
Modulation of Diverse Procoagulant Venom Activities by Combinations of Platinoid Compounds


Abstract
:1. Introduction
2. Results
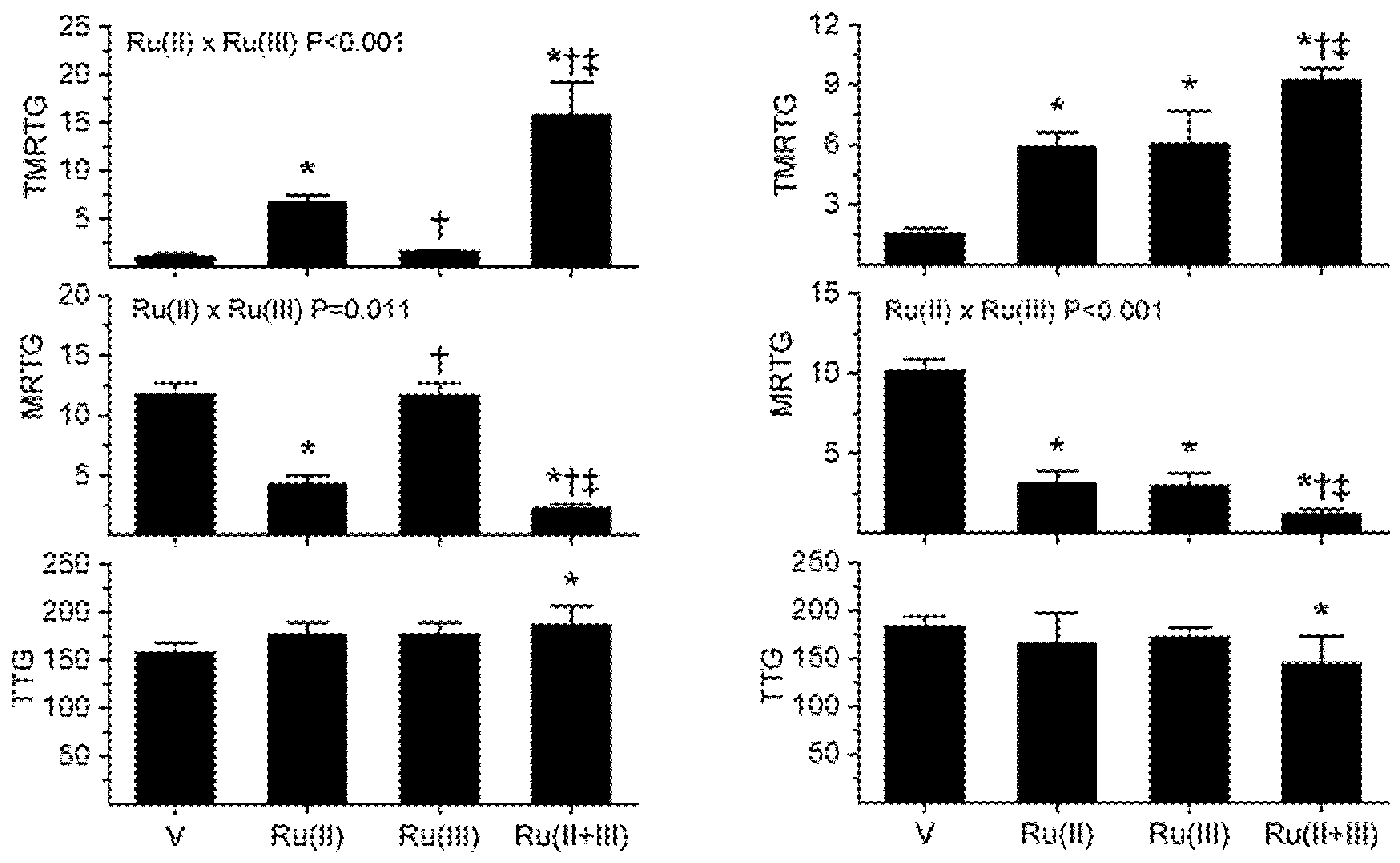
2.1. Assessment of CORM-2 and RuCl3, Separately and in Combination, on the Procoagulant Activity of B. moojeni, C. rhodostoma, E. leucogaster and O. microlepidotus
2.2. Assessment of CORM-3 and RuCl3, Separately and in Combination, on the Procoagulant Activity of B. moojeni, C. rhodostoma, P. textilis and H. suspectum
2.3. Assessment of CORM-2 and Carboplatin, Separately and in Combination, on the Procoagulant Activity of B. moojeni and C. rhodostoma Venoms
3. Discussion
4. Materials and Methods
4.1. Chemicals and Human Plasma
4.2. Thrombelastographic Analyses
4.3. Exposures of Venoms to CORM-2, CORM-3, RuCl3 and Carboplatin
4.4. Statistical Analyses and Graphics
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nielsen, V.G.; Bazzell, C.M. Carbon monoxide releasing molecule-2 inhibition of snake venom thrombin-like activity: Novel biochemical “brake”? J. Thromb. Thrombolysis 2017, 43, 203–208. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Frank, N.; Matika, R.W. Carbon monoxide inhibits hemotoxic activity of Elapidae venoms: Potential role of heme. BioMetals 2018, 31, 51–59. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Frank, N. Role of heme modulation in inhibition of Atheris, Atractaspis, Causus, Cerastes, Echis, and Macrovipera hemotoxic venom activity. Hum. Exp. Toxicol. 2019, 38, 216–226. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Frank, N.; Afshar, S. De Novo Assessment and Review of Pan-American Pit Viper Anticoagulant and Procoagulant Venom Activities via Kinetomic Analyses. Toxins 2019, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Suntravat, M.; Langlais, P.R.; Sánchez, E.E.; Nielsen, V.G. CatroxMP-II: A heme-modulated fibrinogenolytic metalloproteinase isolated from Crotalus atrox venom. BioMetals 2018, 31, 585–593. [Google Scholar] [CrossRef]
- Nielsen, V.G. Carbon monoxide inhibits the anticoagulant activity of phospholipase A2 purified from Crotalus adamanteus venom. J. Thromb. Thrombolysis 2019, 47, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Gessner, G.; Sahoo, N.; Swain, S.M.; Hirth, G.; Schönherr, R.; Mede, R.; Westerhausen, M.; Brewitz, H.H.; Heimer, P.; Imhof, D.; et al. CO-independent modification of K+ channels by tricarbonyldichlororuthenium(II) dimer (CORM-2). Eur. J. Pharmacol. 2017, 815, 33–41. [Google Scholar] [CrossRef]
- Southam, H.M.; Smith, T.W.; Lyon, R.L.; Liao, C.; Trevitt, C.R.; Middlemiss, L.A.; Cox, F.L.; Chapman, J.A.; El-Khamisy, S.F.; Hippler, M.; et al. A thiol-reactive Ru(II) ion, not CO release, underlies the potent antimicrobial and cytotoxic properties of CO-releasing molecule-3. Redox Biol. 2018, 18, 114–123. [Google Scholar] [CrossRef]
- Nielsen, V.G. The anticoagulant effect of Apis mellifera phospholipase A2 is inhibited by CORM-2 via a carbon monoxide-independent mechanism. J. Thromb Thrombolysis 2020, 49, 100–107. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Wagner, M.T.; Frank, N. Mechanisms Responsible for the Anticoagulant Properties of Neurotoxic Dendroaspis Venoms: A Viscoelastic Analysis. Int. J. Mol. Sci. 2020, 21, 2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, V.G. Ruthenium, Not Carbon Monoxide, Inhibits the Procoagulant Activity of Atheris, Echis, and Pseudonaja Venoms. Int. J. Mol. Sci 2020, 21, 2970. [Google Scholar] [CrossRef]
