
The Emerging Clinical Role of Spermine in Prostate Cancer

 , and
, and
Abstract
:1. Introduction
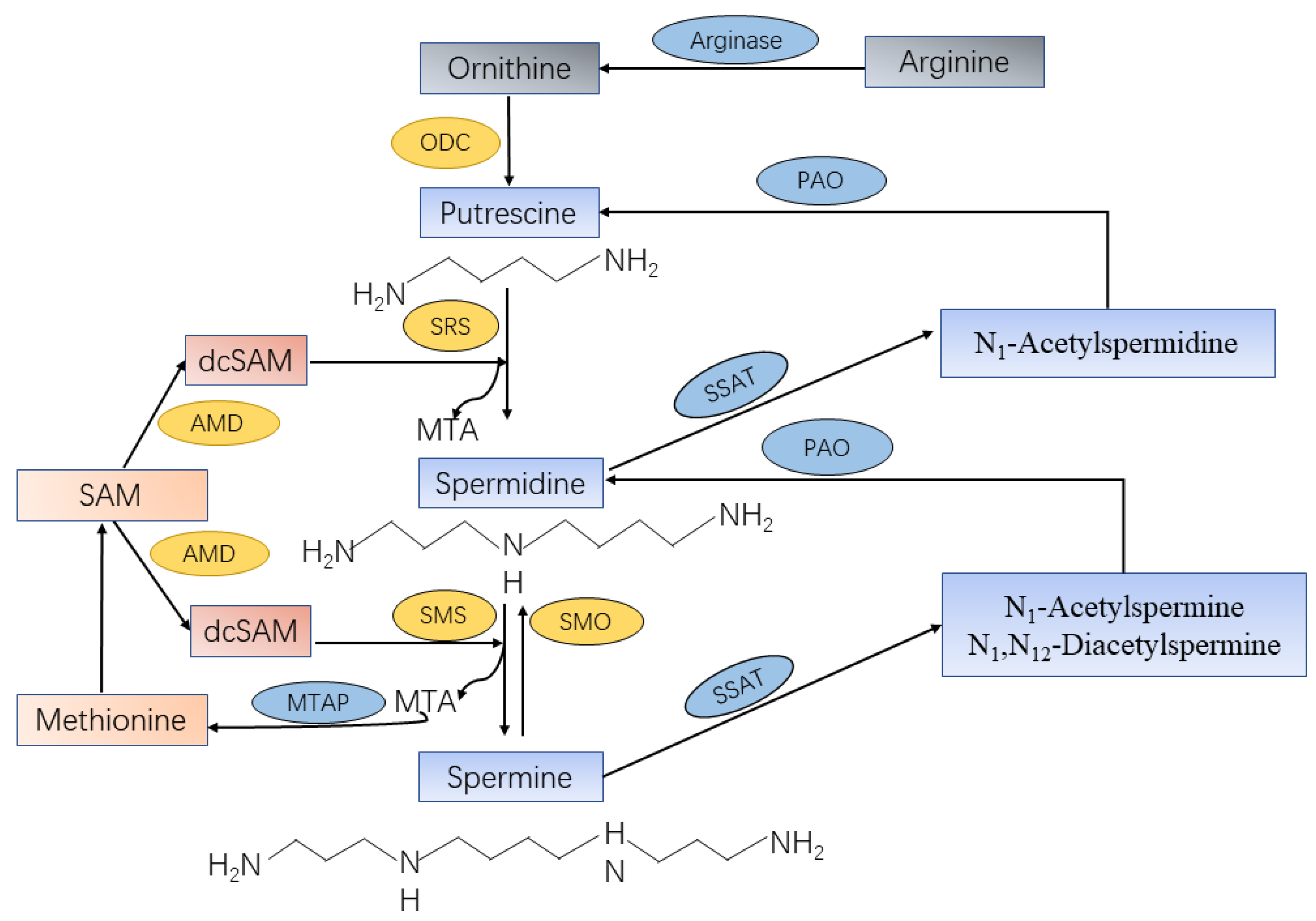
2. The Sources of Spermine
2.1. Extracellular Sources
2.2. Endogenous Biosynthesis
3. Molecular Mechanisms of Spermine for PCa Carcinogenesis and Progression
3.1. Polyamine Metabolism, Oncogenes and Tumor Suppressors
3.2. Polyamine Metabolism in PCa-Associated Oxidative Stress and Inflammation
3.3. Anticancer Immunosurveillance
3.4. Apoptosis
4. Spermine as a Biomarker for PCa
4.1. Urine
4.2. Tissue
4.3. Human Expressed Prostatic Secretions (EPS)
4.4. Erythrocyte
5. Therapeutic Potential for PCa by Targeting Spermine Metabolism Pathway
5.1. Inhibition of Anabolism Pathway
5.2. Catabolism Pathway Activation
5.3. Development and Use of Polyamine Analogues
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gerner, E.W.; Meyskens, F.L., Jr. Polyamines and cancer: Old molecules, new understanding. Nat. Rev. Cancer 2004, 4, 781–792. [Google Scholar] [CrossRef] [Green Version]
- Lima, A.R.; Pinto, J.; Bastos, M.L.; Carvalho, M.; Guedes de Pinho, P. NMR-based metabolomics studies of human prostate cancer tissue. Metabolomics 2018, 14, 88. [Google Scholar] [CrossRef] [PubMed]
- Bachrach, U. The early history of polyamine research. Plant Physiol. Biochem. 2010, 48, 490–495. [Google Scholar] [CrossRef]
- Harrison, G.A. Spermine in human tissues. Biochem. J. 1931, 25, 1885–1892. [Google Scholar] [CrossRef] [PubMed]
- Mi, Z.; Kramer, D.L.; Miller, J.T.; Bergeron, R.J.; Bernacki, R.; Porter, C.W. Human prostatic carcinoma cell lines display altered regulation of polyamine transport in response to polyamine analogs and inhibitors. Prostate 1998, 34, 51–60. [Google Scholar] [CrossRef]
- Litwin, M.S.; Tan, H.J. The Diagnosis and Treatment of Prostate Cancer: A Review. JAMA 2017, 317, 2532–2542. [Google Scholar] [CrossRef] [PubMed]
- van der Graaf, M.; Schipper, R.G.; Oosterhof, G.O.; Schalken, J.A.; Verhofstad, A.A.; Heerschap, A. Proton MR spectroscopy of prostatic tissue focused on the detection of spermine, a possible biomarker of malignant behavior in prostate cancer. MAGMA 2000, 10, 153–159. [Google Scholar] [CrossRef]
- Smith, R.C.; Litwin, M.S.; Lu, Y.; Zetter, B.R. Identification of an endogenous inhibitor of prostatic carcinoma cell growth. Nat. Med. 1995, 1, 1040–1045. [Google Scholar] [CrossRef]
- Koike, C.; Chao, D.T.; Zetter, B.R. Sensitivity to polyamine-induced growth arrest correlates with antizyme induction in prostate carcinoma cells. Cancer Res. 1999, 59, 6109–6112. [Google Scholar]
- Steele, C.B.; Li, J.; Huang, B.; Weir, H.K. Prostate cancer survival in the United States by race and stage (2001-2009): Findings from the CONCORD-2 study. Cancer 2017, 123 (Suppl. 24), 5160–5177. [Google Scholar] [CrossRef]
- Rhodes, D.R.; Barrette, T.R.; Rubin, M.A.; Ghosh, D.; Chinnaiyan, A.M. Meta-analysis of microarrays: Interstudy validation of gene expression profiles reveals pathway dysregulation in prostate cancer. Cancer Res. 2002, 62, 4427–4433. [Google Scholar] [PubMed]
- Cipolla, B.; Guilli, F.; Moulinoux, J.P. Polyamine-reduced diet in metastatic hormone-refractory prostate cancer (HRPC) patients. Biochem. Soc. Trans. 2003, 31, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Kalac, P. Health effects and occurrence of dietary polyamines: A review for the period 2005-mid 2013. Food Chem. 2014, 161, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Milovic, V. Polyamines in the gut lumen: Bioavailability and biodistribution. Eur. J. Gastroenterol. Hepatol. 2001, 13, 1021–1025. [Google Scholar] [CrossRef] [PubMed]
- Ikeguchi, Y.; Bewley, M.C.; Pegg, A.E. Aminopropyltransferases: Function, structure and genetics. J. Biochem. 2006, 139, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Casero, R.A.; Pegg, A.E. Polyamine catabolism and disease. Biochem. J. 2009, 421, 323–338. [Google Scholar] [CrossRef] [Green Version]
- Holst, C.M.; Nevsten, P.; Johansson, F.; Carlemalm, E.; Oredsson, S.M. Subcellular distribution of spermidine/spermine N1-acetyltransferase. Cell Biol. Int. 2008, 32, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Pegg, A.E. Toxicity of polyamines and their metabolic products. Chem. Res. Toxicol. 2013, 26, 1782–1800. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bello-Fernandez, C.; Packham, G.; Cleveland, J.L. The ornithine decarboxylase gene is a transcriptional target of c-Myc. Proc. Natl. Acad. Sci. USA 1993, 90, 7804–7808. [Google Scholar] [CrossRef] [Green Version]
