloid Leukocytes. FASEB J. 2020, 34, 4253–4265. [Google Scholar] [CrossRef] [Green Version]
- Angelina, A.; Pérez-Diego, M.; López-Abente, J.; Palomares, O. The Role of Cannabinoids in Allergic Diseases: Collegium Internationale Allergologicum (CIA) Update 2020. Int. Arch. Allergy Immunol. 2020, 181, 565–584. [Google Scholar] [CrossRef]
- Kondo, M.; Tezuka, T.; Ogawa, H.; Koyama, K.; Bando, H.; Azuma, M.; Nishioka, Y. Lysophosphatidic Acid Regulates the Differentiation of Th2 Cells and Its Antagonist Suppresses Allergic Airway Inflammation. Int. Arch. Allergy Immunol. 2021, 182, 1–13. [Google Scholar] [CrossRef]
- Kim, S.J.; Moon, H.G.; Park, G.Y. The Roles of Autotaxin/Lysophosphatidic Acid in Immune Regulation and Asthma. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158641. [Google Scholar] [CrossRef]
- Nieto-Posadas, A.; Picazo-Juárez, G.; Llorente, I.; Jara-Oseguera, A.; Morales-Lázaro, S.; Escalante-Alcalde, D.; Islas, L.D.; Rosenbaum, T. Lysophosphatidic Acid Directly Activates TRPV1 through a C-Terminal Binding Site. Nat. Chem. Biol. 2012, 8, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Ikeda, S.; Kishimoto, S.; Gokoh, M.; Yanagimoto, S.; Waku, K.; Sugiura, T. 2-Arachidonoylglycerol, an Endogenous Cannabinoid Receptor Ligand, Induces the Migration of EoL-1 Human Eosinophilic Leukemia Cells and Human Peripheral Blood Eosinophils. J. Leukoc. Biol. 2004, 76, 1002–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, S.; Oka, S.; Gokoh, M.; Sugiura, T. Chemotaxis of Human Peripheral Blood Eosinophils to 2-Arachidonoylglycerol: Comparison with Other Eosinophil Chemoattractants. In Proceedings of the International Archives of Allergy and Immunology. Int. Arch. Allergy Immunol. 2006, 140, 3–7. [Google Scholar] [CrossRef]
- Rai, V.; Touré, F.; Chitayat, S.; Pei, R.; Song, F.; Li, Q.; Zhang, J.; Rosario, R.; Ramasamy, R.; Chazin, W.J.; et al. Lysophosphatidic Acid Targets Vascular and Oncogenic Pathways via RAGE Signaling. J. Exp. Med. 2012, 209, 2339–2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Baker, D.L.; Yasuda, S.; Makarova, N.; Balazs, L.; Johnson, L.R.; Marathe, G.K.; McIntyre, T.M.; Xu, Y.; Prestwich, G.D.; et al. Lysophosphatidic Acid Induces Neointima Formation Through PPARγ Activation. J. Exp. Med. 2004, 199, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Oka, S.; Wakui, J.; Ikeda, S.; Yanagimoto, S.; Kishimoto, S.; Gokoh, M.; Nasui, M.; Sugiura, T. Involvement of the Cannabinoid CB2 Receptor and Its Endogenous Ligand 2-Arachidonoylglycerol in Oxazolone-Induced Contact Dermatitis in Mice. J. Immunol. 2006, 177, 8796–8805. [Google Scholar] [CrossRef] [PubMed]
- Maresz, K.; Carrier, E.J.; Ponomarev, E.D.; Hillard, C.J.; Dittel, B.N. Modulation of the Cannabinoid CB2 Receptor in Microglial Cells in Response to Inflammatory Stimuli. J. Neurochem. 2005, 95, 437–445. [Google Scholar] [CrossRef]