- Lazić, D.; Arsenijević, A.; Puchta, R.; Bugarčić, Ž.D.; Rilak, A. DNA binding properties, histidine interaction and cytotoxicity studies of water soluble ruthenium(II) terpyridine complexes. Dalton Trans. 2016, 45, 4633. [Google Scholar] [CrossRef] [Green Version]
- Hanif, M.; Meier, S.M.; Adhireksan, Z.; Henke, H.; Martic, S.; Movassaghi, S.; Labib, M.; Kandioller, W.; Jamieson, S.M.F.; Hejl, M.; et al. Functionalization of Ruthenium(II)(η6-p-cymene)(3-hydroxy-2-pyridone) Complexes with (Thio) Morpholine: Synthesis and Bioanalytical Studies. ChemPlusChem 2017, 82, 841–847. [Google Scholar] [CrossRef]
- Stanic-Vucinic, D.; Nikolic, S.; Vlajic, K.; Radomirovic, M.; Mihailovic, J.; Velickovic, T.C.; Grguric-Sipka, S. The interactions of the ruthenium(II)-cymene complexes with lysozyme and cytochrome c. J. Biol. Inorg. Chem. 2020, 25, 253–265. [Google Scholar] [CrossRef]
- Yocom, K.M.; Shelton, J.B.; Shelton, J.R.; Schroeder, W.A.; Worosila, G.; Isied, S.S.; Bordignon, E.; Gray, H.B. Preparation and characterization of a pentaammineruthenium(III) derivative of horse heart ferricytochrome c. Proc. Natl. Acad. Sci. USA 1982, 79, 7052–7055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratz, K.; Keppler, B.K.; Messori, L.; Smith, C.; Baker, E.N. Protein-binding Properties of two Antitumour Ru(III) Complexes to Human Apotransferrin and Apolactoferrin. Met. Based Drugs 1994, 1, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Webb, M.I.; Walsby, C.J. Albumin binding and ligand-exchange processes of the Ru(III) anticancer agent NAMI-A and its bis-DMSO analogue determined by ENDOR spectroscopy. Dalton Trans. 2015, 44, 17482. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Bobst, C.E.; Kaltashov, I.A. Exploiting His-Tags for Absolute Quantitation of Exogenous Recombinant Proteins in Biological Matrices: Ruthenium as a Protein Tracer. Anal. Chem. 2019, 91, 7189–7198. [Google Scholar] [CrossRef]
- Das, D.; Dutta, S.; Dowerah, D.; Deka, R.C. Unveiling the Role of Hydrogen Bonding and g-Tensor in the Interaction of Ru-Bis-DMSO with Amino Acid Residue and Human Serum Albumin. J. Phys. Chem. B 2020, 124, 6459–6474. [Google Scholar] [CrossRef] [PubMed]
- Artiaga, G.; Iglesias-Jiménez, A.; Moreno-Gordaliza, E.; Mena, M.L.; Gómez-Gómez, M.M. Differences in binding kinetics, bond strength and adduct formation between Pt-based drugs and S- or N-donor groups: A comparative study using mass spectrometry techniques Eur. J. Pharm. Sci. 2019, 132, 96–105. [Google Scholar] [CrossRef]
- Watanabe, L.; Shannon, J.D.; Valente, R.H.; Rucavado, A.; Alape-Girón, A.; Kamiguti, A.S.; Theakston, R.D.G.; Fox, J.W.; Gutiérrez, J.M.; Arni, R.K. Amino acid sequence and crystal structure of BaP1, a metalloproteinase from Bothrops asper snake venom that exerts multiple tissue-damaging activities. Protein Sci. 2003, 12, 2273–2281. [Google Scholar] [CrossRef] [PubMed]