- Origanti, S.; Nowotarski, S.L.; Carr, T.D.; Sass-Kuhn, S.; Xiao, L.; Wang, J.Y.; Shantz, L.M. Ornithine decarboxylase mRNA is stabilized in an mTORC1-dependent manner in Ras-transformed cells. Biochem. J. 2012, 442, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Shantz, L.M. Transcriptional and translational control of ornithine decarboxylase during Ras transformation. Biochem. J. 2004, 377, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Ignatenko, N.A.; Babbar, N.; Mehta, D.; Casero, R.A., Jr.; Gerner, E.W. Suppression of polyamine catabolism by activated Ki-ras in human colon cancer cells. Mol. Carcinog. 2004, 39, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Zabala-Letona, A.; Arruabarrena-Aristorena, A.; Martin-Martin, N.; Fernandez-Ruiz, S.; Sutherland, J.D.; Clasquin, M.; Tomas-Cortazar, J.; Jimenez, J.; Torres, I.; Quang, P.; et al. mTORC1-dependent AMD1 regulation sustains polyamine metabolism in prostate cancer. Nature 2017, 547, 109–113. [Google Scholar] [CrossRef]
- Affronti, H.C.; Rowsam, A.M.; Pellerite, A.J.; Rosario, S.R.; Long, M.D.; Jacobi, J.J.; Bianchi-Smiraglia, A.; Boerlin, C.S.; Gillard, B.M.; Karasik, E.; et al. Pharmacological polyamine catabolism upregulation with methionine salvage pathway inhibition as an effective prostate cancer therapy. Nat. Commun. 2020, 11, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandoth, C.; McLellan, M.D.; Vandin, F.; Ye, K.; Niu, B.; Lu, C.; Xie, M.; Zhang, Q.; McMichael, J.F.; Wyczalkowski, M.A.; et al. Mutational landscape and significance across 12 major cancer types. Nature 2013, 502, 333–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, Y.; Wang, S.J.; Li, D.; Chu, B.; Gu, W. Activation of SAT1 engages polyamine metabolism with p53-mediated ferroptotic responses. Proc. Natl. Acad. Sci. USA 2016, 113, E6806–E6812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiota, M.; Yokomizo, A.; Naito, S. Oxidative stress and androgen receptor signaling in the development and progression of castration-resistant prostate cancer. Free Radic. Biol. Med. 2011, 51, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Bhatia-Gaur, R.; Donjacour, A.A.; Sciavolino, P.J.; Kim, M.; Desai, N.; Young, P.; Norton, C.R.; Gridley, T.; Cardiff, R.D.; Cunha, G.R.; et al. Roles for Nkx3.1 in prostate development and cancer. Genes Dev. 1999, 13, 966–977. [Google Scholar] [CrossRef] [Green Version]
- Tan, P.Y.; Chang, C.W.; Chng, K.R.; Wansa, K.D.; Sung, W.K.; Cheung, E. Integration of regulatory networks by NKX3-1 promotes androgen-dependent prostate cancer survival. Mol. Cell Biol. 2012, 32, 399–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehraein-Ghomi, F.; Basu, H.S.; Church, D.R.; Hoffmann, F.M.; Wilding, G. Androgen receptor requires JunD as a coactivator to switch on an oxidative stress generation pathway in prostate cancer cells. Cancer Res. 2010, 70, 4560–4568. [Google Scholar] [CrossRef] [Green Version]
- Basu, H.S.; Thompson, T.A.; Church, D.R.; Clower, C.C.; Mehraein-Ghomi, F.; Amlong, C.A.; Martin, C.T.; Woster, P.M.; Lindstrom, M.J.; Wilding, G. A small molecule polyamine oxidase inhibitor blocks androgen-induced oxidative stress and delays prostate cancer progression in the transgenic adenocarcinoma of the mouse prostate model. Cancer Res. 2009, 69, 7689–7695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pledgie, A.; Huang, Y.; Hacker, A.; Zhang, Z.; Woster, P.M.; Davidson, N.E.; Casero, R.A., Jr. Spermine oxidase SMO(PAOh1), Not N1-acetylpolyamine oxidase PAO, is the primary source of cytotoxic H2O2 in polyamine analogue-treated human breast cancer cell lines. J. Biol. Chem. 2005, 280, 39843–39851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodwin, A.C.; Jadallah, S.; Toubaji, A.; Lecksell, K.; Hicks, J.L.; Kowalski, J.; Bova, G.S.; De Marzo, A.M.; Netto, G.J.; Casero, R.A., Jr. Increased spermine oxidase expression in human prostate cancer and prostatic intraepithelial neoplasia tissues. Prostate 2008, 68, 766–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.W.; Nelson, P.S. The role of cyclooxygenase-2 inhibition for the prevention and treatment of prostate carcinoma. Clin. Prostate Cancer 2003, 2, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cameron, G.A.; Wallace, H.M. Decreased sensitivity to aspirin is associated with altered polyamine metabolism in human prostate cancer cells. Amino Acids 2016, 48, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Hayes, C.S.; Defeo, K.; Dang, H.; Trempus, C.S.; Morris, R.J.; Gilmour, S.K. A prolonged and exaggerated wound response with elevated ODC activity mimics early tumor development. Carcinogenesis 2011, 32, 1340–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, C.S.; Shicora, A.C.; Keough, M.P.; Snook, A.E.; Burns, M.R.; Gilmour, S.K. Polyamine-blocking therapy reverses immunosuppression in the tumor microenvironment. Cancer Immunol. Res. 2014, 2, 274–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, E.T.; Minton, A.; Peters, M.C.; Phanstiel, O.T.; Gilmour, S.K. A novel polyamine blockade therapy activates an anti-tumor immune response. Oncotarget 2017, 8, 84140–84152. [Google Scholar] [CrossRef] [Green Version]
- Soda, K. The mechanisms by which polyamines accelerate tumor spread. J. Exp. Clin. Cancer Res. 2011, 30, 95. [Google Scholar] [CrossRef] [Green Version]