- Steffens, S.; Veillard, N.R.; Arnaud, C.; Pelli, G.; Burger, F.; Staub, C.; Zimmer, A.; Frossard, J.L.; Mach, F. Low Dose Oral Cannabinoid Therapy Reduces Progression of Atherosclerosis in Mice. Nature 2005, 434, 782–786. [Google Scholar] [CrossRef]
- Ackerman, S.J.; Park, G.Y.; Christman, J.W.; Nyenhuis, S.; Berdyshev, E.; Natarajan, V. Polyunsaturated Lysophosphatidic Acid as a Potential Asthma Biomarker. Biomark. Med. 2016, 10, 123–135. [Google Scholar] [CrossRef]
- Shimizu, Y.; Murao, K.; Tanaka, T.; Kubo, Y.; Tokumura, A. Increased Lysophospholipase D Activity of Autotaxin in Sera of Patients with Atopic Dermatitis. J. Dermatol. Sci. 2014, 74, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Jun, Y.J.; Lee, K.J.; Hwang, S.M.; Kim, T.H.; Lee, S.H.; Lee, S.H. Chronic Rhinosinusitis with Nasal Polyps and without Nasal Polyps Is Associated with Increased Expression of Lysophosphatidic Acid–Related Molecules. Am. J. Rhinol. Allergy 2014, 28, 199–207. [Google Scholar] [CrossRef]
- Knowlden, S.; Georas, S.N. The Autotaxin–LPA Axis Emerges as a Novel Regulator of Lymphocyte Homing and Inflammation. J. Immunol. 2014, 192, 851–857. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Tong, J.; He, D.; Pendyala, S.; Evgeny, B.; Chun, J.; Sperling, A.I.; Natarajan, V. Role of Lysophosphatidic Acid Receptor LPA 2 in the Development of Allergic Airway Inflammation in a Murine Model of Asthma. Respir. Res. 2009, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Oczypok, E.A.; Perkins, T.N.; Oury, T.D. All the “RAGE” in Lung Disease: The Receptor for Advanced Glycation Endproducts (RAGE) Is a Major Mediator of Pulmonary Inflammatory Responses. Paediatr. Respir. Rev. 2017, 23, 40–49. [Google Scholar] [CrossRef]
- Perkins, T.N.; Donnell, M.L.; Oury, T.D. The Axis of the Receptor for Advanced Glycation Endproducts in Asthma and Allergic Airway Disease. Allergy 2020, 14600. [Google Scholar] [CrossRef]
- Milutinovic, P.S.; Alcorn, J.F.; Englert, J.M.; Crum, L.T.; Oury, T.D. The Receptor for Advanced Glycation End Products Is a Central Mediator of Asthma Pathogenesis. Am. J. Pathol. 2012, 181, 1215–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, M.A.; Loh, Z.; Gan, W.J.; Zhang, V.; Yang, H.; Li, J.H.; Yamamoto, Y.; Schmidt, A.M.; Armour, C.L.; Hughes, J.M.; et al. Receptor for Advanced Glycation End Products and Its Ligand High-Mobility Group Box-1 Mediate Allergic Airway Sensitization and Airway Inflammation. J. Allergy Clin. Immunol. 2014, 134, 440–450.e3. [Google Scholar] [CrossRef] [PubMed]
- Brandt, E.B.; Lewkowich, I.P. RAGE-Induced Asthma: A Role for the Receptor for Advanced Glycation End-Products in Promoting Allergic Airway Disease. J. Allergy Clin. Immunol. 2019, 144, 651–653. [Google Scholar] [CrossRef] [Green Version]
