
Fibroinflammatory Signatures Increase with Age in the Human Ovary and Follicular Fluid


 ,
,
Abstract
1. Introduction
2. Results
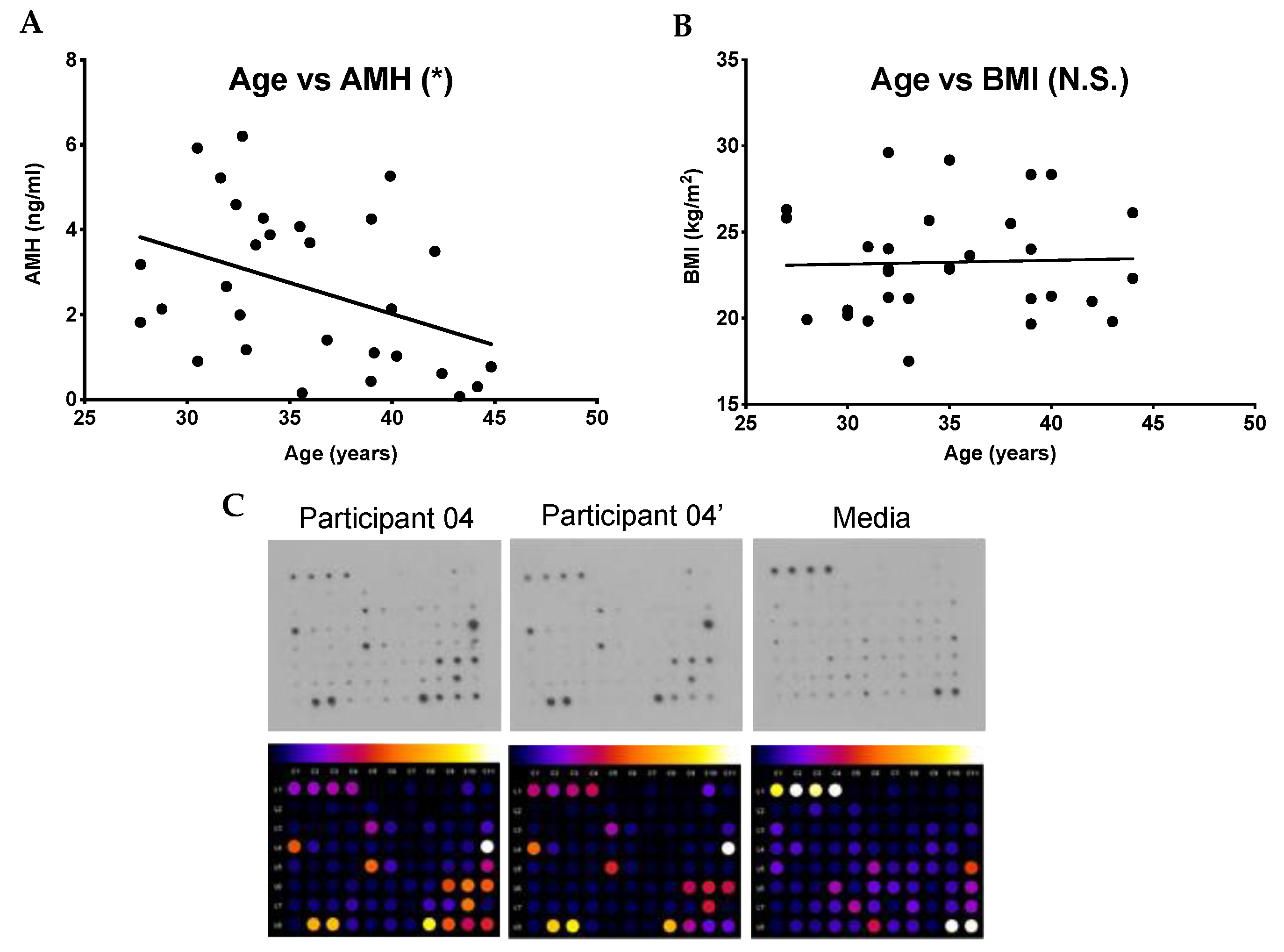
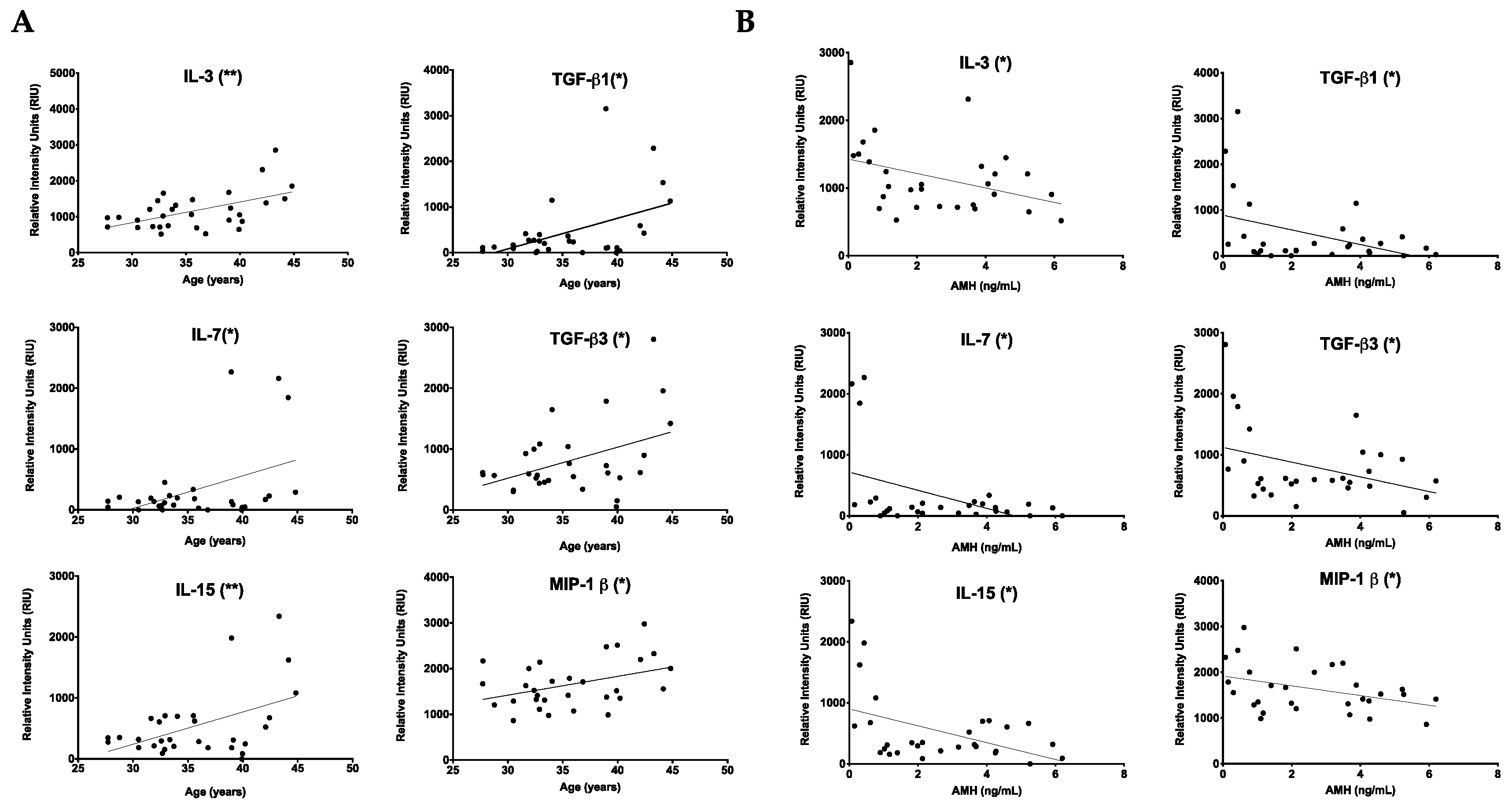
2.1. Chronologic and Reproductive Age Are Associated with an Increase in Specific Subsets of Fibroinflammatory Cytokines
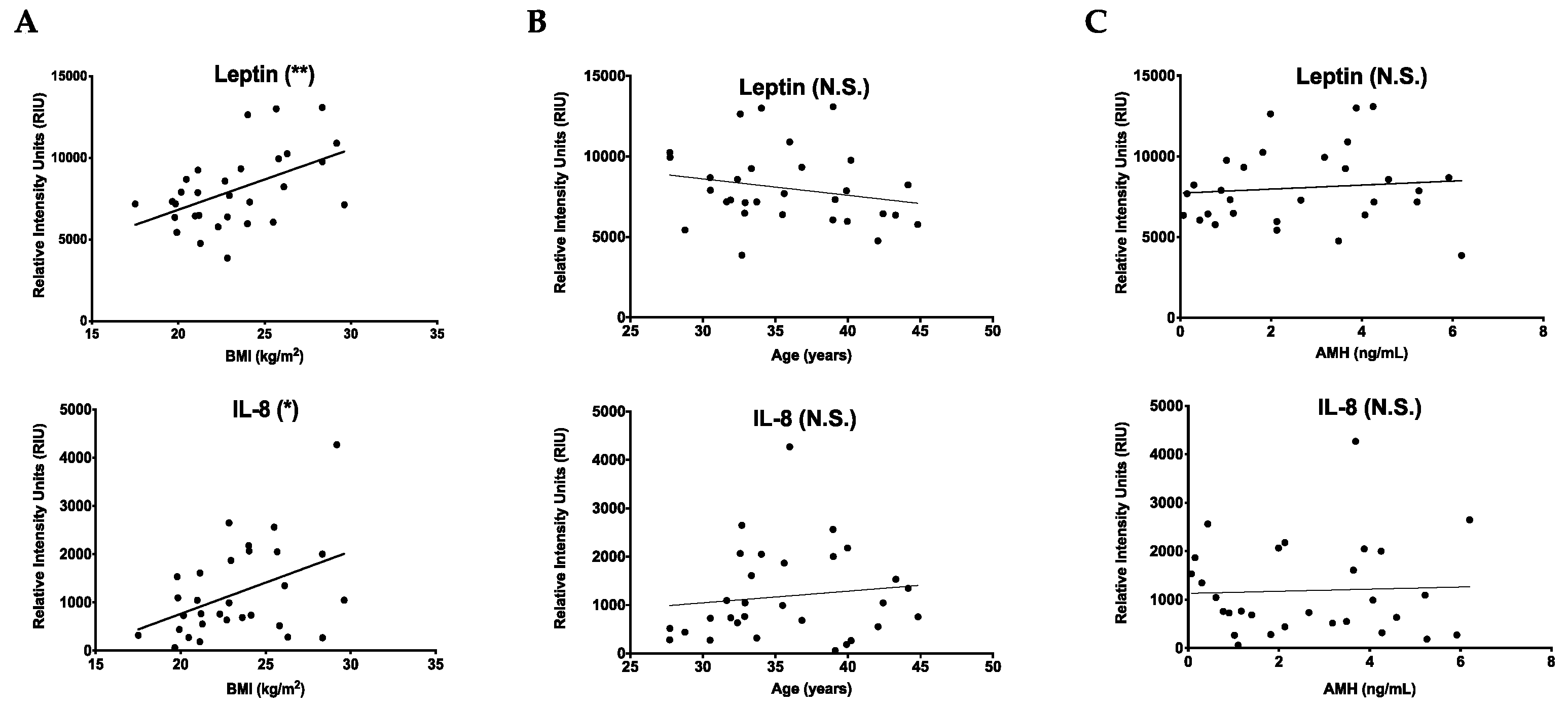
2.2. Cytokines That Increase with BMI Are Unique from Those That Change with Age
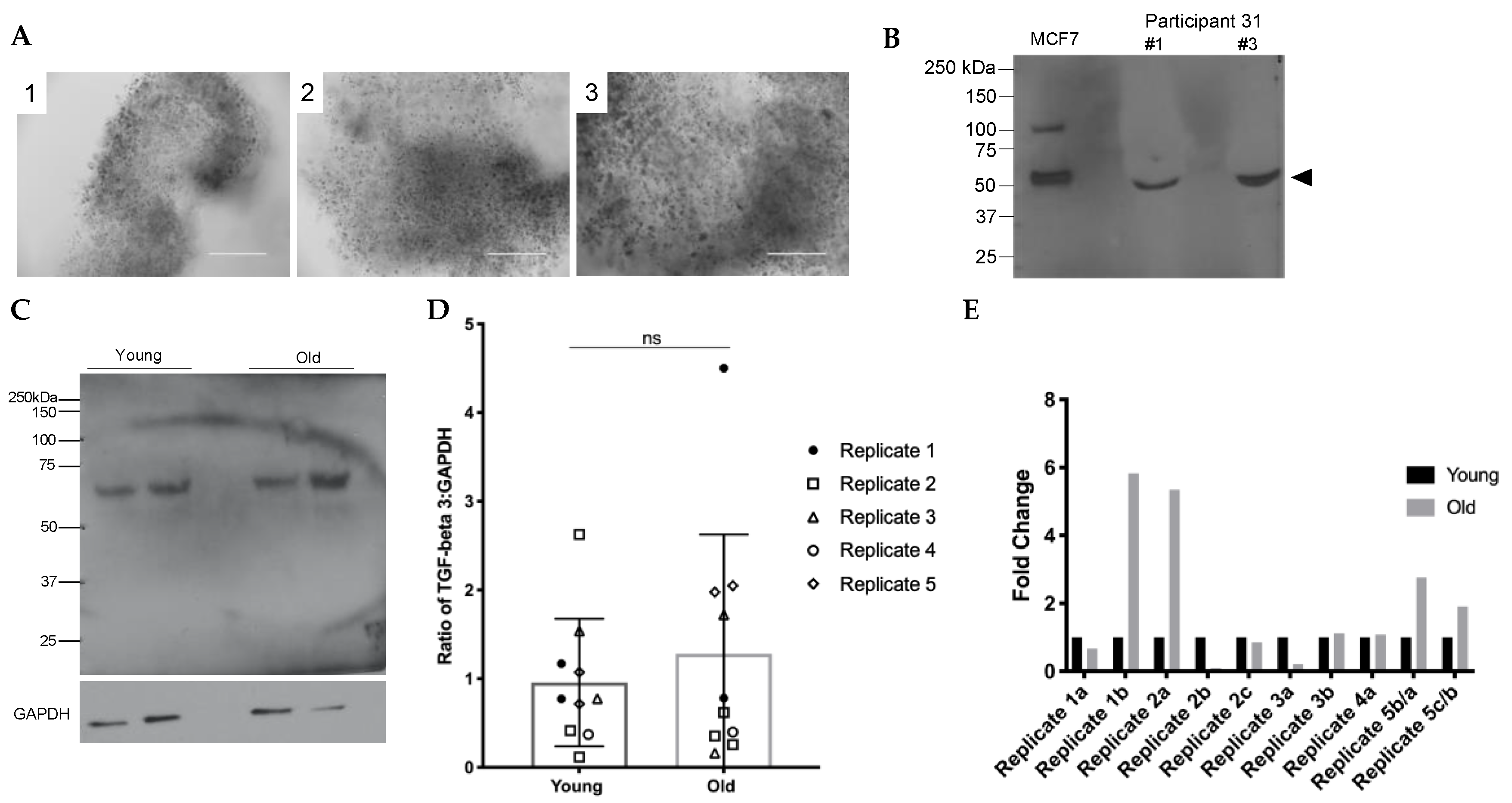
2.3. TGFβ3 Content in Human Cumulus Cells Does Not Increase with Age
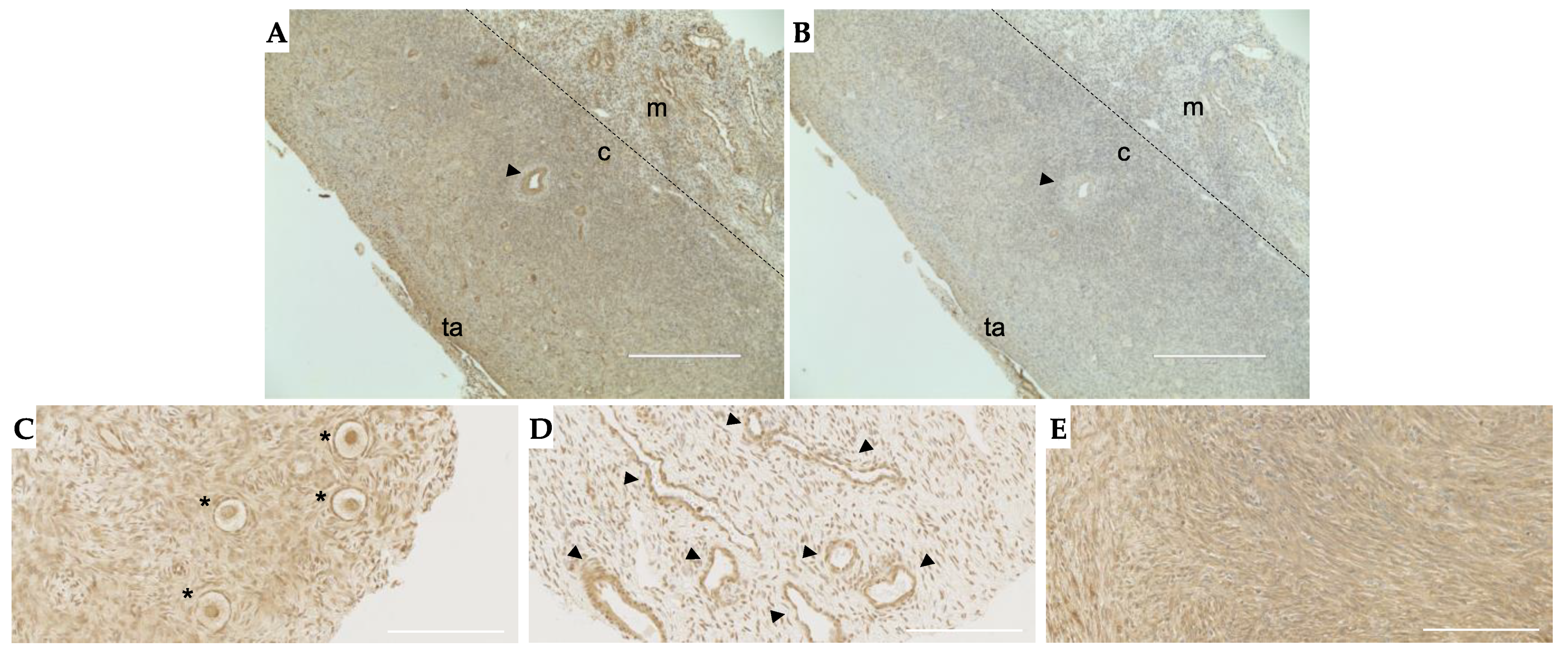
