
Tryptophanyl-tRNA Synthetase as a Potential Therapeutic Target



Abstract
:1. Introduction
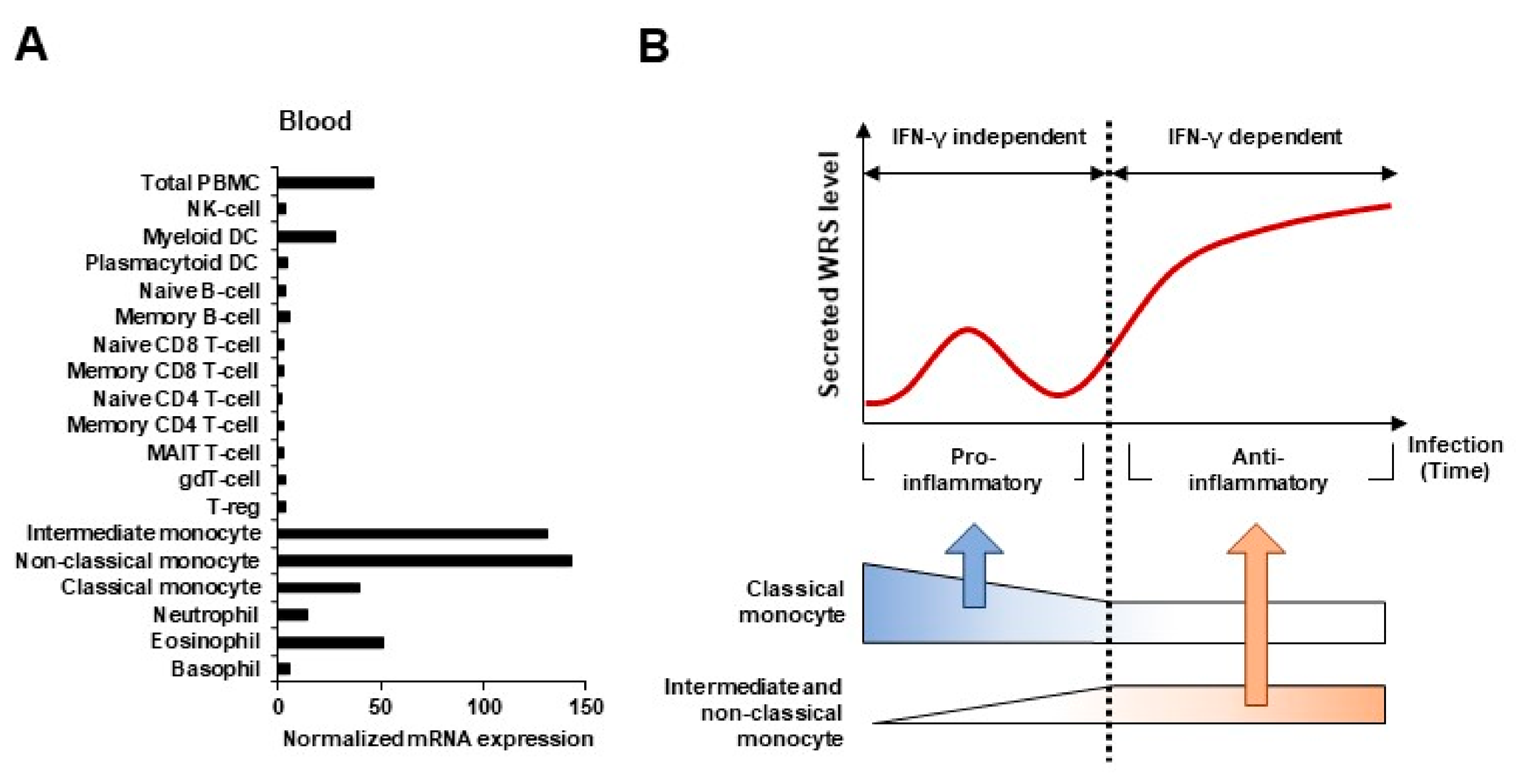
2. WRS as a Biomarker of Immune Response
2.1. WRS Expression in Systemic Inflammatory Response
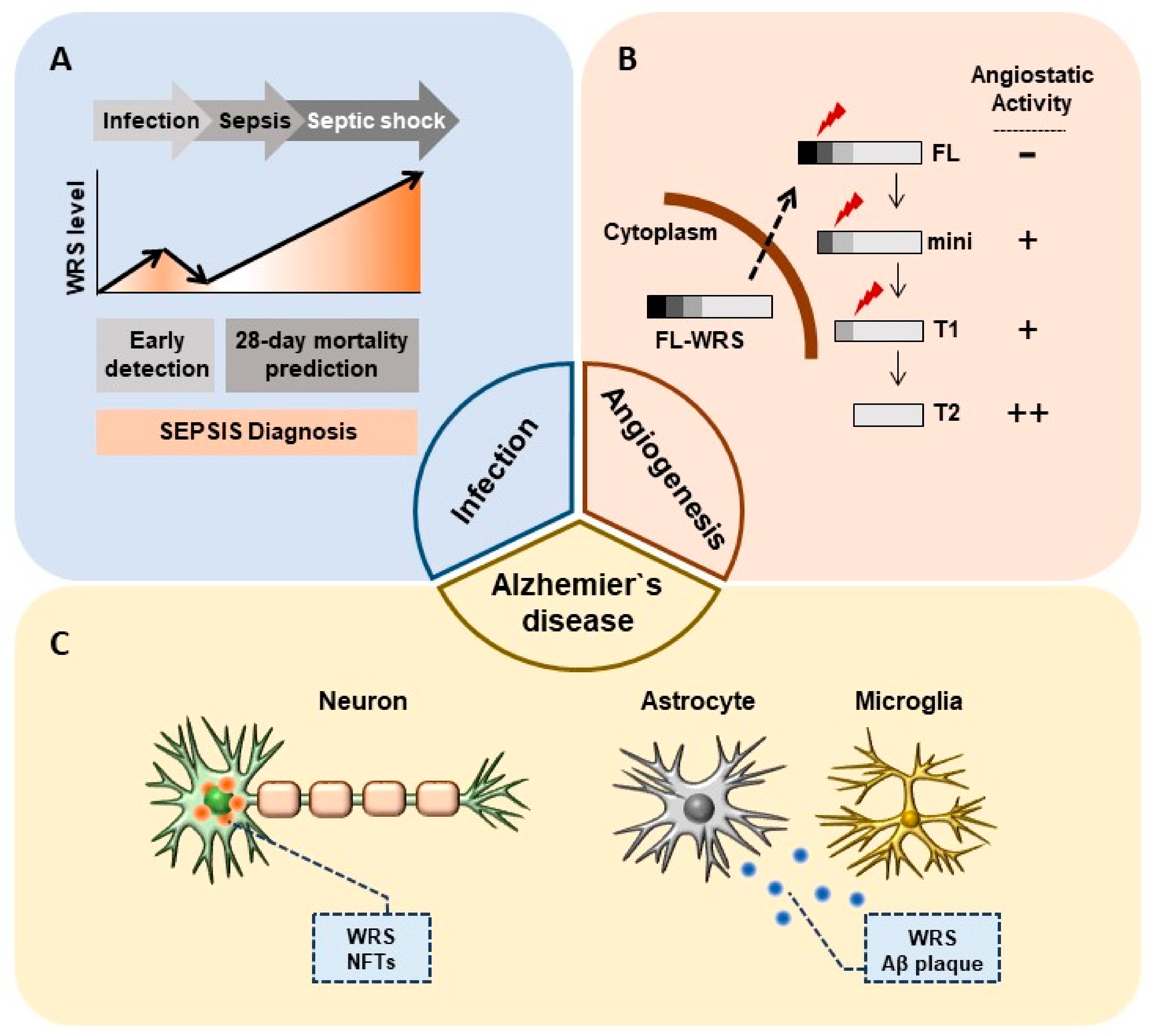
2.2. WRS as a Prognostic Biomarker in Sepsis
3. WRS as a Therapeutic Target in Cancer
3.1. WRS as a Target for Anti-Angiogenic Therapy
3.2. Implications of Angiostaic WRS in Cancer Metastasis
4. Pathological Role of WRS in Alzheimer’s Disease
5. Immunological Role of WRS in Trp Metabolism
5.1. Increased Trp Production by WRS in Cancer Cell
5.2. Implications of Increased WRS in Tolerogenic Immune Response
6. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yao, P.; Fox, P.L. Aminoacyl-tRNA synthetases in medicine and disease. EMBO Mol. Med. 2013, 5, 332–343. [Google Scholar] [CrossRef]
- Kim, S.; You, S.; Hwang, D. Aminoacyl-tRNA synthetases and tumorigenesis: More than housekeeping. Nat. Rev. Cancer 2011, 11, 708–718. [Google Scholar] [CrossRef]
- Fuchs, S.A.; Schene, I.F.; Kok, G.; Jansen, J.M.; Nikkels, P.G.J.; van Gassen, K.L.I.; Terheggen-Lagro, S.W.J.; van der Crabben, S.N.; Hoeks, S.E.; Niers, L.E.M.; et al. Aminoacyl-tRNA synthetase deficiencies in search of common themes. Genet. Med. 2019, 21, 319–330. [Google Scholar] [CrossRef]
- Lee, S.W.; Cho, B.H.; Park, S.G.; Kim, S. Aminoacyl-tRNA synthetase complexes: Beyond translation. J. Cell Sci. 2004, 117, 3725–3734. [Google Scholar] [CrossRef] [Green Version]
- Kwon, N.H.; Fox, P.L.; Kim, S. Aminoacyl-tRNA synthetases as therapeutic targets. Nat. Rev. Drug Discov. 2019, 18, 629–650. [Google Scholar] [CrossRef]
- Guo, M.; Schimmel, P.; Yang, X.L. Functional expansion of human tRNA synthetases achieved by structural inventions. FEBS Lett. 2010, 584, 434–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisselev, L.L.; Favorova, O.O.; Kovaleva, G.K. Tryptophanyl-tRNA synthetase from beef pancreas. Methods Enzymol. 1979, 59, 234–257. [Google Scholar] [PubMed]
- Sallafranque, M.L.; Garret, M.; Benedetto, J.P.; Fournier, M.; Labouesse, B.; Bonnet, J. Tryptophanyl-tRNA synthetase is a major soluble protein species in bovine pancreas. Biochim. Biophys. Acta 1986, 882, 192–199. [Google Scholar] [CrossRef]
- Favorova, O.O.; Zargarova, T.A.; Rukosuyev, V.S.; Beresten, S.F.; Kisselev, L.L. Molecular and cellular studies of tryptophanyl-tRNA synthetases using monoclonal antibodies. Remarkable variations in the content of tryptophanyl-tRNA synthetase in the pancreas of different mammals. Eur. J. Biochem. 1989, 184, 583–588. [Google Scholar] [CrossRef]
- Tuzikov, F.V.; Tuzikova, N.A.; Vavilin, V.I.; Zinov’ev, V.V.; Malygin, E.G.; Favorova, O.O.; Zargarova, T.A.; Sudomoina, M.A.; Kiselev, L.L. Aggregation of tryptophanyl-tRNA synthetase depending on temperature. Study by a low-angle scatter x-ray method. Mol. Biol. 1991, 25, 740–751. [Google Scholar]
