
The Kinetics of Lymphatic Dysfunction and Leukocyte Expansion in the Draining Lymph Node during LTB4 Antagonism in a Mouse Model of Lymphedema

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Leukocyte Populations Expand in dLNs during Lymphedema Progression
2.2. T Helper Cells Increase as a Percentage of T Cells in dLNs during Lymphedema Development
2.3. Tail Swelling Correlates with Changes in Leukocyte Populations in dLNs
2.4. Increased Accumulation of Lymph-Draining Nanoparticles within Intact Vessel dLN Compared to Injured Vessel dLN Following Single Vessel Ligation Diminishes as Swelling Progresses
2.5. Increased Leukocyte Expansion in the Intact Vessel dLN Following Single Vessel Ligation
2.6. Bestatin Treatment Increases Nanoparticle Accumulation within the Intact Vessel dLN during Later Stages of Lymphedema Progression
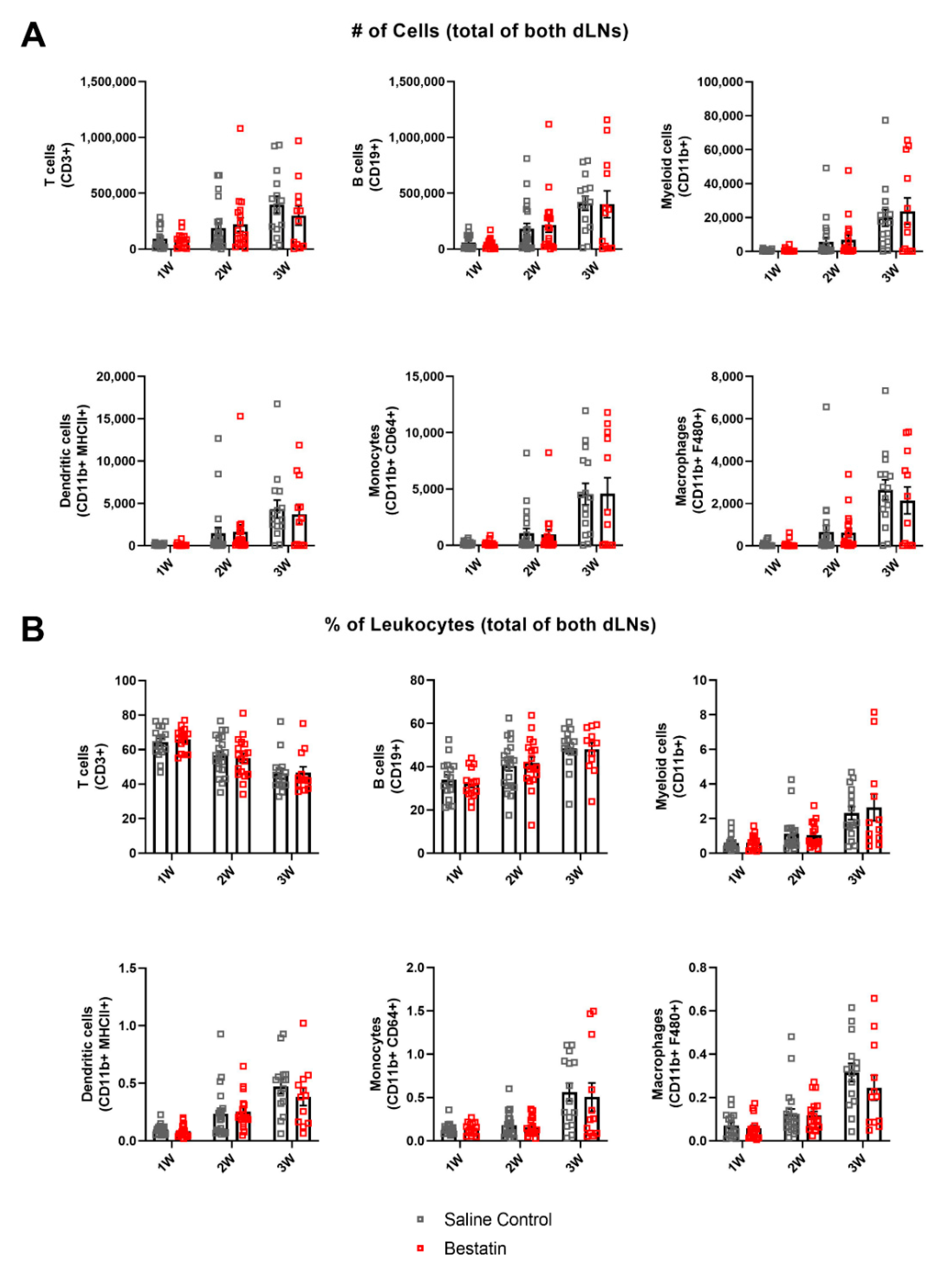
2.7. Bestatin Treatment Has No Significant Effect on Magnitude of Immune Response in dLNs during Lymphedema Progression
2.8. Bestatin Treatment Leads to Leukocyte Expansion in the Intact Vessel dLN Compared to the Injured Vessel dLN
3. Discussion
4. Materials and Methods
4.1. Surgical Lymphedema Model
4.2. Flow Cytometry
4.3. Tail Swelling
4.4. Nanoparticle Synthesis and Characterization
4.5. Nanoparticle NIR Functional Analysis
4.6. Bestatin Treatment
4.7. NIR Lymphatic Functional Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rockson, S.G.; Keeley, V.; Kilbreath, S.; Szuba, A.; Towers, A. Cancer-Associated Secondary Lymphoedema. Nat. Rev. Dis. Primers 2019, 5, 22. [Google Scholar] [CrossRef]
- Ly, C.L.; Kataru, R.P.; Mehrara, B.J. Inflammatory Manifestations of Lymphedema. Int. J. Mol. Sci. 2017, 18, 171. [Google Scholar] [CrossRef]
- Ghanta, S.; Cuzzone, D.A.; Torrisi, J.S.; Albano, N.J.; Joseph, W.J.; Savetsky, I.L.; Gardenier, J.C.; Chang, D.; Zampell, J.C.; Mehrara, B.J. Regulation of Inflammation and Fibrosis by Macrophages in Lymphedema. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H1065–H1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogata, F.; Fujiu, K.; Matsumoto, S.; Nakayama, Y.; Shibata, M.; Oike, Y.; Koshima, I.; Watabe, T.; Nagai, R.; Manabe, I. Excess Lymphangiogenesis Cooperatively Induced by Macrophages and CD4+ T Cells Drives the Pathogenesis of Lymphedema. J. Investig. Dermatol. 2016, 136, 706–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gousopoulos, E.; Proulx, S.T.; Scholl, J.; Uecker, M.; Detmar, M. Prominent Lymphatic Vessel Hyperplasia with Progressive Dysfunction and Distinct Immune Cell Infiltration in Lymphedema. Am. J. Pathol. 2016, 186, 2193–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avraham, T.; Zampell, J.C.; Yan, A.; Elhadad, S.; Weitman, E.S.; Rockson, S.G.; Bromberg, J.; Mehrara, B.J. Th2 Differentiation Is Necessary for Soft Tissue Fibrosis and Lymphatic Dysfunction Resulting from Lymphedema. FASEB J. 2013, 27, 1114–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabibiazar, R.; Cheung, L.; Han, J.; Swanson, J.; Beilhack, A.; An, A.; Dadras, S.S.; Rockson, N.; Joshi, S.; Wagner, R.; et al. Inflammatory Manifestations of Experimental Lymphatic Insufficiency. PLoS Med. 2006, 3, e254. [Google Scholar] [CrossRef] [Green Version]