- Curran, C.S.; Bertics, P.J. Human Eosinophils Express RAGE, Produce RAGE Ligands, Exhibit PKC-Delta Phosphorylation and Enhanced Viability in Response to the RAGE Ligand, S100B. Int. Immunol. 2011, 23, 713–728. [Google Scholar] [CrossRef] [Green Version]
- Dyer, K.D.; Rosenberg, H.F. Physiologic Concentrations of HMGB1 Have No Impact on Cytokine-Mediated Eosinophil Survival or Chemotaxis in Response to Eotaxin-2 (CCL24). PLoS ONE 2015, 10, e0118887. [Google Scholar] [CrossRef]
- Wood, L.G.; Gibson, P.G.; Garg, M.L. Biomarkers of Lipid Peroxidation, Airway Inflammation and Asthma. Eur. Respir. J. 2003, 21, 177–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, S.J. Bronchial Epithelial Cell-Cytokine Interactions in Airway Inflammation. J. Investig. Med. Off. Publ. Am. Fed. Clin. Res. 1995, 43, 241–249. [Google Scholar]
- Doelman, C.J.A.; Bast, A. Oxygen Radicals in Lung Pathology. Free Radic. Biol. Med. 1990, 9, 381–400. [Google Scholar] [CrossRef]
- Dweik, R.A.; Comhair, S.A.A.; Gaston, B.; Thunnissen, F.B.J.M.; Farver, C.; Thomassen, M.J.; Kavuru, M.; Hammel, J.; Abu-Soud, H.M.; Erzurum, S.C. NO Chemical Events in the Human Airway during the Immediate and Late Antigen-Induced Asthmatic Response. Proc. Natl. Acad. Sci. USA 2001, 98, 2622–2627. [Google Scholar] [CrossRef] [Green Version]
- MacPherson, J.C.; Comhair, S.A.A.; Erzurum, S.C.; Klein, D.F.; Lipscomb, M.F.; Kavuru, M.S.; Samoszuk, M.K.; Hazen, S.L. Eosinophils Are a Major Source of Nitric Oxide-Derived Oxidants in Severe Asthma: Characterization of Pathways Available to Eosinophils for Generating Reactive Nitrogen Species. J. Immunol. 2001, 166, 5763–5772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comhair, S.A.A.; Erzurum, S.C. Redox Control of Asthma: Molecular Mechanisms and Therapeutic Opportunities. Antioxid. Redox Signal. 2010, 12, 93–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulsmann, A.R.; Raatgeep, H.R.; Den Hollander, J.C.; Stijnen, T.; Saxena, P.R.; Kerrebijn, K.F.; De Jongste, J.C. Oxidative Epithelial Damage Produces Hyperresponsiveness of Human Peripheral Airways. Am. J. Respir. Crit. Care Med. 1994, 149, 519–525. [Google Scholar] [CrossRef]
- Qu, J.; Li, Y.; Zhong, W.; Gao, P.; Hu, C. Recent Developments in the Role of Reactive Oxygen Species in Allergic Asthma. J. Thorac. Dis. 2017, 9, E32–E43. [Google Scholar] [CrossRef] [Green Version]
- Petreccia, D.C.; Nauseef, W.M.; Clark, R.A. Respiratory Burst of Normal Human Eosinophils. J. Leukoc. Biol. 1987, 41, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; DiDonato, J.A.; Buffa, J.; Comhair, S.A.; Aronica, M.A.; Dweik, R.A.; Lee, N.A.; Lee, J.J.; Thomassen, M.J.; Kavuru, M.; et al. Eosinophil Peroxidase Catalyzed Protein Carbamylation Participates in Asthma. J. Biol. Chem. 2016, 291, 22118–22135. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.N.; Slungaard, A.; Hazen, S.L. Redox Report Communications in Free Radical Research Role of Eosinophil Peroxidase in the Origins of Protein Oxidation in Asthma. Redox Rep. 2013. [Google Scholar] [CrossRef]