2.4. TGFβ3 Is Expressed in Follicles, Vasculature and Stroma in the Human Ovary
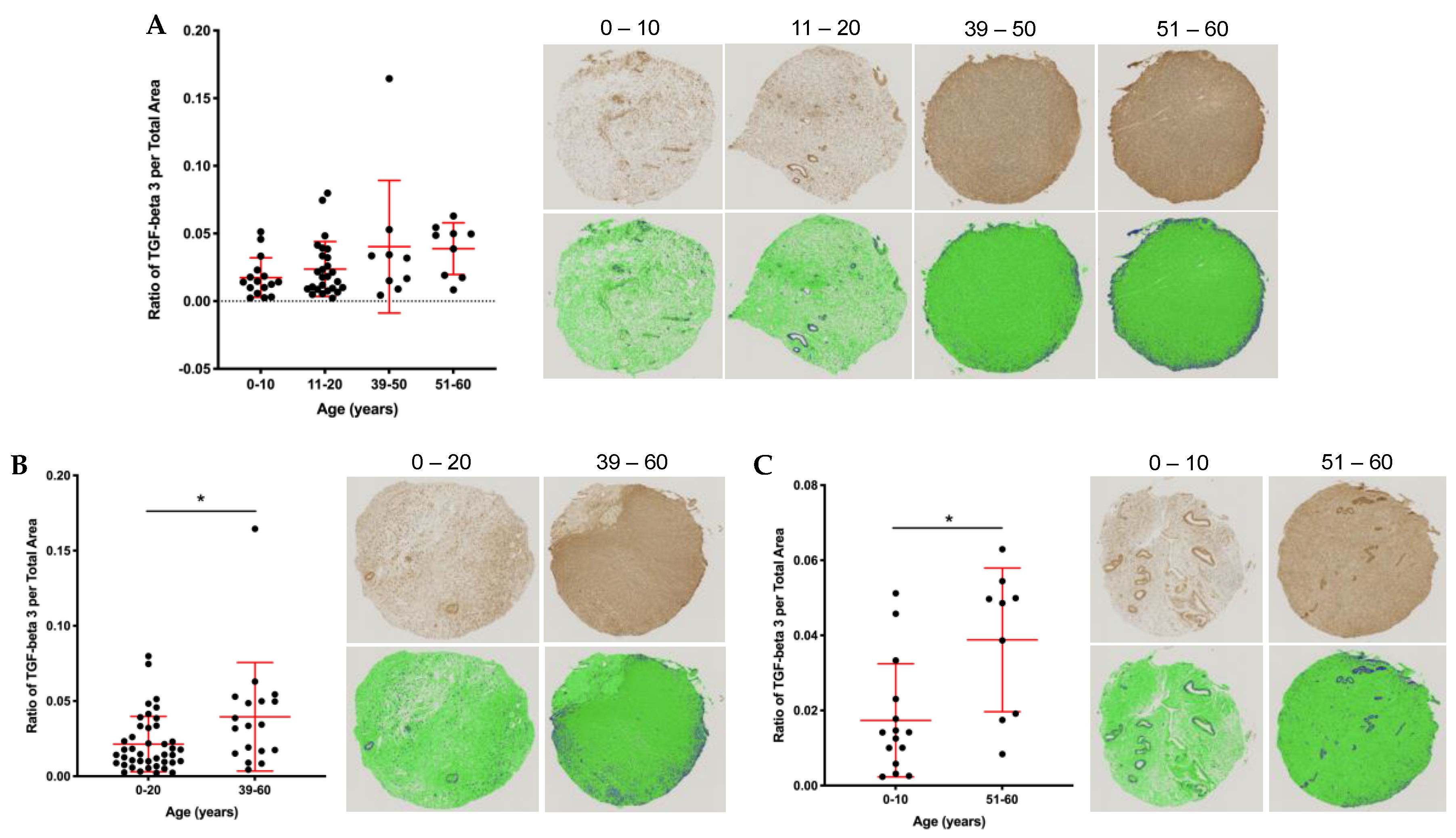
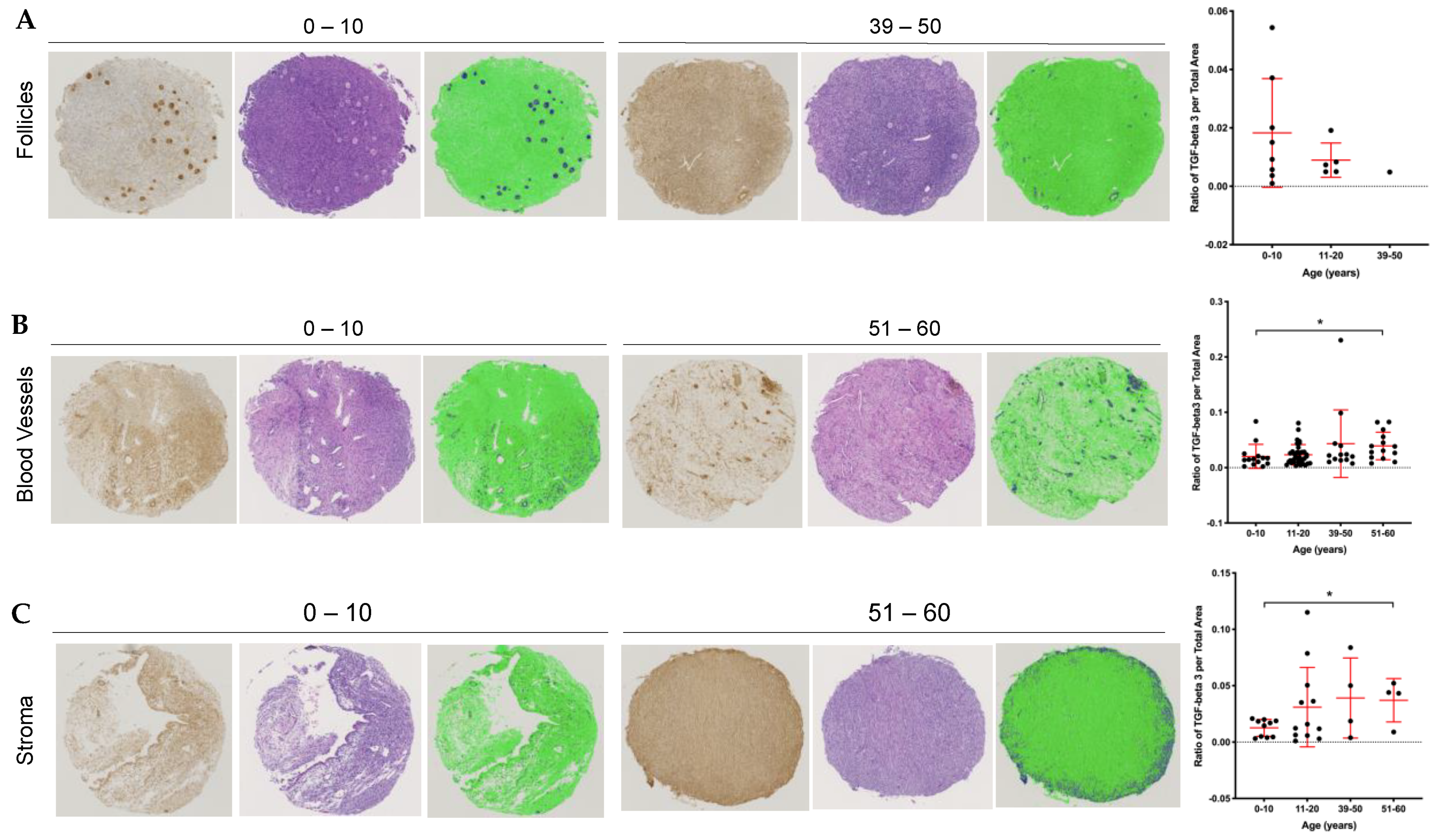
2.5. Stromal and Vascular TGFβ3 Content Increases with Age in the Human Ovary
3. Discussion
4. Materials and Methods
4.1. Human Follicular Fluid, Cumulus Cells, and Ovarian Tissue
4.2. Cytokine Antibody Arrays
4.3. Antibodies
4.4. Protein Extraction and Immunoblot Analysis (Non-Reducing Conditions)
4.5. Immunohistochemistry (IHC)