- Gousopoulos, E.; Proulx, S.T.; Bachmann, S.B.; Scholl, J.; Dionyssiou, D.; Demiri, E.; Halin, C.; Dieterich, L.C.; Detmar, M. Regulatory T Cell Transfer Ameliorates Lymphedema and Promotes Lymphatic Vessel Function. JCI Insight 2016, 1, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gardenier, J.C.; Kataru, R.P.; Hespe, G.E.; Savetsky, I.L.; Torrisi, J.S.; Nores, G.D.; Jowhar, D.K.; Nitti, M.D.; Schofield, R.C.; Carlow, D.C.; et al. Topical Tacrolimus for the Treatment of Secondary Lymphedema. Nat. Commun. 2017, 8, 14345. [Google Scholar] [CrossRef]
- Nakamura, K.; Radhakrishnan, K.; Wong, Y.M.; Rockson, S.G. Anti-Inflammatory Pharmacotherapy with Ketoprofen Ameliorates Experimental Lymphatic Vascular Insufficiency in Mice. PLoS ONE 2009, 4, e8380. [Google Scholar] [CrossRef] [Green Version]
- Yokomizo, T.; Izumi, T.; Shimizu, T. Leukotriene B4: Metabolism and Signal Transduction. Arch. Biochem. Biophys. 2001, 385, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Rockson, S.G.; Jiang, X.; Kim, J.; Begaye, A.; Shuffle, E.M.; Tu, A.B.; Cribb, M.; Nepiyushchikh, Z.; Feroze, A.H.; et al. Leukotriene B4 Antagonism Ameliorates Experimental Lymphedema. Sci. Transl. Med. 2017, 9, eaal3920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García Nores, G.D.; Ly, C.L.; Cuzzone, D.A.; Kataru, R.P.; Hespe, G.E.; Torrisi, J.S.; Huang, J.J.; Gardenier, J.C.; Savetsky, I.L.; Nitti, M.D.; et al. CD4+ T Cells Are Activated in Regional Lymph Nodes and Migrate to Skin to Initiate Lymphedema. Nat. Commun. 2018, 9, 1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt, A.M.; Rutkowski, J.M.; Martel, C.; Kuan, E.L.; Ivanov, S.; Swartz, M.A.; Randolph, G.J. Normal Dendritic Cell Mobilization to Lymph Nodes under Conditions of Severe Lymphatic Hypoplasia. J. Immunol. 2013, 190, 4608–4620. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.N.; Rutkowski, J.M.; Pasquier, M.; Kuan, E.L.; Alitalo, K.; Randolph, G.J.; Swartz, M.A. Impaired Humoral Immunity and Tolerance in K14-VEGFR-3-Ig Mice That Lack Dermal Lymphatic Drainage. J. Immunol. 2012, 189, 2181–2190. [Google Scholar] [CrossRef] [Green Version]
- Armer, J.M.; Stewart, B.R. Post-breast cancer lymphedema: Incidence increases from 12 to 30 to 60 months. Lymphology 2010, 43, 118–127. [Google Scholar]