- Sedgwick, J.B.; Geiger, K.M.; Busse, W.W. Superoxide Generation by Hypodense Eosinophils from Patients with Asthma. Am. Rev. Respir. Dis. 1990, 142, 120–125. [Google Scholar] [CrossRef]
- Lavinskiene, S.; Malakauskas, K.; Jeroch, J.; Hoppenot, D.; Sakalauskas, R. Functional Activity of Peripheral Blood Eosinophils in Allergen-Induced Late-Phase Airway Inflammation in Asthma Patients. J. Inflamm. 2015, 12, 25. [Google Scholar] [CrossRef] [Green Version]
- Dworski, R. Oxidant Stress in Asthma. Thorax 2000, 55, 51S–53S. [Google Scholar] [CrossRef] [Green Version]
- Silveira, J.S.; Antunes, G.L.; Kaiber, D.B.; da Costa, M.S.; Marques, E.P.; Ferreira, F.S.; Gassen, R.B.; Breda, R.V.; Wyse, A.T.S.; Pitrez, P.; et al. Reactive Oxygen Species Are Involved in Eosinophil Extracellular Traps Release and in Airway Inflammation in Asthma. J. Cell. Physiol. 2019, 234, 23633–23646. [Google Scholar] [CrossRef] [PubMed]
- Repetto, M.; Semprine, J.; Boveris, A. Lipid Peroxidation: Chemical Mechanism, Biological Implications and Analytical Determination. In Lipid Peroxidation; InTech: Londond, UK, 2012. [Google Scholar]
- Bultel-Poncé, V.; Durand, T.; Guy, A.; Oger, C.; Galano, J.-M. Non Enzymatic Metabolites of Polyunsaturated Fatty Acids: Friend or Foe. OCL 2016, 23, D118. [Google Scholar] [CrossRef] [Green Version]
- Leung, K.S.; Galano, J.M.; Durand, T.; Lee, J.C.Y. Current Development in Non-Enzymatic Lipid Peroxidation Products, Isoprostanoids and Isofuranoids, in Novel Biological Samples. Free Radic. Res. 2015, 49, 816–826. [Google Scholar] [CrossRef]
- Vigor, C.; Bertrand-Michel, J.; Pinot, E.; Oger, C.; Vercauteren, J.; Le Faouder, P.; Galano, J.M.; Lee, J.C.Y.; Durand, T. Non-Enzymatic Lipid Oxidation Products in Biological Systems: ASSESSMENT of the Metabolites from Polyunsaturated Fatty Acids. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 964, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Dworski, R.; Murray, J.J.; Roberts, L.J.; Oates, J.A.; Morrow, J.D.; Fisher, L.; Sheller, J.R. Allergen-Induced Synthesis of F2-Isoprostanes in Atopic Asthmatics: Evidence for Oxidant Stress. Am. J. Respir. Crit. Care Med. 1999, 160, 1947–1951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, T.K.; Loh, X.Y.; Peh, H.Y.; Tan, W.N.F.; Tan, W.S.D.; Li, N.; Tay, I.J.J.; Wong, W.S.F.; Engelward, B.P. House Dust Mite–Induced Asthma Causes Oxidative Damage and DNA Double-Strand Breaks in the Lungs. J. Allergy Clin. Immunol. 2016, 138, 84–96.