- Southan, G.J.; Szabo, C.; Thiemermann, C. Inhibition of the induction of nitric oxide synthase by spermine is modulated by aldehyde dehydrogenase. Biochem. Biophys. Res. Commun. 1994, 203, 1638–1644. [Google Scholar] [CrossRef]
- Zhang, M.; Caragine, T.; Wang, H.; Cohen, P.S.; Botchkina, G.; Soda, K.; Bianchi, M.; Ulrich, P.; Cerami, A.; Sherry, B.; et al. Spermine inhibits proinflammatory cytokine synthesis in human mononuclear cells: A counterregulatory mechanism that restrains the immune response. J. Exp. Med. 1997, 185, 1759–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Wang, H.; Tracey, K.J. Regulation of macrophage activation and inflammation by spermine: A new chapter in an old story. Crit. Care Med. 2000, 28, N60–N66. [Google Scholar] [CrossRef] [PubMed]
- Quan, C.P.; Roux, C.; Pillot, J.; Bouvet, J.P. Delineation between T and B suppressive molecules from human seminal plasma: II. Spermine is the major suppressor of T-lymphocytes in vitro. Am. J. Reprod. Immunol. 1990, 22, 64–69. [Google Scholar] [CrossRef]
- Singh, K.; Coburn, L.A.; Asim, M.; Barry, D.P.; Allaman, M.M.; Shi, C.; Washington, M.K.; Luis, P.B.; Schneider, C.; Delgado, A.G.; et al. Ornithine Decarboxylase in Macrophages Exacerbates Colitis and Promotes Colitis-Associated Colon Carcinogenesis by Impairing M1 Immune Responses. Cancer Res. 2018, 78, 4303–4315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegg, A.E.; Lockwood, D.H.; Williams-Ashman, H.G. Concentrations of putrescine and polyamines and their enzymic synthesis during androgen-induced prostatic growth. Biochem. J. 1970, 117, 17–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arisan, E.D.; Obakan, P.; Coker-Gurkan, A.; Calcabrini, A.; Agostinelli, E.; Unsal, N.P. CDK inhibitors induce mitochondria-mediated apoptosis through the activation of polyamine catabolic pathway in LNCaP, DU145 and PC3 prostate cancer cells. Curr. Pharm. Des. 2014, 20, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Schipper, R.G.; Deli, G.; Deloyer, P.; Lange, W.P.; Schalken, J.A.; Verhofstad, A.A. Antitumor activity of the polyamine analog N(1), N(11)-diethylnorspermine against human prostate carcinoma cells. Prostate 2000, 44, 313–321. [Google Scholar] [CrossRef]
- Schipper, R.G.; Penning, L.C.; Verhofstad, A.A. Involvement of polyamines in apoptosis. Facts and controversies: Effectors or protectors? Semin. Cancer Biol. 2000, 10, 55–68. [Google Scholar] [CrossRef]
- Eiseman, J.L.; Rogers, F.A.; Guo, Y.; Kauffman, J.; Sentz, D.L.; Klinger, M.F.; Callery, P.S.; Kyprianou, N. Tumor-targeted apoptosis by a novel spermine analogue, 1,12-diaziridinyl-4,9-diazadodecane, results in therapeutic efficacy and enhanced radiosensitivity of human prostate cancer. Cancer Res. 1998, 58, 4864–4870. [Google Scholar]
- Redman, C.; Xu, M.J.; Peng, Y.M.; Scott, J.A.; Payne, C.; Clark, L.C.; Nelson, M.A. Involvement of polyamines in selenomethionine induced apoptosis and mitotic alterations in human tumor cells. Carcinogenesis 1997, 18, 1195–1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, J.M.; Carson, D.A. Ca2+/Mg(2+)-dependent endonuclease from human spleen: Purification, properties, and role in apoptosis. Biochemistry 1993, 32, 9129–9136. [Google Scholar] [CrossRef] [PubMed]
- Basu, H.S.; Smirnov, I.V.; Peng, H.F.; Tiffany, K.; Jackson, V. Effects of spermine and its cytotoxic analogs on nucleosome formation on topologically stressed DNA in vitro. Eur. J. Biochem. 1997, 243, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.C.; Sirisoma, N.S.; Kuppusamy, P.; Zweier, J.L.; Woster, P.M.; Casero, R.A., Jr. The natural polyamine spermine functions directly as a free radical scavenger. Proc. Natl. Acad. Sci. USA 1998, 95, 11140–11145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanford, E.J.; Drago, J.R.; Rohner, T.J.; Kessler, G.F.; Sheehan, L.; Lipton, A. Preliminary evaluation of urinary polyamines in the diagnosis of genitourinary tract malignancy. J. Urol. 1975, 113, 218–221. [Google Scholar] [CrossRef]
- Fair, W.R.; Wehner, N.; Brorsson, U. Urinary polyamine levels in the diagnosis of carcinoma of the prostate. J. Urol. 1975, 114, 88–92. [Google Scholar] [CrossRef]
- Sugimoto, M.; Hiramatsu, K.; Kamei, S.; Kinoshita, K.; Hoshino, M.; Iwasaki, K.; Kawakita, M. Significance of urinary N1,N8-diacetylspermidine and N1,N12-diacetylspermine as indicators of neoplastic diseases. J. Cancer Res. Clin. Oncol. 1995, 121, 317–319. [Google Scholar] [CrossRef]
- Hiramatsu, K.; Sugimoto, M.; Kamei, S.; Hoshino, M.; Kinoshita, K.; Iwasaki, K.; Kawakita, M. Diagnostic and prognostic usefulness of N1,N8-diacetylspermidine and N1,N12-diacetylspermine in urine as novel markers of malignancy. J. Cancer Res. Clin. Oncol. 1997, 123, 539–545. [Google Scholar] [CrossRef]