4.6. Human Ovarian Tissue Microarray (TMA) TGFβ3 Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Navarro-Costa, P. A (micro)environmental perspective on the evolution of female reproductive aging. J. Assist. Reprod. Genet. 2018, 35, 2129–2131. [Google Scholar] [CrossRef]
- Hall, J.E. Endocrinology of the Menopause. Endocrinol. Metab. Clin. N. Am. 2015, 44, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, H.; Takahashi, T.; Nagase, S. Oocyte aging underlies female reproductive aging: Biological mechanisms and therapeutic strategies. Reprod. Med. Biol. 2015, 14, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Cimadomo, D.; Fabozzi, G.; Vaiarelli, A.; Ubaldi, N.; Ubaldi, F.M.; Rienzi, L. Impact of Maternal Age on Oocyte and Embryo Competence. Front. Endocrinol. 2018, 9, 327. [Google Scholar] [CrossRef] [PubMed]
- Ameratunga, D.; Weston, G.; Osianlis, T.; Catt, J.; Vollenhoven, B. In vitro fertilisation (IVF) with donor eggs in post-menopausal women: Are there differences in pregnancy outcomes in women with premature ovarian failure (POF) compared with women with physiological age-related menopause? J. Assist. Reprod. Genet. 2009, 26, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Ma, J.; Schatten, H.; Sun, Q. Epigenetic