- Yoon, Y.S.; Murayama, T.; Gravereaux, E.; Tkebuchava, T.; Silver, M.; Curry, C.; Wecker, A.; Kirchmair, R.; Hu, C.S.; Kearney, M.; et al. VEGF-C gene therapy augments postnatal lymphangiogenesis and ameliorates secondary lymphedema. J. Clin. Investig. 2003, 111, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Rutkowski, J.M.; Moya, M.; Johannes, J.; Goldman, J.; Swartz, M.A. Secondary Lymphedema in the Mouse Tail: Lymphatic Hyperplasia, VEGF-C Upregulation, and the Protective Role of MMP-9. Microvasc. Res. 2006, 72, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Weiler, M.J.; Cribb, M.T.; Nepiyushchikh, Z.; Nelson, T.S.; Dixon, J.B. A Novel Mouse Tail Lymphedema Model for Observing Lymphatic Pump Failure during Lymphedema Development. Sci. Rep. 2019, 9, 10405. [Google Scholar] [CrossRef] [Green Version]
- Schudel, A.; Sestito, L.F.; Thomas, S.N. S-nitrosated poly(propylene sulfide) nanoparticles for enhanced nitric oxide delivery to lymphatic tissues. J. Biomed. Mater. Res. A 2018, 106, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Sestito, L.F.; Thomas, S.N. Lymph-Directed Nitric Oxide Increases Immune Cell Access to Lymph-Borne Nanoscale Solutes. Biomaterials 2021, 265, 120411. [Google Scholar] [CrossRef]
- Reddy, S.T.; Rehor, A.; Schmoekel, H.G.; Hubbell, J.A.; Swartz, M.A. In Vivo Targeting of Dendritic Cells in Lymph Nodes with Poly(Propylene Sulfide) Nanoparticles. J. Control. Release 2006, 112, 26–34. [Google Scholar] [CrossRef]
- Thomas, S.N.; Vokali, E.; Lund, A.W.; Hubbell, J.A.; Swartz, M.A. Targeting the Tumor-Draining Lymph Node with Adjuvanted Nanoparticles Reshapes the Anti-Tumor Immune Response. Biomaterials 2014, 35, 814–824. [Google Scholar] [CrossRef] [Green Version]
- Schudel, A.; Chapman, A.P.; Yau, M.K.; Higginson, C.J.; Francis, D.M.; Manspeaker, M.P.; Avecilla, A.R.; Rohner, N.A.; Finn, M.G.; Thomas, S.N. Programmable Multistage Drug Delivery to Lymph Nodes. Nat. Nanotechnol. 2020, 15, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, K.; Goodarzi, M.; Tager, A.M.; Luster, A.D.; von Andrian, U.H. Leukotriene B4 and BLT1 Control Cytotoxic Effector T Cell Recruitment to Inflamed Tissues. Nat. Immunol. 2003, 4, 965–973. [Google Scholar] [CrossRef]
- Lämmermann, T.; Afonso, P.V.; Angermann, B.R.; Wang, J.M.; Kastenmüller, W.; Parent, C.A.; Germain, R.N. Neutrophil Swarms Require LTB4 and Integrins at Sites of Cell Death in Vivo. Nature 2013, 498, 371–375. [Google Scholar] [CrossRef]
- Weiler, M.; Kassis, T.; Dixon, J.B. Sensitivity Analysis of Near-Infrared Functional Lymphatic Imaging. J. Biomed. Opt. 2012, 17, 066019. [Google Scholar] [CrossRef] [PubMed]
- Weiler, M.; Dixon, J.B. Differential Transport Function of Lymphatic Vessels in the Rat Tail Model and the Long-Term Effects of Indocyanine Green as Assessed with near-Infrared Imaging. Front. Physiol. 2013, 4, 215. [Google Scholar] [CrossRef] [Green Version]