- Tsoi, T.H.; Chan, C.F.; Chan, W.L.; Chiu, K.F.; Wong, W.T.; Ng, C.F.; Wong, K.L. Urinary Polyamines: A Pilot Study on Their Roles as Prostate Cancer Detection Biomarkers. PLoS ONE 2016, 11, e0162217. [Google Scholar] [CrossRef] [Green Version]
- Chiu, P.K.; Fung, Y.H.; Teoh, J.Y.; Chan, C.H.; Lo, K.L.; Li, K.M.; Tse, R.T.; Leung, C.H.; Wong, Y.P.; Roobol, M.J.; et al. Urine spermine and multivariable Spermine Risk Score predict high-grade prostate cancer. Prostate Cancer Prostatic Dis. 2021. [Google Scholar] [CrossRef]
- Shukla-Dave, A.; Castillo-Martin, M.; Chen, M.; Lobo, J.; Gladoun, N.; Collazo-Lorduy, A.; Khan, F.M.; Ponomarev, V.; Yi, Z.; Zhang, W.; et al. Ornithine Decarboxylase Is Sufficient for Prostate Tumorigenesis via Androgen Receptor Signaling. Am. J. Pathol. 2016, 186, 3131–3145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schipper, R.G.; Romijn, J.C.; Cuijpers, V.M.; Verhofstad, A.A. Polyamines and prostatic cancer. Biochem. Soc. Trans. 2003, 31, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Swanson, M.G.; Vigneron, D.B.; Tabatabai, Z.L.; Males, R.G.; Schmitt, L.; Carroll, P.R.; James, J.K.; Hurd, R.E.; Kurhanewicz, J. Proton HR-MAS spectroscopy and quantitative pathologic analysis of MRI/3D-MRSI-targeted postsurgical prostate tissues. Magn. Reson. Med. 2003, 50, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Giskeodegard, G.F.; Bertilsson, H.; Selnaes, K.M.; Wright, A.J.; Bathen, T.F.; Viset, T.; Halgunset, J.; Angelsen, A.; Gribbestad, I.S.; Tessem, M.B. Spermine and citrate as metabolic biomarkers for assessing prostate cancer aggressiveness. PLoS ONE 2013, 8, e62375. [Google Scholar] [CrossRef] [Green Version]
- Swanson, M.G.; Zektzer, A.S.; Tabatabai, Z.L.; Simko, J.; Jarso, S.; Keshari, K.R.; Schmitt, L.; Carroll, P.R.; Shinohara, K.; Vigneron, D.B.; et al. Quantitative analysis of prostate metabolites using 1H HR-MAS spectroscopy. Magn. Reson. Med. 2006, 55, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Basharat, M.; Payne, G.S.; Morgan, V.A.; Parker, C.; Dearnaley, D.; deSouza, N.M. TE = 32 ms vs TE = 100 ms echo-time (1)H-magnetic resonance spectroscopy in prostate cancer: Tumor metabolite depiction and absolute concentrations in tumors and adjacent tissues. J. Magn. Reson. Imaging 2015, 42, 1086–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxeiner, A.; Adkins, C.B.; Zhang, Y.; Taupitz, M.; Halpern, E.F.; McDougal, W.S.; Wu, C.L.; Cheng, L.L. Retrospective analysis of prostate cancer recurrence potential with tissue metabolomic profiles. Prostate 2010, 70, 710–717. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Martin, M.L.; Adrados, M.; Ortega, M.P.; Fernandez Gonzalez, I.; Lopez-Larrubia, P.; Viano, J.; Garcia-Segura, J.M. Quantitative (1) H MR spectroscopic imaging of the prostate gland using LCModel and a dedicated basis-set: Correlation with histologic findings. Magn. Reson. Med. 2011, 65, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, R.; Gomez, A.M.; Raman, S.S.; Margolis, D.J.; McClure, T.; Thomas, M.A. Correlation of endorectal 2D JPRESS findings with pathological Gleason scores in prostate cancer patients. NMR Biomed. 2010, 23, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Kobus, T.; Wright, A.J.; Weiland, E.; Heerschap, A.; Scheenen, T.W. Metabolite ratios in 1H MR spectroscopic imaging of the prostate. Magn. Reson. Med. 2015, 73, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Selnaes, K.M.; Gribbestad, I.S.; Bertilsson, H.; Wright, A.; Angelsen, A.; Heerschap, A.; Tessem, M.B. Spatially matched in vivo and ex vivo MR metabolic profiles of prostate cancer—Investigation of a correlation with Gleason score. NMR Biomed. 2013, 26, 600–606. [Google Scholar] [CrossRef]
- Klomp, D.W.; Bitz, A.K.; Heerschap, A.; Scheenen, T.W. Proton spectroscopic imaging of the human prostate at 7 T. NMR Biomed. 2009, 22, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wang, Y.; Xiao, Z.G.; Zou, C.; Zhang, X.; Wang, Z.; Wu, D.; Yu, S.; Chan, F.L. Nuclear receptor ERRalpha and transcription factor ERG form a reciprocal loop in the regulation of TMPRSS2:ERG fusion gene in prostate cancer. Oncogene 2018, 37, 6259–6274. [Google Scholar] [CrossRef]
- Hansen, A.F.; Sandsmark, E.; Rye, M.B.; Wright, A.J.; Bertilsson, H.; Richardsen, E.; Viset, T.; Bofin, A.M.; Angelsen, A.; Selnaes, K.M.; et al. Presence of TMPRSS2-ERG is associated with alterations of the metabolic profile in human prostate cancer. Oncotarget 2016, 7, 42071–42085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braadland, P.R.; Giskeodegard, G.; Sandsmark, E.; Bertilsson, H.; Euceda, L.R.; Hansen, A.F.; Guldvik, I.J.; Selnaes, K.M.; Grytli, H.H.; Katz, B.; et al. Ex vivo metabolic fingerprinting identifies biomarkers predictive of prostate cancer recurrence following radical prostatectomy. Br. J. Cancer 2017, 117, 1656–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, R.J.; McNeal, J.E.; Edgar, S.G.; Robertson, T.; Dawkins, H.J. Characterization of cytoplasmic secretory granules (PSG), in prostatic epithelium and their transformation-induced loss in dysplasia and adenocarcinoma. Hum. Pathol. 1998, 29, 1488–1494. [Google Scholar] [CrossRef]