changes associated with oocyte aging. Sci. China Life Sci. 2012, 55, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Eichenlaub-Ritter, U.; Wieczorek, M.; Lüke, S.; Seidel, T. Age related changes in mitochondrial function and new approaches to study redox regulation in mammalian oocytes in response to age or maturation conditions. Mitochondrion 2011, 11, 783–796. [Google Scholar] [CrossRef]
- Hornick, J.E.; Duncan, F.E.; Sun, M.; Kawamura, R.; Marko, J.F.; Woodruff, T.K. Age-associated alterations in the micromechanical properties of chromosomes in the mammalian egg. J. Assist. Reprod. Genet. 2015, 32, 765–769. [Google Scholar] [CrossRef]
- Duncan, F.E.; Jasti, S.; Paulson, A.; Kelsh, J.M.; Fegley, B.; Gerton, J.L. Age-associated dysregulation of protein metabolism in the mammalian oocyte. Aging Cell 2017, 16, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Thannickal, V.J.; Murthy, M.; Balch, W.E.; Chandel, N.S.; Meiners, S.; Eickelberg, O.; Selman, M.; Pardo, A.; White, E.S.; Levy, B.D.; et al. Blue Journal Conference. Aging and Susceptibility to Lung Disease. Am. J. Respir. Crit. Care Med. 2015, 191, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Weiss, U. Inflammation. Nature 2008, 454, 427. [Google Scholar] [CrossRef]