- Paley, E.L.; Smelyanski, L.; Malinovskii, V.; Subbarayan, P.R.; Berdichevsky, Y.; Posternak, N.; Gershoni, J.M.; Sokolova, O.; Denisova, G. Mapping and molecular characterization of novel monoclonal antibodies to conformational epitopes on NH2 and COOH termini of mammalian tryptophanyl-tRNA synthetase reveal link of the epitopes to aggregation and Alzheimer’s disease. Mol. Immunol. 2007, 44, 541–557. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Yang, X.L. Architecture and metamorphosis. Top. Curr. Chem. 2014, 344, 89–118. [Google Scholar] [PubMed] [Green Version]
- Jin, M. Unique roles of tryptophanyl-tRNA synthetase in immune control and its therapeutic implications. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyeon, D.Y.; Kim, J.H.; Ahn, T.J.; Cho, Y.; Hwang, D.; Kim, S. Evolution of the multi-tRNA synthetase complex and its role in cancer. J. Biol. Chem. 2019, 294, 5340–5351. [Google Scholar] [CrossRef] [Green Version]
- Ray, P.S.; Sullivan, J.C.; Jia, J.; Francis, J.; Finnerty, J.R.; Fox, P.L. Evolution of function of a fused metazoan tRNA synthetase. Mol. Biol. Evol. 2011, 28, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Cahuzac, B.; Berthonneau, E.; Birlirakis, N.; Guittet, E.; Mirande, M. A recurrent RNA-binding domain is appended to eukaryotic aminoacyl-tRNA synthetases. EMBO J. 2000, 19, 445–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, J.; Arif, A.; Ray, P.S.; Fox, P.L. WHEP domains direct noncanonical function of glutamyl-Prolyl tRNA synthetase in translational control of gene expression. Mol. Cell 2008, 29, 679–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, R.; Jia, J.; Arif, A.; Ray, P.S.; Fox, P.L. The GAIT system: A gatekeeper of inflammatory gene expression. Trends Biochem. Sci. 2009, 34, 324–331. [Google Scholar] [CrossRef] [Green Version]
- Fleckner, J.; Martensen, P.M.; Tolstrup, A.B.; Kjeldgaard, N.O.; Justesen, J. Differential regulation of the human, interferon inducible tryptophanyl-tRNA synthetase by various cytokines in cell lines. Cytokine 1995, 7, 70–77. [Google Scholar] [CrossRef]
- Jobin, P.G.; Solis, N.; Machado, Y.; Bell, P.A.; Kwon, N.H.; Kim, S.; Overall, C.M.; Butler, G.S. Matrix metalloproteinases inactivate the proinflammatory functions of secreted moonlighting tryptophanyl-tRNA synthetase. J. Biol. Chem. 2019, 294, 12866–12879. [Google Scholar] [CrossRef] [Green Version]
- Kang, I.; Lee, B.C.; Lee, J.Y.; Kim, J.J.; Lee, S.E.; Shin, N.; Choi, S.W.; Kang, K.S. Interferon-gamma-mediated secretion of tryptophanyl-tRNA synthetases has a role in protection of human umbilical cord blood-derived mesenchymal stem cells against experimental colitis. BMB Rep. 2019, 52, 318–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, M.; Zhou, Q.; Otero, F.; Myers, C.A.; Bates, A.; Belani, R.; Liu, J.; Luo, J.K.; Tzima, E.; Zhang, D.E.; et al. Evidence for annexin II-S100A10 complex and plasmin in mobilization of cytokine activity of human TrpRS. J. Biol. Chem. 2008, 283, 2070–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kisselev, L.; Frolova, L.; Haenni, A.L. Interferon inducibility of mammalian tryptophanyl-tRNA synthetase: New perspectives. Trends Biochem. Sci. 1993, 18, 263–267. [Google Scholar] [CrossRef]