- Cohen, R.J.; Fujiwara, K.; Holland, J.W.; McNeal, J.E. Polyamines in prostatic epithelial cells and adenocarcinoma; the effects of androgen blockade. Prostate 2001, 49, 278–284. [Google Scholar] [CrossRef]
- Lima, A.R.; Bastos Mde, L.; Carvalho, M.; Guedes de Pinho, P. Biomarker Discovery in Human Prostate Cancer: An Update in Metabolomics Studies. Transl. Oncol. 2016, 9, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.J.; Nicholson, J.K. Proton MRS of human prostatic fluid: Correlations between citrate, spermine, and myo-inositol levels and changes with disease. Prostate 1997, 30, 248–255. [Google Scholar] [CrossRef]
- Serkova, N.J.; Gamito, E.J.; Jones, R.H.; O'Donnell, C.; Brown, J.L.; Green, S.; Sullivan, H.; Hedlund, T.; Crawford, E.D. The metabolites citrate, myo-inositol, and spermine are potential age-independent markers of prostate cancer in human expressed prostatic secretions. Prostate 2008, 68, 620–628. [Google Scholar] [CrossRef]
- Moulinoux, J.P.; Le Calve, M.; Quemener, V.; Quash, G. In vitro studies on the entry of polyamines into normal red blood cells. Biochimie 1984, 66, 385–393. [Google Scholar] [CrossRef]
- Cipolla, B.; Moulinoux, J.P.; Quemener, V.; Havouis, R.; Martin, L.A.; Guille, F.; Lobel, B. Erythrocyte polyamine levels in human prostatic carcinoma. J. Urol. 1990, 144, 1164–1166. [Google Scholar] [CrossRef]
- Cipolla, B.; Guille, F.; Moulinoux, J.P.; Quemener, V.; Staerman, F.; Corbel, L.; Lobel, B. Polyamines and prostatic carcinoma: Clinical and therapeutic implications. Eur. Urol. 1993, 24, 124–131. [Google Scholar] [CrossRef]
- Cipolla, B.; Guille, F.; Moulinoux, J.P.; Bansard, J.Y.; Roth, S.; Staerman, F.; Corbel, L.; Quemener, V.; Lobel, B. Erythrocyte polyamines and prognosis in stage D2 prostatic carcinoma patients. J. Urol. 1994, 151, 629–633. [Google Scholar] [CrossRef]
- Cipolla, B.G.; Ziade, J.; Bansard, J.Y.; Moulinoux, J.P.; Staerman, F.; Quemener, V.; Lobel, B.; Guille, F. Pretherapeutic erythrocyte polyamine spermine levels discriminate high risk relapsing patients with M1 prostate carcinoma. Cancer 1996, 78, 1055–1065. [Google Scholar] [CrossRef]
- Martinez-Outschoorn, U.E.; Peiris-Pages, M.; Pestell, R.G.; Sotgia, F.; Lisanti, M.P. Cancer metabolism: A therapeutic perspective. Nat. Rev. Clin. Oncol. 2017, 14, 11–31. [Google Scholar] [CrossRef]
- Bettuzzi, S.; Davalli, P.; Astancolle, S.; Carani, C.; Madeo, B.; Tampieri, A.; Corti, A. Tumor progression is accompanied by significant changes in the levels of expression of polyamine metabolism regulatory genes and clusterin (sulfated glycoprotein 2) in human prostate cancer specimens. Cancer Res. 2000, 60, 28–34. [Google Scholar]
- Dunzendorfer, U.; Relyea, N.M.; Kleinert, E.; Balis, M.E.; Whitmore, W.F., Jr. Antigrowth effect of some inhibitors of polyamine synthesis on transplantable prostate cancer. Oncology 1983, 40, 57–62. [Google Scholar] [CrossRef]
- Herr, H.W.; Kleinert, E.L.; Relyea, N.M.; Whitmore, W.F., Jr. Potentiation of methylglyoxal-bis-guanylhydrazone by alpha-difluoromethylornithine in rat prostate cancer. Cancer 1984, 53, 1294–1298. [Google Scholar] [CrossRef]
- Delworth, M.; Nishioka, K.; Pettaway, C.; Gutman, M.; Killion, J.; Voneschenbach, A.; Fidler, I. Systemic administration of 4-amidinoindanon-1-(2′-amidino)-hydrazone, a new inhibitor of s-adenosylmethionine decarboxylase, produces cytostasis of human prostate-cancer in athymic nude-mice. Int. J. Oncol. 1995, 6, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Heston, W.D.; Kadmon, D.; Lazan, D.W.; Fair, W.R. Copenhagen rat prostatic tumor ornithine decarboxylase activity (ODC) and the effect of the ODC inhibitor alpha-difluoromethylornithine. The Prostate 1982, 3, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Kadmon, D. Chemoprevention in prostate cancer: The role of difluoromethylornithine (DFMO). J. Cell. Biochem. 1992, 50 (Suppl. 16H), 122–127. [Google Scholar] [CrossRef]
- Devens, B.H.; Weeks, R.S.; Burns, M.R.; Carlson, C.L.; Brawer, M.K. Polyamine depletion therapy in prostate cancer. Prostate Cancer Prostatic Dis. 2000, 3, 275–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herr, H.W.; Warrel, R.P.; Burchenal, J.H. Phase I trial of alpha-difluoromethyl ornithine (DFMO) and methylglyoxal bis (guanylhydrazone) (MGBG) in patients with advanced prostatic cancer. Urology 1986, 28, 508–511. [Google Scholar] [CrossRef]
- Horn, Y.; Schechter, P.J.; Marton, L.J. Phase I-II clinical trial with alpha-difluoromethylornithine--an inhibitor of polyamine biosynthesis. Eur. J. Cancer Clin. Oncol. 1987, 23, 1103–1107. [Google Scholar] [CrossRef]