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montuschi, P.; Corradi, M.; Ciabattoni, G.; Nightingale, J.; Kharitonov, S.A.; Barnes, P.J. Increased 8-Isoprostane, a Marker of Oxidative Stress, in Exhaled Condensate of Asthma Patients. Am. J. Respir. Crit. Care Med. 1999, 160, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Montuschi, P.; Barnes, P.J. Isoprostanes and Asthma. Drug Discov. Today Ther. Strateg. 2006, 3, 287–292. [Google Scholar] [CrossRef]
- Shahid, S.K.; Kharitonov, S.A.; Wilson, N.M.; Bush, A.; Barnes, P.J. Exhaled 8-Isoprostane in Childhood Asthma. Respir. Res. 2005, 6, 79. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Zhang, Q.; Zhong, N.; Lai, K. BAL Fluid 8-Isoprostane Concentrations in Eosinophilic Bronchitis and Asthma. J. Asthma 2009, 46, 712–715. [Google Scholar] [CrossRef]
- Groot, L.E.S.; Sabogal Piñeros, Y.S.; Bal, S.M.; Pol, M.A.; Hamann, J.; Sterk, P.J.; Kulik, W.; Lutter, R. Do Eosinophils Contribute to Oxidative Stress in Mild Asthma? Clin. Exp. Allergy 2019, 49, 929–931. [Google Scholar] [CrossRef]
- M Leung, D.Y.; Ledford, D.K.; Moreno-Macias, H.; Romieu, I.; Mexico City, S. Clinical Reviews in Allergy and Immunology Effects of Antioxidant Supplements and Nutrients on Patients with Asthma and Allergies. J. Allergy Clin. Immunol. 2014, 133, 1237–1281. [Google Scholar]
- Patel, B.D.; Welch, A.A.; Bingham, S.A.; Luben, R.N.; Day, N.E.; Khaw, K.T.; Lomas, D.A.; Wareham, N.J. Dietary Antioxidants and Asthma in Adults. Thorax 2006, 61, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.-Y.; Ni, Z.-H.; Luo, X.-M.; Wang, X.-B. Advance of Antioxidants in Asthma Treatment. World J. Respirol. 2017, 7, 17. [Google Scholar] [CrossRef]
- Burton, G.W.; Joyce, A.; Ingold, K.U. Is Vitamin E the Only Lipid-Soluble, Chain-Breaking Antioxidant in Human Blood Plasma and Erythrocyte Membranes? Arch. Biochem. Biophys. 1983, 221, 281–290. [Google Scholar] [CrossRef]
- Smit, H.A.; Grievink, L.; Tabak, C. Dietary Influences on Chronic Obstructive Lung Disease and Asthma: A Review of the Epidemiological Evidence. In Nutrition Society; CAB International: Oxfordshire, UK, 1999; Volume 58, pp. 309–319. [Google Scholar]
- Dow, L.; Tracey, M.; Villar, A.; Coggon, D.; Margetts, B.M.; Campbell, M.J.; Holgate, S.T. Does Dietary Intake of Vitamins C and E Influence Lung Function in Older People? Am. J. Respir. Crit. Care Med. 1996, 154, 1401–1404. [Google Scholar] [CrossRef] [PubMed]
- Berdnikovs, S.; Abdala-Valencia, H.; McCary, C.; Somand, M.; Cole, R.; Garcia, A.; Bryce, P.; Cook-Mills, J.M. Isoforms of Vitamin E Have Opposing Immunoregulatory Functions during Inflammation by Regulating Leukocyte Recruitment. J. Immunol. 2009, 182, 4395–4405. [Google Scholar] [CrossRef] [PubMed] [Green Version]



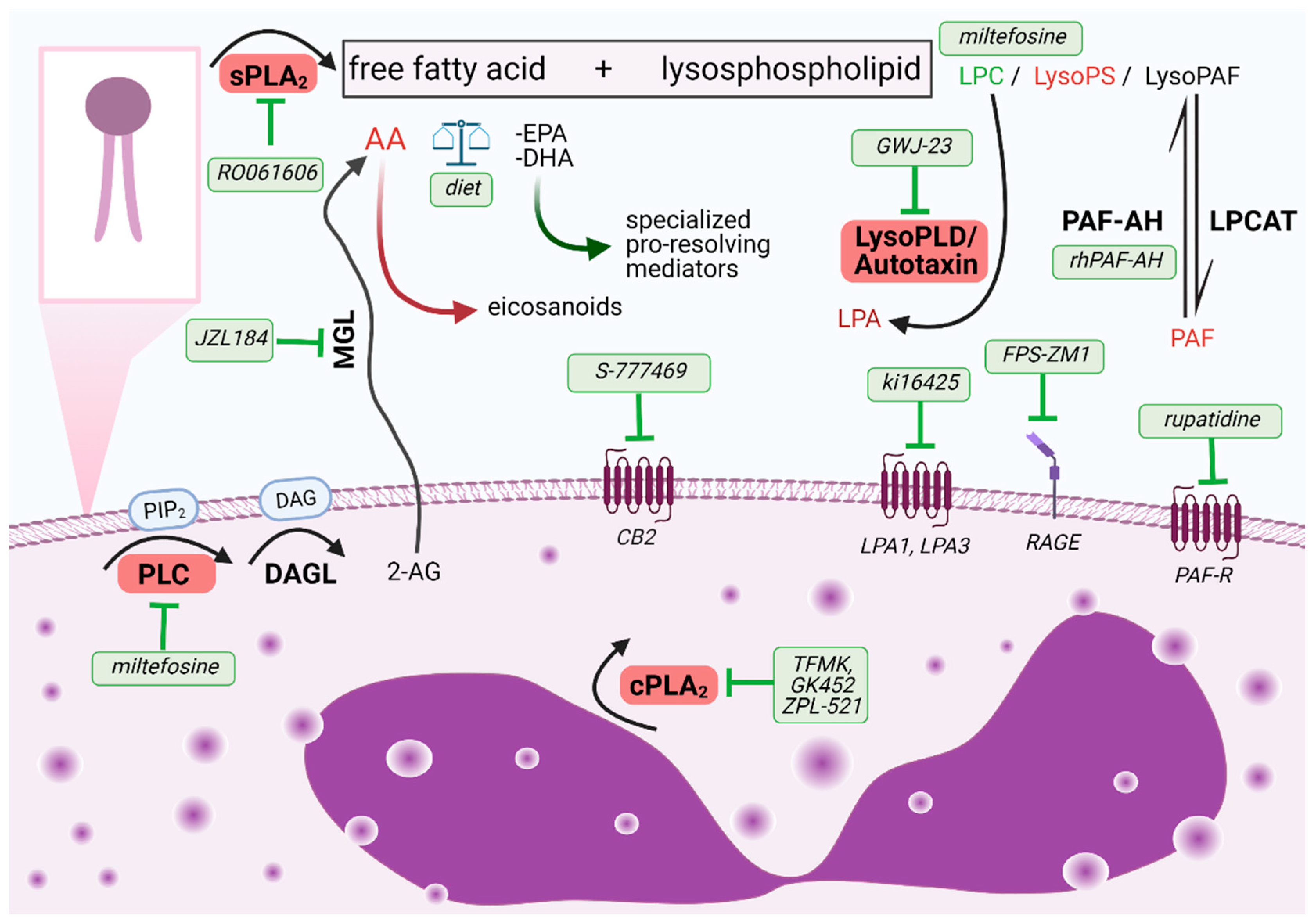{kind=link}