- Ahn, Y.H.; Park, S.; Choi, J.J.; Park, B.K.; Rhee, K.H.; Kang, E.; Ahn, S.; Lee, C.H.; Lee, J.S.; Inn, K.S.; et al. Secreted tryptophanyl-tRNA synthetase as a primary defence system against infection. Nat. Microbiol. 2016, 2, 16191. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Lee, E.S.; Uddin, M.B.; Kim, T.H.; Kim, J.H.; Chathuranga, K.; Chathuranga, W.A.G.; Jin, M.; Kim, S.; Kim, C.J.; et al. Released Tryptophanyl-tRNA Synthetase Stimulates Innate Immune Responses against Viral Infection. J. Virol. 2019, 93, e01291-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakasugi, K.; Slike, B.M.; Hood, J.; Otani, A.; Ewalt, K.L.; Friedlander, M.; Cheresh, D.A.; Schimmel, P. A human aminoacyl-tRNA synthetase as a regulator of angiogenesis. Proc. Natl. Acad. Sci. USA 2002, 99, 173–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolstrup, A.B.; Bejder, A.; Fleckner, J.; Justesen, J. Transcriptional regulation of the interferon-gamma-inducible tryptophanyl-tRNA synthetase includes alternative splicing. J. Biol. Chem. 1995, 270, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Turpaev, K.T.; Zakhariev, V.M.; Sokolova, I.V.; Narovlyansky, A.N.; Amchenkova, A.M.; Justesen, J.; Frolova, L.Y. Alternative processing of the tryptophanyl-tRNA synthetase mRNA from interferon-treated human cells. Eur. J. Biochem. 1996, 240, 732–737. [Google Scholar] [CrossRef]
- Lindner, D.J. Interferons as antiangiogenic agents. Curr Oncol. Rep. 2002, 4, 510–514. [Google Scholar] [CrossRef]
- Otani, A.; Slike, B.M.; Dorrell, M.I.; Hood, J.; Kinder, K.; Ewalt, K.L.; Cheresh, D.; Schimmel, P.; Friedlander, M. A fragment of human TrpRS as a potent antagonist of ocular angiogenesis. Proc. Natl. Acad. Sci. USA 2002, 99, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Tzima, E.; Reader, J.S.; Irani-Tehrani, M.; Ewalt, K.L.; Schwartz, M.A.; Schimmel, P. Biologically active fragment of a human tRNA synthetase inhibits fluid shear stress-activated responses of endothelial cells. Proc. Natl. Acad. Sci. USA 2003, 100, 14903–14907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzima, E.; Reader, J.S.; Irani-Tehrani, M.; Ewalt, K.L.; Schwartz, M.A.; Schimmel, P. VE-cadherin links tRNA synthetase cytokine to anti-angiogenic function. J. Biol. Chem. 2005, 280, 2405–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paley, E.L.; Denisova, G.; Sokolova, O.; Posternak, N.; Wang, X.; Brownell, A.L. Tryptamine induces tryptophanyl-tRNA synthetase-mediated neurodegeneration with neurofibrillary tangles in human cell and mouse models. Neuromolecular. Med. 2007, 9, 55–82. [Google Scholar] [CrossRef]
- Choi, J.S.; Yoon, B.R.; Shin, J.H.; Lee, S.H.; Leem, A.Y.; Park, M.S.; Kim, Y.S.; Chung, K.S. Clinical value of full-length tryptophanyl-tRNA synthetase for sepsis detection in critically ill patients-A retrospective clinical assessment. Int. J. Infect. Dis. 2020, 97, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Arif, A.; Jia, J.; Mukhopadhyay, R.; Willard, B.; Kinter, M.; Fox, P.L. Two-site phosphorylation of EPRS coordinates multimodal regulation of noncanonical translational control activity. Mol. Cell 2009, 35, 164–180. [Google Scholar] [CrossRef] [Green Version]