- Markland, F.S., Jr.; Swenson, S. Snake venom metalloproteinases. Toxicon 2013, 62, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Schrader, M.; Raida, M.; McLane, M.A.; Romero, A.; Niewiarowski, S. The disulphide bond pattern of bitistatin, a disintegrin isolated from the venom of the viper Bitis arietans. FEBS Lett. 1997, 416, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Braud, S.; Parry, M.A.; Maroun, R.; Bon, C.; Wisner, A. The contribution of residues 192 and 193 to the specificity of snake venom serine proteinases. J. Biol. Chem. 2000, 275, 1823–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentin, E.; Lambeau, G. What can venom phospholipases A(2) tell us about the functional diversity of mammalian secreted phospholipases A(2)? Biochimie 2000, 82, 815–831. [Google Scholar] [CrossRef]
- Soares, A.M.; Giglio, J.R. Chemical modifications of phospholipases A2 from snake venoms: Effects on catalytic and pharmacological properties. Toxicon 2003, 42, 855–868. [Google Scholar] [CrossRef]
- Aguiar, W.D.S.; Galizio, N.D.C.; Serino-Silva, C.; Sant’Anna, S.S.; Grego, K.F.; Tashima, A.K.; Nishiduka, E.S.; Morais-Zani, K.; Tanaka-Azevedo, A.M. Comparative compositional and functional analyses of Bothrops moojeni specimens reveal several individual variations. PLoS ONE 2019, 14, e0222206. [Google Scholar] [CrossRef] [PubMed]
- Tang, E.L.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Venomics of Calloselasma rhodostoma, the Malayan pit viper: A complex toxin arsenal unraveled. J. Proteom. 2016, 148, 44–56. [Google Scholar] [CrossRef]
- Patra, A.; Kalita, B.; Chanda, A.; Mukherjee, A.K. Proteomics and antivenomics of Echis carinatus carinatus venom: Correlation with pharmacological properties and pathophysiology of envenomation. Sci. Rep. 2017, 7, 17119. [Google Scholar] [CrossRef] [Green Version]
- Yamada, D.; Morita, T. CA-1 method, a novel assay for quantification of normal prothrombin using a Ca2+-dependent prothrombin activator, carinactivase-1. Thromb. Res. 1999, 94, 221–226. [Google Scholar] [CrossRef]
- Chen, Y.L.; Tsai, I.H. Functional and sequence characterization of coagulation factor IX/factor X-binding protein from the venom of Echis carinatus leucogaster. Biochemistry 1996, 35, 5264–52671. [Google Scholar] [CrossRef]
- Koludarov, I.; Jackson, T.N.; Sunagar, K.; Nouwens, A.; Hendrikx, I.; Fry, B.G. Fossilized venom: The unusually conserved venom profiles of Heloderma species (beaded lizards and gila monsters). Toxins 2014, 6, 3582–3595. [Google Scholar] [CrossRef] [Green Version]
- Sanggaard, K.W.; Dyrlund, T.F.; Thomsen, L.R.; Nielsen, T.A.; Brøndum, L.; Wang, T.; Thøgersen, I.B.; Enghild, J.J. Characterization of the gila monster (Heloderma suspectum suspectum) venom proteome. J. Proteom. 2015, 117, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Herrera, M.; Fernández, J.; Vargas, M.; Villalta, M.; Segura, Á.; León, G.; Angulo, Y.; Paiva, O.; Matainaho, T.; Jensen, S.D.; et al. Comparative proteomic analysis of the venom of the taipan snake, Oxyuranus scutellatus, from Papua New Guinea and Australia: Role of neurotoxic and procoagulant effects in venom toxicity. J. Proteom. 2012, 75, 2128–2140. [Google Scholar] [CrossRef]