- Carbone, P.P.; Douglas, J.A.; Larson, P.O.; Verma, A.K.; Blair, I.A.; Pomplun, M.; Tutsch, K.D. Phase I chemoprevention study of piroxicam and alpha-difluoromethylornithine. Cancer Epidemiol. Biomark. Prev. 1998, 7, 907–912. [Google Scholar]
- Wallace, H.M.; Niiranen, K. Polyamine analogues—An update. Amino Acids 2007, 33, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Ahmad, N.; Marengo, S.R.; MacLennan, G.T.; Greenberg, N.M.; Mukhtar, H. Chemoprevention of prostate carcinogenesis by alpha-difluoromethylornithine in TRAMP mice. Cancer Res. 2000, 60, 5125–5133. [Google Scholar] [PubMed]
- Simoneau, A.R.; Gerner, E.W.; Nagle, R.; Ziogas, A.; Fujikawa-Brooks, S.; Yerushalmi, H.; Ahlering, T.E.; Lieberman, R.; McLaren, C.E.; Anton-Culver, H.; et al. The effect of difluoromethylornithine on decreasing prostate size and polyamines in men: Results of a year-long phase IIb randomized placebo-controlled chemoprevention trial. Cancer Epidemiol. Biomark. Prev. 2008, 17, 292–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kee, K.; Vujcic, S.; Merali, S.; Diegelman, P.; Kisiel, N.; Powell, C.T.; Kramer, D.L.; Porter, C.W. Metabolic and antiproliferative consequences of activated polyamine catabolism in LNCaP prostate carcinoma cells. J. Biol. Chem. 2004, 279, 27050–27058. [Google Scholar] [CrossRef] [Green Version]
- Kee, K.; Foster, B.A.; Merali, S.; Kramer, D.L.; Hensen, M.L.; Diegelman, P.; Kisiel, N.; Vujcic, S.; Mazurchuk, R.V.; Porter, C.W. Activated polyamine catabolism depletes acetyl-CoA pools and suppresses prostate tumor growth in TRAMP mice. J. Biol. Chem. 2004, 279, 40076–40083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffers, L.; Church, D.; Basu, H.; Marton, L.; Wilding, G. Effects of the polyamine analogues BE-4-4-4-4, BE-3-7-3, and BE-3-3-3 on the proliferation of three prostate cancer cell lines. Cancer Chemother. Pharmacol. 1997, 40, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Zagaja, G.P.; Shrivastav, M.; Fleig, M.J.; Marton, L.J.; Rinker-Schaeffer, C.W.; Dolan, M.E. Effects of polyamine analogues on prostatic adenocarcinoma cells in vitro and in vivo. Cancer Chemother. Pharmacol. 1998, 41, 505–512. [Google Scholar] [CrossRef] [PubMed]
- McCloskey, D.E.; Woster, P.M.; Casero, R.A., Jr.; Davidson, N.E. Effects of the polyamine analogues N1-ethyl-N11-((cyclopropyl)methyl)-4,8-diazaundecane and N1-ethylN-11-((cycloheptyl)methyl)-4,8-diazaundecane in human prostate cancer cells. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2000, 6, 17–23. [Google Scholar]
- Valasinas, A.; Sarkar, A.; Reddy, V.K.; Marton, L.J.; Basu, H.S.; Frydman, B. Conformationally restricted analogues of 1N,14N-bisethylhomospermine (BE-4-4-4): Synthesis and growth inhibitory effects on human prostate cancer cells. J. Med. Chem. 2001, 44, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.K.; Sarkar, A.; Valasinas, A.; Marton, L.J.; Basu, H.S.; Frydman, B. cis-Unsaturated analogues of 3,8,13,18,23-pentaazapentacosane (BE-4-4-4-4): Synthesis and growth inhibitory effects on human prostate cancer cell lines. J. Med. Chem. 2001, 44, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Frydman, B.; Bhattacharya, S.; Sarkar, A.; Drandarov, K.; Chesnov, S.; Guggisberg, A.; Popaj, K.; Sergeyev, S.; Yurdakul, A.; Hesse, M.; et al. Macrocyclic polyamines deplete cellular ATP levels and inhibit cell growth in human prostate cancer cells. J. Med. Chem. 2004, 47, 1051–1059. [Google Scholar] [CrossRef]
- Valasinas, A.; Reddy, V.K.; Blokhin, A.V.; Basu, H.S.; Bhattacharya, S.; Sarkar, A.; Marton, L.J.; Frydman, B. Long-chain polyamines (oligoamines) exhibit strong cytotoxicities against human prostate cancer cells. Bioorg. Med. Chem. 2003, 11, 4121–4131. [Google Scholar] [CrossRef]
- Frydman, B.; Blokhin, A.V.; Brummel, S.; Wilding, G.; Maxuitenko, Y.; Sarkar, A.; Bhattacharya, S.; Church, D.; Reddy, V.K.; Kink, J.A.; et al. Cyclopropane-containing polyamine analogues are efficient growth inhibitors of a human prostate tumor xenograft in nude mice. J. Med. Chem. 2003, 46, 4586–4600. [Google Scholar] [CrossRef]

{kind=link}
| Publication | Sample Type | Analytical Platform | PCa Group | Control Group | Main Results | Statistical Significance | Ref. |
|---|---|---|---|---|---|---|---|
| Sanford et al., 1975 | 24-h urine | Beckman spectrophot-ometer | n = 15 PCa | n = 42 healthy controls | ↑Polyamines in 11/15 PCa | NR | [56] |
| Fair et al., 1975 | 12-h/ 24-h urine | Spectronic 20 colorimeter | n = 44 PCa | n = 13 healthy controls | Similarly low levels of spermine detected in cancer and healthy controls | NR | [57] |
| Sugimoto et al., 1995 | Morning Urine | HPLC | n = 24 urogenital cancer, including 13 PCa | n = 43 benign urogenital disorders; n = 52 healthy controls | ↑DiAcSpm in urogenital cancer | NR | [58] |
| Hiramatsu et al., 1997 | Morning Urine | HPLC | n = 31 urogenital cancer, including 15 PCa | n = 42 benign urogenital disorders; n = 52 healthy controls | ↑DiAcSpm in urogenital cancer ↑DiAcSpm in cancer patients with poor prognosis and recurrence | NR | [59] |