- Yao, P.; Potdar, A.A.; Arif, A.; Ray, P.S.; Mukhopadhyay, R.; Willard, B.; Xu, Y.; Yan, J.; Saidel, G.M.; Fox, P.L. Coding region polyadenylation generates a truncated tRNA synthetase that counters translation repression. Cell 2012, 149, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.Y.; Maeng, S.J.; Cho, H.J.; Choi, Y.S.; Chung, J.M.; Lee, S.; Kim, H.K.; Kim, J.H.; Eom, C.Y.; Kim, Y.G.; et al. Assembly of Multi-tRNA Synthetase Complex via Heterotetrameric Glutathione Transferase-homology Domains. J. Biol Chem 2015, 290, 29313–29328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, N.H.; Kang, T.; Lee, J.Y.; Kim, H.H.; Kim, H.R.; Hong, J.; Oh, Y.S.; Han, J.M.; Ku, M.J.; Lee, S.Y.; et al. Dual role of methionyl-tRNA synthetase in the regulation of translation and tumor suppressor activity of aminoacyl-tRNA synthetase-interacting multifunctional protein-3. Proc. Natl. Acad. Sci. USA 2011, 108, 19635–19640. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Kim, D.G.; Kim, B.G.; Yang, W.S.; Hong, J.; Kang, T.; Oh, Y.S.; Kim, K.R.; Han, B.W.; Hwang, B.J.; et al. Promiscuous methionyl-tRNA synthetase mediates adaptive mistranslation to protect cells against oxidative stress. J. Cell Sci. 2014, 127, 4234–4245. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Wei, Z.; Zhou, J.J.; Ye, F.; Lo, W.S.; Wang, F.; Lau, C.F.; Wu, J.; Nangle, L.A.; Chiang, K.P.; et al. Internally deleted human tRNA synthetase suggests evolutionary pressure for repurposing. Structure 2012, 20, 1470–1477. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.J.; Wang, F.; Xu, Z.; Lo, W.S.; Lau, C.F.; Chiang, K.P.; Nangle, L.A.; Ashlock, M.A.; Mendlein, J.D.; Yang, X.L.; et al. Secreted histidyl-tRNA synthetase splice variants elaborate major epitopes for autoantibodies in inflammatory myositis. J. Biol. Chem. 2014, 289, 19269–19275. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Yang, X.L.; Schimmel, P. New functions of aminoacyl-tRNA synthetases beyond translation. Nat. Rev. Mol. Cell Biol. 2010, 11, 668–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.C.; Kang, T.; Jin, D.; Han, J.M.; Kim, S.B.; Park, Y.J.; Cho, K.; Park, Y.W.; Guo, M.; He, W.; et al. Secreted human glycyl-tRNA synthetase implicated in defense against ERK-activated tumorigenesis. Proc. Natl. Acad Sci. USA 2012, 109, E640–E647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.R.; Kim, H.J.; Yang, S.R.; Park, C.H.; Lee, H.Y.; Hong, I.S. A novel endogenous damage signal, glycyl tRNA synthetase, activates multiple beneficial functions of mesenchymal stem cells. Cell Death Differ. 2018, 25, 2023–2036. [Google Scholar] [CrossRef]
- Fagerberg, L.; Hallstrom, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell Proteomics 2014, 13, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Ravenhill, B.J.; Soday, L.; Houghton, J.; Antrobus, R.; Weekes, M.P. Comprehensive cell surface proteomics defines markers of classical, intermediate and non-classical monocytes. Sci. Rep. 2020, 10, 4560. [Google Scholar] [CrossRef]
- Narasimhan, P.B.; Marcovecchio, P.; Hamers, A.A.J.; Hedrick, C.C. Nonclassical Monocytes in Health and Disease. Annu Rev. Immunol. 2019, 37, 439–456. [Google Scholar] [CrossRef]
- Krause, S.W.; Rehli, M.; Kreutz, M.; Schwarzfischer, L.; Paulauskis, J.D.; Andreesen, R. Differential screening identifies genetic markers of monocyte to macrophage maturation. J. Leukoc Biol. 1996, 60, 540–545. [Google Scholar] [CrossRef]