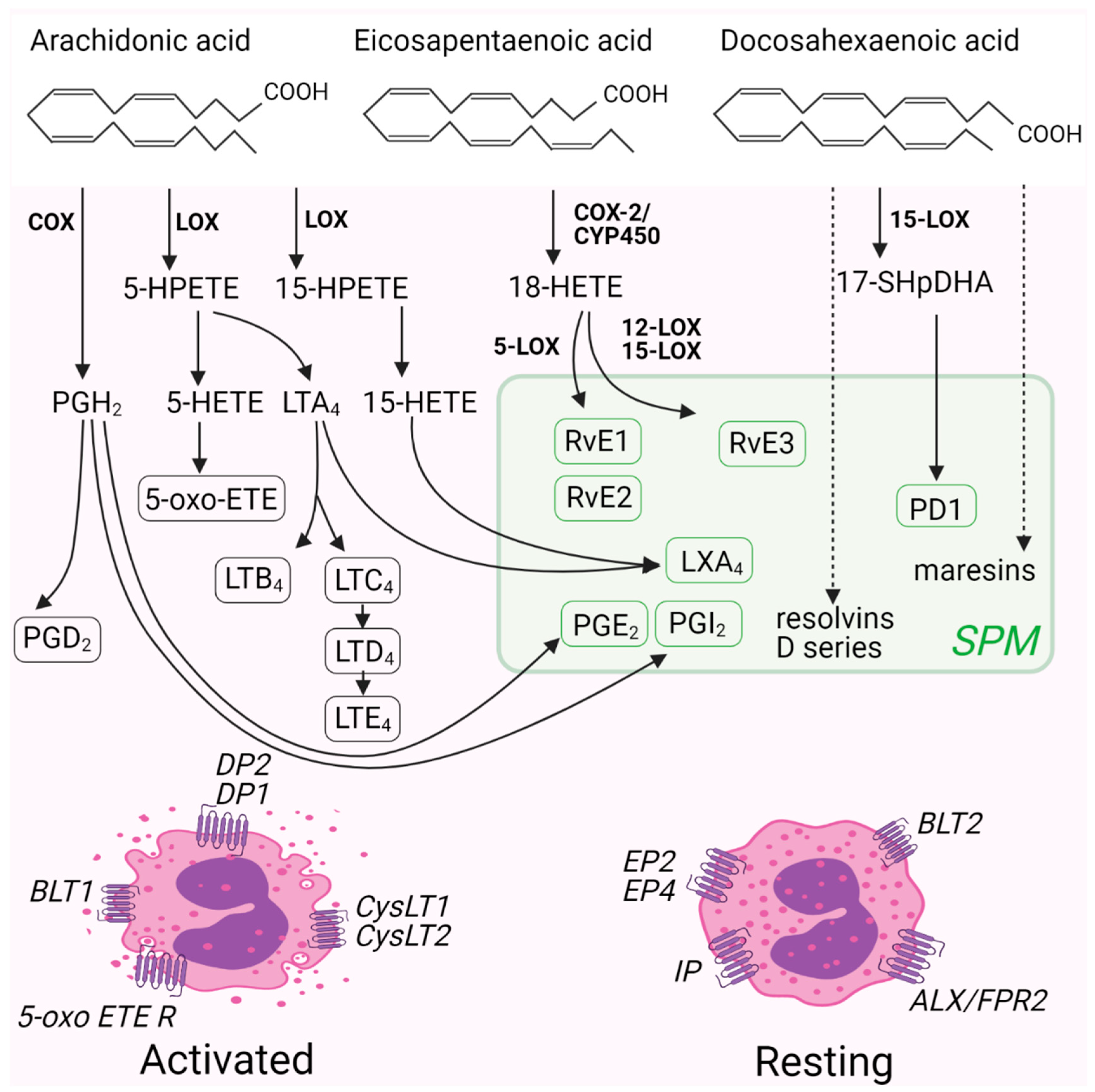{kind=link}
| Compound | Mode of Action | Published Effects | References |
|---|---|---|---|
| RO061606 ROC-0929 | inhibitor of sPLA2-X | Inhibition of eicosanoid formation. Decrease in allergen-induced airway inflammation, mucus hypersecretion and hyperresponsiveness. | [40,42] |
| trifluoromethyl ketone | inhibitor of cPLA2 | Inhibition of antigen-induced airway eosinophil accumulation and airway hyperresponsiveness to methacholine. | [47] |
| ZPL-521 | inhibitor of cPLA2 | Clinical data: efficacious for treatment of atopic dermatitis:improvement on the eczema severity score, safe, well tolerated. | [48] |
| GK452, GK504, GK484 | inhibitors of cPLA2 with improved ADME properties | Decreased eicosanoid production. | [49,50] |
| rhPAF-AH | hydrolysis of PAF to lysoPAF | Preclinical model: significant reduction in airway eosinophil infiltration, mucus hypersecretion, and airway hyperreactivity in response to methacholine challenge. Clinical data: No improvement in symptoms in mild asthma. | [51,52] |
| rupatadine | dual antagonist of H1 receptor and PAF receptor | In use for the treatment of urticaria and allergic rhinitis in adults and children. Reduces the levels of infiltrating eosinophils in allergic rhinitis. | [53,54] |
| miltefosine | lipid raft modulator, PLC inhibitor | Inhibits activation, degranulation, and migration of human eosinophils; decreases immune cell infiltration and improves lung function parameters in a model of allergic lung inflammation. Clinical data: efficacious and safe in an antihistamine-resistant chronic spontaneous urticaria. | [55,56] |
| resolvinE3 (RvE3),18-deoxy-RvE3 | antagonism of BLT1 | Decreased immune cell infiltration in a model of allergic inflammation, improvement in lung function parameters. Decreased polymorphonuclear leukocytes (PMNs) in the peritoneal exudates. | [57,58] |
| protectin D1 | proposed action on GPR37 | Decreased eosinophil recruitment to the lungs, improvement in lung function parameters and airway mucus secretion. Accelerated resolution of airway inflammation. | [59,60] |
| stable LXA4 analogues | ALX/FPR2 receptor agonist | Preclinical: Inhibition of airway hyperresponsiveness and inflammation; Clinical: well tolerated, topical- reduces the severity of infantile eczema, inhaled- improves lung function in mild asthmatics | [61,62,63] |
| S-777469 | CB2 receptor agonist | Preclinical: inhibition of skin inflammation and eosinophil infiltration in mice by blocking the activities of 2-AG. Phase Ib/IIa clinical study for atopic dermatitis ongoing (NCT00697710). | [64] |
| JZL184 | MGL inhibitor | Inhibition of eosinophil migration induced by 2-AG in vitro. Inhibition of airway hyperreactivity and airway inflammation in an LPS-induced model of airway inflammation. | [65,66] |