| Tsoi et al., 2016 | Pre-biopsy urine with serum PSA level >4.0 ng/mL | UPLC–MS/MS | n = 66 PCa | n = 88 BPH, n = 11 healthy controls | ↓Spermine in PCa; AUC of spermine for PCa: 0.83 | p < 0.0001 | [60] |
| Chiu et al., 2021 | Pre-biopsy urine with serum PSA level 4–20 ng/mL | UPLC–MS/MS | n = 185 PCa; n = 103 HGPCa | n = 415 healthy controls | ↓Spermine in PCa and HGPCa; AUC of spermine risk score: PCa 0.78, HGPCa 0.82 | p < 0.001 | [61] |
| Graaf et al., 2000 | Tissue | HPLC | n = 7 PCa | n = 4 healthy controls, n = 3 BPH | ↓Spermine in PCa | p < 0.05 | [7] |
| Swanson et al., 2003 | Tissue | HRMAS | n = 7 PCa (gland percentage < 20) n = 13 PCa (gland percentage ≥ 20, 8 with GS ≤ 6, 5 with GS ≥ 7) | n = 33 healthy controls | ↓Spermine in PCa compared with controls ↓Spermine in PCa with higher GS | p = 0.01 p = 0.05 | [64] |
| Swanson et al., 2006 | Tissue | HRMAS | n = 60 PCa | n = 6 healthy controls | ↓Spermine in PCa | p < 0.01 | [66] |
| Maxeiner et al., 2010 | Tissue | HRMAS | n = 16 PCa with BCR | n = 32 PCa without BCR (16 clinical-stage-matched and 16 pathological-stage-matched) | Spermine alteration predicts PCa recurrence | NR | [68] |
| Nagarajan et al., 2010 | Tissue | (2D) J-resolved spectroscopy (JPRESS) | n = 7 PCa with GS = 4 + 3 | n = 7 PCa with GS = 3 + 4 | ↑(Cho + Cr)/Spm ratio in PCa with GS = 4 + 3 | p = 0.07 | [70] |
| García-Martín et al., 2011 | Tissue | 1H-MRS | n = 30 | n = 249 | ↑Cho/(Cit + Spm) ratio in PCa | p < 0.001 | [69] |
| Giskeodegar-d et al., 2013 | Tissue | HRMAS | n = 30 PCa with GS = 6; n = 81 HGPCa with GS ≥ 7 | n = 47 normal adjacent samples | ↓Spermine in PCa and HGPCa compared with normal ↓Spermine in HGPCa compared with PCa ↑(Cho+Spm+Cr/Cit) ratio in HGPCa | p = 0.022 p = 0.0044 p = 2.17 × 10-4 | [65] |
| Selnaes et al., 2013 | Tissue | In vivo MRSI and ex vivo HRMAS | n = 15 PCa with GS ≥ 4 + 3 for ex vivo HRMAS n = 19 PCa with GS ≥ 4 + 3 for in vivo MRSI | n = 16 PCa with GS ≤ 3 + 4 for ex vivo HRMASn = 12 PCa with (GS ≤ 3 + 4) for in vivo MRSI | ↑(Cho+Spm+Cr/Cit) ratio with increasing GS | p = 0.035 (ex vivo) p = 0.001 (in vivo) | [72] |
| Basharat et al., 2015 | Tissue | HRMAS | n = 8 PCa with T3 stage n = 19 PCa with GS = 7 | n = 7 PCa with T1 stage, n = 11 with T2 stagen = 6 PCa with GS = 6 | ↓Spermine in PCa with advanced stage and higher GS | T3 vs. T1 p = 0.04 T3 vs. T2 p = 0.08 GS = 7 vs. GS = 6 p = 0.01 | [67] |
| Hansen et al., 2016 | Tissue | HRMAS | n = 34 ERGhigh PCa | n = 30 ERGlow PCa | ↓Spermine in ERGhigh PCa compared with ERGlow PCa | p < 0.001 | [75] |
| Shukla-Dave et al., 2016 | Tissue | Immunofluo-rescence | n = 18 HGPIN; n = 120 PCa | n = 103 healthy controls | ↓Spermine in HGPIN and PCa | p < 0.0001 | [62] |
| Braadland et al., 2017 | Tissue | HRMAS | n = 50 PCa with recurrence | n = 60 PCa without recurrence | ↑Spermine independently associated with better RFS ↑(Cho+Cr)/Spm independently associated with worse RFS | RFS: HR = 0.72, p = 0.016 RFS: HR = 1.43, p = 0.014 | [76] |
| Lynch et al., 1997 | EPS by prostatic massage | 1H-MRS | n = 4 PCa | n = 12 healthy controls; n = 10 BPH; n = 11 vasal aplasia, n = 1 prostatodynia | ↓(Cit to Spm) ratio in PCa | p < 0.02 | [80] |
| Serkova et al., 2008 | EPS by prostatic massage | 1H-MRS | n = 52 PCa | n = 26 healthy controls | ↓Spermine in PCa | p < 0.002 | [81] |
| Cipolla et al., 1990 | Erythrocyte spermine | HPLC | n = 36 PCa with metastases; n = 12 PCa with hormonal escape | n = 17 PCa without metastases; n = 41 PCa with hormonal responsiveness | ↑Spermine in PCa with metastases ↑Spermine in hormone-refractory PCa | p < 0.01 p < 0.001 | [83] |
| Cipolla et al., 1993 | Erythrocyte spermine | HPLC | n = 28 endocrine-treated PCa with progression | n = 23 endocrine-treated PCa without progression | ↑Pretherapeutic spermine level in PCa with progression | p < 0.01 | [84] |
| Cipolla et al., 1994 | Erythrocyte spermine | HPLC | n = 40 newly diagnosed, stage D2 PCa | NA | ↑Spermine associated with shorter PFS and CSS in PCa | PFS: p = 0.001 CSS: p = 0.0025 | [85] |
| Cipolla et al., 1996 | Erythrocyte spermine | HPLC | n = 88 PCa with metastases | NA | ↑Pretherapeutic spermine level predicts worse PFS and CSS in metastatic PCa | PFS: p < 0.0001 CSS: p < 0.0005 | [86] |
| Publication | Inhibitor | Target | PCa Cell Lines | PCa Animal Models | Clinical Trials | Main Results | Ref. | |
|---|---|---|---|---|---|---|---|---|
| Polyamine Synthesis Inhibitor | Heston et al., 1982 | DFMO | ODC | R3327 MAT-Lua | Rat injected with R3327 MAT-Lu | NR | In vitro and in vivo: Inhibition of R3327MAT-Lu growth | [92] |
| Dunzendorfer et al., 1983 | DFMO/MGBG/DFMO + MGBG | ODC + AMD | NR | Rat injected with R3327-Gb | NR | In vivo: Inhibition of R3327-G growth by either DFMO or MGBG; their combination was more effective | [89] | |