- Matsunaga, T.; Ishida, T.; Takekawa, M.; Nishimura, S.; Adachi, M.; Imai, K. Analysis of gene expression during maturation of immature dendritic cells derived from peripheral blood monocytes. Scand. J. Immunol. 2002, 56, 593–601. [Google Scholar] [CrossRef]
- Shi, C.; Pamer, E.G. Monocyte recruitment during infection and inflammation. Nat. Rev. Immunol. 2011, 11, 762–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, R.; Kanti Barman, P.; Kumar Thatoi, P.; Tripathy, R.; Kumar Das, B.; Ravindran, B. Non-Classical monocytes display inflammatory features: Validation in Sepsis and Systemic Lupus Erythematous. Sci. Rep. 2015, 5, 13886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fingerle, G.; Pforte, A.; Passlick, B.; Blumenstein, M.; Strobel, M.; Ziegler-Heitbrock, H.W. The novel subset of CD14+/CD16+ blood monocytes is expanded in sepsis patients. Blood 1993, 82, 3170–3176. [Google Scholar] [CrossRef]
- Tanaka, H.; Sugimoto, H.; Yoshioka, T.; Sugimoto, T. Role of granulocyte elastase in tissue injury in patients with septic shock complicated by multiple-organ failure. Ann. Surg. 1991, 213, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Pacher, R.; Redl, H.; Frass, M.; Petzl, D.H.; Schuster, E.; Woloszczuk, W. Relationship between neopterin and granulocyte elastase plasma levels and the severity of multiple organ failure. Crit. Care Med. 1989, 17, 221–226. [Google Scholar] [CrossRef]
- Van der Poll, T.; Opal, S.M. Host-pathogen interactions in sepsis. Lancet Infect. Dis. 2008, 8, 32–43. [Google Scholar] [CrossRef]
- Husabo, G.; Nilsen, R.M.; Flaatten, H.; Solligard, E.; Frich, J.C.; Bondevik, G.T.; Braut, G.S.; Walshe, K.; Harthug, S.; Hovlid, E. Early diagnosis of sepsis in emergency departments, time to treatment, and association with mortality: An observational study. PLoS ONE 2020, 15, e0227652. [Google Scholar] [CrossRef]
- Kargaltseva, N.M.; Kotcherovets, V.I.; Mironov, A.Y.; Borisova, O.Y.; Burbello, A.T. Inflammation markers and bloodstream infection (review of literature). Klin. Lab. Diagn. 2019, 64, 435–442. [Google Scholar] [CrossRef]
- Bossink, A.W.; Groeneveld, A.B.; Thijs, L.G. Prediction of microbial infection and mortality in medical patients with fever: Plasma procalcitonin, neutrophilic elastase-alpha1-antitrypsin, and lactoferrin compared with clinical variables. Clin. Infect. Dis. 1999, 29, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Ryoo, S.M.; Han, K.S.; Ahn, S.; Shin, T.G.; Hwang, S.Y.; Chung, S.P.; Hwang, Y.J.; Park, Y.S.; Jo, Y.H.; Chang, H.L.; et al. The usefulness of C-reactive protein and procalcitonin to predict prognosis in septic shock patients: A multicenter prospective registry-based observational study. Sci. Rep. 2019, 9, 6579. [Google Scholar] [CrossRef]
- Tzima, E.; Schimmel, P. Inhibition of tumor angiogenesis by a natural fragment of a tRNA synthetase. Trends Biochem. Sci. 2006, 31, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Kise, Y.; Lee, S.W.; Park, S.G.; Fukai, S.; Sengoku, T.; Ishii, R.; Yokoyama, S.; Kim, S.; Nureki, O. A short peptide insertion crucial for angiostatic activity of human tryptophanyl-tRNA synthetase. Nat. Struct. Mol. Biol. 2004, 11, 149–156. [Google Scholar] [CrossRef]