- McCleary, R.J.; Sridharan, S.; Dunstan, N.L.; Mirtschin, P.J.; Kini, R.M. Proteomic comparisons of venoms of long-term captive and recently wild-caught Eastern brown snakes (Pseudonaja textilis) indicate venom does not change due to captivity. J. Proteom. 2016, 144, 51–62. [Google Scholar] [CrossRef]
- Nielsen, V.G.; Frank, N. The kallikrein-like activity of Heloderma venom is inhibited by carbon monoxide. J. Thromb. Thrombolysis 2019, 47, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G. Platinoid effects on human plasmatic coagulation kinetics: A viscoelastic analysis. J. Thromb. Thrombolysis 2021. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, V.G.; Chawla, N.; Mangla, D.; Gomes, S.B.; Arkebauer, M.R.; Wasko, K.A.; Sadacharam, K.; Vosseller, K. Carbon monoxide-releasing molecule-2 enhances coagulation in rabbit plasma and decreases bleeding time in clopidogrel/aspirin-treated rabbits. Blood Coagul. Fibrinolysis 2011, 22, 756–759. [Google Scholar] [CrossRef] [PubMed]
- Stagni, E.; Privitera, M.G.; Bucolo, C.; Leggio, G.M.; Motterlini, R.; Drago, F. A water-soluble carbon monoxide-releasing molecule (CORM-3) lowers intraocular pressure in rabbits. Br. J. Ophthalmol. 2009, 93, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, Y.; Zhu, J.; Ma, J.; Niu, Q.; Wang, X. Carbon monoxide attenuates LPS-induced myocardial dysfunction in rats by regulating the mitochondrial dynamic equilibrium. Eur. J. Pharm. 2020, 889, 173726. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Bai, Y.; Wang, X.P.; Zheng, W.C.; Zhang, L.M. Carbon Monoxide-Releasing Molecule-3 Ameliorates Acute Lung Injury in a Model of Hemorrhagic Shock and Resuscitation: Roles of p38MAPK Signaling Pathway. Shock 2020. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Proteome |
|---|---|---|
| Bothrops moojeni [27] | Brazilian Lancehead | SP, MP |
| Calloselasma rhodostoma [28] | Malayan Pit Viper | SP, MP |
| Echis leucogaster [29,30,31] | White-Bellied Carpet Viper | SP, MP |
| Heloderma suspectum [32,33] | Gila Monster | Kallikrein-like SP |
| Oxyuranus microlepidotus [34] | Inland Taipan | Factor V-like, SP, MP |
| Pseudonaja textilis [35] | Eastern Brown Snake | Factor V, X-like SP, MP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, V.G. Modulation of Diverse Procoagulant Venom Activities by Combinations of Platinoid Compounds. Int. J. Mol. Sci. 2021, 22, 4612. https://doi.org/10.3390/ijms22094612
Nielsen VG. Modulation of Diverse Procoagulant Venom Activities by Combinations of Platinoid Compounds. International Journal of Molecular Sciences. 2021; 22(9):4612. https://doi.org/10.3390/ijms22094612
Chicago/Turabian StyleNielsen, Vance G. 2021. "Modulation of Diverse Procoagulant Venom Activities by Combinations of Platinoid Compounds" International Journal of Molecular Sciences 22, no. 9: 4612. https://doi.org/10.3390/ijms22094612
APA StyleNielsen, V. G. (2021). Modulation of Diverse Procoagulant Venom Activities by Combinations of Platinoid Compounds. International Journal of Molecular Sciences, 22(9), 4612. https://doi.org/10.3390/ijms22094612