- Nelson, T.S.; Akin, R.E.; Weiler, M.J.; Kassis, T.; Kornuta, J.A.; Dixon, J.B. Minimally Invasive Method for Determining the Effective Lymphatic Pumping Pressure in Rats Using Near-Infrared Imaging. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 306, R281–R290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, M.C.; Teijeira, A.; Halin, C. T Cell Trafficking through Lymphatic Vessels. Front. Immunol. 2016, 7, 613. [Google Scholar] [CrossRef] [Green Version]
- Teijeira, A.; Hunter, M.C.; Russo, E.; Proulx, S.T.; Frei, T.; Debes, G.F.; Coles, M.; Melero, I.; Detmar, M.; Rouzaut, A.; et al. T Cell Migration from Inflamed Skin to Draining Lymph Nodes Requires Intralymphatic Crawling Supported by ICAM-1/LFA-1 Interactions. Cell Rep. 2017, 18, 857–865. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.Y.; Vogt, T.K.; Favre, S.; Scarpellino, L.; Huang, H.Y.; Tacchini-Cottier, F.; Luther, S.A. Trapping of Naive Lymphocytes Triggers Rapid Growth and Remodeling of the Fibroblast Network in Reactive Murine Lymph Nodes. Proc. Natl. Acad. Sci. USA 2014, 111, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.W.; Yeo, K.P.; Wong, F.H.; Lim, H.Y.; Khoo, K.L.; Abastado, J.P.; Angeli, V. Expansion of Cortical and Medullary Sinuses Restrains Lymph Node Hypertrophy during Prolonged Inflammation. J. Immunol. 2012, 188, 4065–4080. [Google Scholar] [CrossRef] [Green Version]
- Buckley, C.D.; Barone, F.; Nayar, S.; Bénézech, C.; Caamaño, J. Stromal Cells in Chronic Inflammation and Tertiary Lymphoid Organ Formation. Annu. Rev. Immunol. 2015, 33, 715–745. [Google Scholar] [CrossRef]
- Zampell, J.C.; Yan, A.; Elhadad, S.; Avraham, T.; Weitman, E.; Mehrara, B.J. CD4+ Cells Regulate Fibrosis and Lymphangiogenesis in Response to Lymphatic Fluid Stasis. PLoS ONE 2012, 7, e49940. [Google Scholar] [CrossRef] [Green Version]
- Angeli, V.; Ginhoux, F.; Llodrà, J.; Quemeneur, L.; Frenette, P.S.; Skobe, M.; Jessberger, R.; Merad, M.; Randolph, G.J. B Cell-Driven Lymphangiogenesis in Inflamed Lymph Nodes Enhances Dendritic Cell Mobilization. Immunity 2006, 24, 203–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrisi, J.S.; Hespe, G.E.; Cuzzone, D.A.; Savetsky, I.L.; Nitti, M.D.; Gardenier, J.C.; Nores, G.D.; Jowhar, D.; Kataru, R.P.; Mehrara, B.J. Inhibition of Inflammation and INOS Improves Lymphatic Function in Obesity. Sci. Rep. 2016, 6, 19817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, S.; Randolph, G.J. Myeloid Cells Pave the Way for Lymphatic System Development and Maintenance. Pflüg. Arch. Eur. J. Physiol. 2017, 469, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Jakubzick, C.; Gautier, E.L.; Gibbings, S.L.; Sojka, D.K.; Schlitzer, A.; Johnson, T.E.; Ivanov, S.; Duan, Q.; Bala, S.; Condon, T.; et al. Minimal Differentiation of Classical Monocytes as They Survey Steady-State Tissues and Transport Antigen to Lymph Nodes. Immunity 2013, 39, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Zou, L.; Zhao, L.; Yang, W.; Xiong, Y.; Li, B.; He, R. Leukotriene