- Zhou, Q.; Kapoor, M.; Guo, M.; Belani, R.; Xu, X.; Kiosses, W.B.; Hanan, M.; Park, C.; Armour, E.; Do, M.H.; et al. Orthogonal use of a human tRNA synthetase active site to achieve multifunctionality. Nat. Struct. Mol. Biol. 2010, 17, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Morita, A.; Miyagi, E.; Yasumitsu, H.; Kawasaki, H.; Hirano, H.; Hirahara, F. Proteomic search for potential diagnostic markers and therapeutic targets for ovarian clear cell adenocarcinoma. Proteomics 2006, 6, 5880–5890. [Google Scholar] [CrossRef]
- Arnouk, H.; Merkley, M.A.; Podolsky, R.H.; Stoppler, H.; Santos, C.; Alvarez, M.; Mariategui, J.; Ferris, D.; Lee, J.R.; Dynan, W.S. Characterization of Molecular Markers Indicative of Cervical Cancer Progression. Proteom. Clin. Appl. 2009, 3, 516–527. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.W.; Chang, K.P.; Chen, Y.Y.; Liang, Y.; Hsueh, C.; Yu, J.S.; Chang, Y.S.; Yu, C.J. Overexpressed tryptophanyl-tRNA synthetase, an angiostatic protein, enhances oral cancer cell invasiveness. Oncotarget 2015, 6, 21979–21992. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.P.; Yu, X.H.; Zhou, J. Tryptophanyl-tRNA synthetase (WARS) expression in uveal melanoma-Possible contributor during uveal melanoma progression. Biosci. Biotechnol. Biochem. 2020, 84, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, L.J.; Lombardo, K.; Kwak, Y.; Kim, W.H.; Resnick, M.B. Expression of Indoleamine 2, 3-dioxygenase 1 (IDO1) and Tryptophanyl-tRNA Synthetase (WARS) in Gastric Cancer Molecular Subtypes. Appl. Immunohistochem. Mol. Morphol. 2020, 28, 360–368. [Google Scholar] [CrossRef]
- Ghanipour, A.; Jirstrom, K.; Ponten, F.; Glimelius, B.; Pahlman, L.; Birgisson, H. The prognostic significance of tryptophanyl-tRNA synthetase in colorectal cancer. Cancer Epidemiol. Biomark. Prev. 2009, 18, 2949–2956. [Google Scholar] [CrossRef] [Green Version]
- Chi, L.M.; Lee, C.W.; Chang, K.P.; Hao, S.P.; Lee, H.M.; Liang, Y.; Hsueh, C.; Yu, C.J.; Lee, I.N.; Chang, Y.J.; et al. Enhanced interferon signaling pathway in oral cancer revealed by quantitative proteome analysis of microdissected specimens using 16O/18O labeling and integrated two-dimensional LC-ESI-MALDI tandem MS. Mol. Cell Proteom. 2009, 8, 1453–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campone, M.; Valo, I.; Jezequel, P.; Moreau, M.; Boissard, A.; Campion, L.; Loussouarn, D.; Verriele, V.; Coqueret, O.; Guette, C. Prediction of Recurrence and Survival for Triple-Negative Breast Cancer (TNBC) by a Protein Signature in Tissue Samples. Mol. Cell Proteom. 2015, 14, 2936–2946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.; Mohammed, S. Breast cancer metastasis and the lymphatic system. Oncol. Lett. 2015, 10, 1233–1239. [Google Scholar] [CrossRef] [PubMed]
- Colleoni, M.; Rotmensz, N.; Maisonneuve, P.; Sonzogni, A.; Pruneri, G.; Casadio, C.; Luini, A.; Veronesi, P.; Intra, M.; Galimberti, V.; et al. Prognostic role of the extent of peritumoral vascular invasion in operable breast cancer. Ann. Oncol. 2007, 18, 1632–1640. [Google Scholar] [CrossRef]