| Herr et al., 1984 | DFMO/MGBG/DFMO + MGBG | ODC/AMD/ ODC + AMD | NR | Rat injected with R3327-G | NR | In vivo: DFMO had no antitumor effect, MGBG retarded tumor growth; their combination inhibited tumor growth | [90] | |
| Herr et al., 1986 | DFMO + MGBG | ODC + AMD | NR | NR | Phase I; 5 advanced, hormone-resistant PCa patients | Clinical trial: No antitumor effects, but drugs were well tolerated | [95] | |
| Horn et al., 1987 | DFMO ± doxorubicin + cyclophosphamide | ODC ± conventional chemotherapy | NR | NR | Phase I-II; 9 PCa patients (DFMO + conventional chemotherapy); 5 PCa patients (conventional chemotherapy) | Clinical trial: No effect | [96] | |
| Kadmon et al., 1992 | DFMO | ODC | NR | Rat injected with R3327 MAT-Lu | NR | In vivo: Modest inhibition of R3327 MAT-Lu growth | [93] | |
| Delworth et al., 1995 | CGP-48664 | AMD | LNCaP, LNCaP-LN3, PC-3M, and PC-3M-MM2 | Nude mice injected with LNCaP-LN3 cells or PC-3M-MM2 cells | NR | In vitro and in vivo: Induction of cytostasis; Inhibition of tumor growth in slow-growing tumor, but not fast-growing tumor | [91] | |
| Mi et al., 1998 | CGP-48664 | AMD | LNCaP, DU145, and PC-3 | NR | NR | In vitro: Inhibition of PCa cells growth | [5] | |
| Carbone et al., 1998 | DFMO/PXM/ DFMO + PXM | ODC/ prostaglandin Inhibitor/ODC + prostaglandin Inhibitor | NR | NR | Phase I; 31 cancer patients, including stage A or B PCa | Clinical trial: Dosage toxicity assessment | [97] | |
| Gupta et al., 2000 | DFMO | ODC | NR | TRAMP | NR | In vivo: DFMO prevent prostate tumorigenesis | [99] | |
| Devens et al., 2000 | DFMO | ODC | PC3, LNCaP, DU145 | Nude mice injected with PC-3 cells | NR | In vitro and in vivo: Inhibition of tumor growth | [94] | |
| Simoneau et al., 2008 | DFMO/ placebo | ODC | NR | NR | 81 men with PCa family history but without personal PCa history | Clinical trial: Induction of slow prostate growth and no grade 3 or 4 toxicities | [100] | |
| Polyamine Analogues | Jeffers et al., 1997 | BE-4-4-4-4/BE-3-7-3/BE-3-3-3 | PA analogue | DU145, LNCaP and PC-3 | BALB/c mice injected with DU145 cells | NR | In vitro and in vivo: Inhibition of PCa cells growth by all three polyamine analogues | [103] |
| Zagaj et al., 1998 | BE-4-4-4-4 | PA analogue | AT3.1, AT6.1 and AT6.3; DU145, DuPro-1 and TSU-Pr1 | Nude mice injected with DuPro-1 and PC-3 PCa cells | NR | In vitro and in vivo: BE-4-4-4-4 was cytotoxic against rat and human PCa cells | [104] | |
| Mi et al., 1998 | DENSpm | PA analogue | LNCaP, DU145, and PC-3 | NR | NR | In vitro: Inhibition of PCa cells growth with varied sensitivity, DU145 > PC-3 > LNCaP | [5] | |
| Eiseman et al., 1998 | BIS | PA analogue | PC-3 and DU-145 | Nude mice injected with PC-3 and DU-145 cells | NR | In vitro and in vivo: Inhibition of PCa growth and enhanced radiosensitivity | [51] | |
| Schipper et al., 2000 | DENSpm | PA analogue | PC-3, TSU-pr1, DU-145, and JCA-1 | Nude mice injected with Du145 cells | NR | In vitro and in vivo: Inhibition of PCa growth | [49] | |
| McCloskey et al., 2000 | CPENSpm, CHENSpm and BE 3-3-3 | PA analogue | LNCaP, PC3, and Du145 | NR | NR | In vitro: Inhibition of PCa cells growth, especially DU145 | [105] | |
| Reddy et al., 2001 | BE-4-4-4-4 | PA analogue | LnCap, DU145, PC-3, and DuPro | NR | NR | In vitro: Inhibition of PCa cells growth with varied sensitivity, LnCap and DU145 > DuPro >PC-3 | [107] | |
| Valasinas et al., 2001 | BE-4-4-4-4 | PA analogue | LnCap, DU145, DuPro, and PC-3 | NR | NR | In vitro: Inhibition of PCa cells growth, PC3 was the least sensitive | [106] | |
| Frydman et al., 2003 | Cyclopropane-Containing Polyamine Analogues | PA analogue | DU-145, DuPro, and PC-3 | NR | NR | In vitro: Inhibition of PCa cells growth, especially DU145 | [110] | |
| Valasinas et al., 2003 | Long-chain Polyamines (Oligoamines) | PA analogue | LnCap, DU-145, DuPro and PC-3 | NR | NR | In vitro: Inhibition of PCa cells growth with varied sensitivity, LnCaP, DU145 > DuPro and PC-3 | [109] | |
| Frydman et al., 2004 | Macrocyclic Polyamines | PA analogue | DuPro and PC-3 | NR | NR | In vitro: The macrocycles were cytotoxic against PCa cells | [108] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Q.; Wong, C.Y.-P.; Cheuk, I.W.-y.; Teoh, J.Y.-C.; Chiu, P.K.-F.; Ng, C.-F. The Emerging Clinical Role of Spermine in Prostate Cancer. Int. J. Mol. Sci. 2021, 22, 4382. https://doi.org/10.3390/ijms22094382
Peng Q, Wong CY-P, Cheuk IW-y, Teoh JY-C, Chiu PK-F, Ng C-F. The Emerging Clinical Role of Spermine in Prostate Cancer. International Journal of Molecular Sciences. 2021; 22(9):4382. https://doi.org/10.3390/ijms22094382
Chicago/Turabian StylePeng, Qiang, Christine Yim-Ping Wong, Isabella Wai-yin Cheuk, Jeremy Yuen-Chun Teoh, Peter Ka-Fung Chiu, and Chi-Fai Ng. 2021. "The Emerging Clinical Role of Spermine in Prostate Cancer" International Journal of Molecular Sciences 22, no. 9: 4382. https://doi.org/10.3390/ijms22094382