| GWJ-23 | LysoPLD (autotaxin) inhibitor | Reduced inflammation in a triple-allergen mouse asthma model | [67] |
| ki16425 | antagonist of LPA receptor 1 and 3 | Prevents bradykinin induced asthmatic bronchoconstriction and reduces associated respiratory distress following allergen exposure | [68] |
| FPS-ZM1 | RAGE receptor antagonist | Inhibits rIL-5/13–mediated airway mucus metaplasia and allergic inflammation, including eosinophil infiltration. | [69] |
| zileuton | 5-LO inhibitor | In use: prophylaxis and treatment of chronic asthma | [70] |
| montelukast zafirlukast | Cys-LT1 receptor antagonist | In use: prophylaxis and chronic treatment of asthma, the prevention of exercise-induced bronchospasm, and the relief of symptoms of allergic rhinitis. | [71] |
| TAS-205 HQL-79 | hPGDS inhibitor | In combination with montelukast showed additive inhibitory effects on eosinophil infiltration and late phase nasal obstruction in a model of allergic rhinitis. Ameliorated airway inflammation in an ovalbumin model of allergic airway inflammation. | [72,73] |
| fevipiprant | DP2 receptor antagonist | Phase III clinical studies did not show a statistically significant reduction in asthma exacerbations after adjusting for multiple testing; however, consistent and modest reductions in exacerbations rates were observed with the 450 mg (higher) dose. | [74] |
| ONO-0453 | DP1 receptor antagonist | Shows improvement in all nasal and eye symptoms in a Phase II clinical study in seasonal allergic rhinitis patients. | [75] |
| butaprost | EP2 receptor agonist | Ameliorates airway inflammation and allergen-induced accumulation of eosinophils in the lungs of ovalbumin-sensitized mice. Limits airway hyperresponsiveness and inflammation in a house dust mite induced model. | [76,77] |
| iloprost | stable analogue of PGI2 | Inhibits eosinophil trafficking and improves cardinal features of asthma in preclinical models | [78,79] |
| S-Y048 | 5-oxo-ETE receptor antagonist | Inhibits eosinophil infiltration in the skin following intradermal allergen challenge in primates. | [80] |
| Bioactive Lipid Mediator | Mode of Action on Eosinophils | References |
|---|---|---|
| 2-arachidonolyglycerol | Induces chemotaxis Induces calcium flux | [68,101,102] [103] |
| 5-oxo-ETE | Increases chemotaxis Increases calcium flux, actin reorganization and ROS production Increases infiltration in the skin following intradermal application Upregulates eosinophil cationic protein in nasal polyps in vitro | [104,105,106] [104] [107] [108] |
| Cysteinyl leukotrienes | Induce chemotaxis and adhesion molecule upregulation Induce eosinophil trans endothelial migration, superoxide production and degranulation Elicit secretion from within cell-free human eosinophil granules | [109] [110] [111] |