- Hirakawa, S.; Brown, L.F.; Kodama, S.; Paavonen, K.; Alitalo, K.; Detmar, M. VEGF-C-induced lymphangiogenesis in sentinel lymph nodes promotes tumor metastasis to distant sites. Blood 2007, 109, 1010–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perl, D.P. Neuropathology of Alzheimer’s disease. Mt. Sinai J. Med. 2010, 77, 32–42. [Google Scholar] [CrossRef]
- Li, Y.; Nowotny, P.; Holmans, P.; Smemo, S.; Kauwe, J.S.; Hinrichs, A.L.; Tacey, K.; Doil, L.; van Luchene, R.; Garcia, V.; et al. Association of late-onset Alzheimer’s disease with genetic variation in multiple members of the GAPD gene family. Proc. Natl. Acad. Sci. USA 2004, 101, 15688–15693. [Google Scholar] [CrossRef] [Green Version]
- Sirover, M.A. On the functional diversity of glyceraldehyde-3-phosphate dehydrogenase: Biochemical mechanisms and regulatory control. Biochim. Biophys. Acta 2011, 1810, 741–751. [Google Scholar] [CrossRef]
- Tamaoka, A.; Endoh, R.; Shoji, S.; Takahashi, H.; Hirokawa, K.; Teplow, D.B.; Selkoe, D.J.; Mori, H. Antibodies to amyloid beta protein (A beta) crossreact with glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Neurobiol. Aging 1996, 17, 405–414. [Google Scholar] [CrossRef]
- Wang, Q.; Woltjer, R.L.; Cimino, P.J.; Pan, C.; Montine, K.S.; Zhang, J.; Montine, T.J. Proteomic analysis of neurofibrillary tangles in Alzheimer disease identifies GAPDH as a detergent-insoluble paired helical filament tau binding protein. FASEB J. 2005, 19, 869–871. [Google Scholar] [CrossRef]
- Cumming, R.C.; Schubert, D. Amyloid-beta induces disulfide bonding and aggregation of GAPDH in Alzheimer’s disease. FASEB J. 2005, 19, 2060–2062. [Google Scholar] [CrossRef] [PubMed]
- McGaha, T.L.; Huang, L.; Lemos, H.; Metz, R.; Mautino, M.; Prendergast, G.C.; Mellor, A.L. Amino acid catabolism: A pivotal regulator of innate and adaptive immunity. Immunol. Rev. 2012, 249, 135–157. [Google Scholar] [CrossRef] [Green Version]
- Zhai, L.; Bell, A.; Ladomersky, E.; Lauing, K.L.; Bollu, L.; Sosman, J.A.; Zhang, B.; Wu, J.D.; Miller, S.D.; Meeks, J.J.; et al. Immunosuppressive IDO in Cancer: Mechanisms of Action, Animal Models, and Targeting Strategies. Front. Immunol. 2020, 11, 1185. [Google Scholar] [CrossRef] [PubMed]
- Miyanokoshi, M.; Yokosawa, T.; Wakasugi, K. Tryptophanyl-tRNA synthetase mediates high-affinity tryptophan uptake into human cells. J. Biol. Chem. 2018, 293, 8428–8438. [Google Scholar] [CrossRef] [Green Version]
- Adam, I.; Dewi, D.L.; Mooiweer, J.; Sadik, A.; Mohapatra, S.R.; Berdel, B.; Keil, M.; Sonner, J.K.; Thedieck, K.; Rose, A.J.; et al. Upregulation of tryptophanyl-tRNA synthethase adapts human cancer cells to nutritional stress caused by tryptophan degradation. Oncoimmunology 2018, 7, e1486353. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, F.; Kuroiwa, T.; Takikawa, O.; Kido, R. Human indolylamine 2,3-dioxygenase. Its tissue distribution, and characterization of the placental enzyme. Biochem. J. 1985, 230, 635–638. [Google Scholar] [CrossRef]
- Platten, M.; Wick, W.; Van den Eynde, B.J. Tryptophan catabolism in cancer: Beyond IDO and tryptophan depletion. Cancer Res. 2012, 72, 5435–5440. [Google Scholar] [CrossRef] [Green Version]
- Sedlmayr, P.; Blaschitz, A.; Stocker, R. The role of placental tryptophan catabolism. Front. Immunol. 2014, 5, 230. [Google Scholar] [CrossRef] [Green Version]