B4-Leukotriene B4 Receptor Axis Promotes Oxazolone-Induced Contact Dermatitis by Directing Skin Homing of Neutrophils and CD8+ T Cells. Immunology 2015, 146, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Rockson, S.G.; Tian, W.; Jiang, X.; Kuznetsova, T.; Haddad, F.; Zampell, J.; Mehrara, B.; Sampson, J.P.; Roche, L.; Kim, J.; et al. Pilot Studies Demonstrate the Potential Benefits of Antiinflammatory Therapy in Human Lymphedema. JCI Insight 2018, 3, 1–11. [Google Scholar] [CrossRef]
- Miyahara, N.; Ohnishi, H.; Miyahara, S.; Takeda, K.; Matsubara, S.; Matsuda, H.; Okamoto, M.; Loader, J.E.; Joetham, A.; Tanimoto, M.; et al. Leukotriene B4 Release from Mast Cells in IgE-Mediated Airway Hyperresponsiveness and Inflammation. Am. J. Respir. Cell Mol. Biol. 2009, 40, 672–682. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Ba, X.; Cui, L.; Xue, Y.; Zeng, X. Leukotriene B4 Regulates Proliferation and Differentiation of Cultured Rat Myoblasts via the BLT1 Pathway. Mol. Cells 2009, 27, 403–408. [Google Scholar] [CrossRef]
- Liu, X.; Yang, T.; Miao, L.; Mei, Y.A.; Hu, C. Leukotriene B4 Inhibits L-Type Calcium Channels via P38 Signaling Pathway in Vascular Smooth Muscle Cells. Cell. Physiol. Biochem. 2015, 37, 1903–1913. [Google Scholar] [CrossRef] [PubMed]
- Johnston, M.G.; Kanalec, A.; Gordon, J.L. Effects of arachidonic acid and its cyclo-oxygenase and lipoxygenase products on lymphatic vessel contractility in vitro. Prostaglandins 1983, 25, 85–98. [Google Scholar] [CrossRef]
- Ferguson, M.K.; Shahinian, H.K.; Michelassi, F. Lymphatic smooth muscle responses to leukotrienes, histamine and platelet activating factor. J. Surg. Res. 1988, 44, 172–177. [Google Scholar] [CrossRef]
- Rehor, A.; Hubbell, J.A.; Tirelli, N. Oxidation-Sensitive Polymeric Nanoparticles. Langmuir 2005, 21, 411–417. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cribb, M.T.; Sestito, L.F.; Rockson, S.G.; Nicolls, M.R.; Thomas, S.N.; Dixon, J.B. The Kinetics of Lymphatic Dysfunction and Leukocyte Expansion in the Draining Lymph Node during LTB4 Antagonism in a Mouse Model of Lymphedema. Int. J. Mol. Sci. 2021, 22, 4455. https://doi.org/10.3390/ijms22094455
Cribb MT, Sestito LF, Rockson SG, Nicolls MR, Thomas SN, Dixon JB. The Kinetics of Lymphatic Dysfunction and Leukocyte Expansion in the Draining Lymph Node during LTB4 Antagonism in a Mouse Model of Lymphedema. International Journal of Molecular Sciences. 2021; 22(9):4455. https://doi.org/10.3390/ijms22094455
Chicago/Turabian StyleCribb, Matthew T., Lauren F. Sestito, Stanley G. Rockson, Mark R. Nicolls, Susan N. Thomas, and J. Brandon Dixon. 2021. "The Kinetics of Lymphatic Dysfunction and Leukocyte Expansion in the Draining Lymph Node during LTB4 Antagonism in a Mouse Model of Lymphedema" International Journal of Molecular Sciences 22, no. 9: 4455. https://doi.org/10.3390/ijms22094455