| Leukotriene B4 | Induces infiltration to the inflamed peritoneum | [112] |
| Lipoxin A4 | Inhibits eicosanoid-induced tissue migration and airway hyperresponsiveness | [61] |
| Lysophosphatidic Acid | Increases CD11b upregulation, superoxide production, chemotaxis, calcium flux and actin reorganization Increases airway infiltration and superoxide production | [113] [114] |
| Lysophosphatidylcholine | Pro-inflammatory Induces CD11b upregulation, adhesion and calcium flux Increases infiltration in the airways and airway resistance Anti-inflammatory Inhibits CD11b upregulation, calcium flux, Akt and Erk phosphorylation as well as in vitro and in vivo migration | [115] [116] [101] |
| Lysophosphatidylserine | Induces degranulation Induces eosinophil extracellular trap formation | [102] [117] |
| Platelet Activating Factor | Induces calcium flux Increases chemokinesis and chemotaxis Induces degranulation Induces prostanoid secretion | [92] [93] [90] [91] |
| Prostaglandin D2 | DP2 induces calcium flux, CD11b upregulation, migration and degranulation primes eosinophils for other chemoattractants DP1 prolongs survival enhances DP2 signaling | [118,119] [120,121] [118,122] [122,123] |
| Prostaglandin E2 | Inhibits chemotaxis, CD11b upregulation, degranulation, calcium flux, cytoskeletal rearrangement and reactive oxygen production Prolongs their survival Decreases sputum counts | [76,124] [125,126] [127] |
| Prostacyclin | Inhibits migration and adhesion in vitro, reduces CD11b upregulation and activation Inhibits migration in vivo | [128] [79] |
| Protectin D1 | Inhibits chemotaxis as well as CD11b and L-selectin expression Decreases airway infiltration and airway hyperresponsiveness | [129] [60] |
| Resolvin E3 | Reduces infiltrating airway eosinophils in an HDM-induced airway inflammation model | [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knuplez, E.; Sturm, E.M.; Marsche, G. Emerging Role of Phospholipase-Derived Cleavage Products in Regulating Eosinophil Activity: Focus on Lysophospholipids, Polyunsaturated Fatty Acids and Eicosanoids. Int. J. Mol. Sci. 2021, 22, 4356. https://doi.org/10.3390/ijms22094356
Knuplez E, Sturm EM, Marsche G. Emerging Role of Phospholipase-Derived Cleavage Products in Regulating Eosinophil Activity: Focus on Lysophospholipids, Polyunsaturated Fatty Acids and Eicosanoids. International Journal of Molecular Sciences. 2021; 22(9):4356. https://doi.org/10.3390/ijms22094356
Chicago/Turabian StyleKnuplez, Eva, Eva Maria Sturm, and Gunther Marsche. 2021. "Emerging Role of Phospholipase-Derived Cleavage Products in Regulating Eosinophil Activity: Focus on Lysophospholipids, Polyunsaturated Fatty Acids and Eicosanoids" International Journal of Molecular Sciences 22, no. 9: 4356. https://doi.org/10.3390/ijms22094356