- Boasso, A.; Herbeuval, J.P.; Hardy, A.W.; Winkler, C.; Shearer, G.M. Regulation of indoleamine 2,3-dioxygenase and tryptophanyl-tRNA-synthetase by CTLA-4-Fc in human CD4+ T cells. Blood 2005, 105, 1574–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Jun, L.; Shiyong, C.; Li, H.; Zhu, M.; Shen, B. Increased TTS expression in patients with rheumatoid arthritis. Clin. Exp. Med. 2015, 15, 25–30. [Google Scholar] [CrossRef]
- Zhu, L.; Ji, F.; Wang, Y.; Zhang, Y.; Liu, Q.; Zhang, J.Z.; Matsushima, K.; Cao, Q.; Zhang, Y. Synovial autoreactive T cells in rheumatoid arthritis resist IDO-mediated inhibition. J. Immunol. 2006, 177, 8226–8233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massalska, M.; Kuca-Warnawin, E.; Janicka, I.; Plebanczyk, M.; Pawlak, D.; Dallos, T.; Olwert, A.; Radzikowska, A.; Maldyk, P.; Kontny, E.; et al. Survival of lymphocytes is not restricted by IDO-expressing fibroblast from rheumatoid arthritis patients. Immunopharmacol. Immunotoxicol. 2019, 41, 214–223. [Google Scholar] [CrossRef]
- Wang, C.Y.; Shi, Y.; Min, Y.N.; Zhu, X.J.; Guo, C.S.; Peng, J.; Dong, X.Y.; Qin, P.; Sun, J.Z.; Hou, M. Decreased IDO activity and increased TTS expression break immune tolerance in patients with immune thrombocytopenia. J. Clin. Immunol. 2011, 31, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Mao, C.; Zhao, Z.; Gu, Q.; Jin, M.; Xiao, Y.; Jiang, X.; Zhao, Y.; Zhang, Y.; Ning, G. Increased TTS abrogates IDO-mediated CD4(+) T cells suppression in patients with Graves’ disease. Endocrine 2009, 36, 119–125. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| ARSs | Schematic Structure | Alternative Variants | Proteolytic Cleavage | References | |
|---|---|---|---|---|---|
| MCS form | EPRS | 1  1512 1512 | EPRSN1 | [17,18,35,36] | |
| MRS | 1  900 900 | [37,38,39] | |||
| Free form | WRS | 1  471 471 | mini-WRS | T1-WRS T2-WRS | [6,13,20,22,27,28,30,31,32] |
| HRS | 1  509 509 | HRSΔC | [40,41,42] | ||
| GRS | 1  739 739 | [43,44] |

| WRS Variants | Schematic Structure | Aminoacylation Activity | Angiostatic Activity |
|---|---|---|---|
| FL-WRS | 1  471 471 | + | − |
| mini-WRS | 48  471 471 | + | + |
| T1-WRS | 71  471 471 | + | + |
| T2-WRS | 94  471 471 | − | ++ |

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, Y.H.; Oh, S.-C.; Zhou, S.; Kim, T.-D. Tryptophanyl-tRNA Synthetase as a Potential Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 4523. https://doi.org/10.3390/ijms22094523
Ahn YH, Oh S-C, Zhou S, Kim T-D. Tryptophanyl-tRNA Synthetase as a Potential Therapeutic Target. International Journal of Molecular Sciences. 2021; 22(9):4523. https://doi.org/10.3390/ijms22094523
Chicago/Turabian StyleAhn, Young Ha, Se-Chan Oh, Shengtao Zhou, and Tae-Don Kim. 2021. "Tryptophanyl-tRNA Synthetase as a Potential Therapeutic Target" International Journal of Molecular Sciences 22, no. 9: 4523. https://doi.org/10.3390/ijms22094523
APA StyleAhn, Y. H., Oh, S. -C., Zhou, S., & Kim, T. -D. (2021). Tryptophanyl-tRNA Synthetase as a Potential Therapeutic Target. International Journal of Molecular Sciences, 22(9), 4523. https://doi.org/10.3390/ijms22094523