- Nathan, C.; Ding, A. Nonresolving Inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef]
- Sahin, H.; Wasmuth, H.E. Chemokines in tissue fibrosis. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 1041–1048. [Google Scholar] [CrossRef]
- Briley, S.M.; Jasti, S.; McCracken, J.M.; Hornick, J.E.; Fegley, B.; Pritchard, M.T.; Duncan, F.E. Reproductive age-associated fibrosis in the stroma of the mammalian ovary. Reproduction 2016, 152, 245–260. [Google Scholar] [CrossRef]
- Amargant, F.; Manuel, S.L.; Tu, Q.; Parkes, W.S.; Rivas, F.; Zhou, L.T.; Rowley, J.E.; Villanueva, C.E.; Hornick, J.E.; Shekhawat, G.S.; et al. Ovarian stiffness increases with age in the mammalian ovary and depends on collagen and hyaluronan matrices. Aging Cell 2020, 19, e13259. [Google Scholar] [CrossRef] [PubMed]
- Foley, K.G.; Pritchard, M.T.; Duncan, F.E. Macrophage-derived multinucleated giant cells: Hallmarks of the aging ovary. Reproduction 2021, 161, V5–V9. [Google Scholar] [CrossRef]
- Huang, Y.; Hu, C.; Ye, H.; Luo, R.; Fu, X.; Li, X.; Huang, J.; Chen, W.; Zheng, Y. Inflamm-Aging: A New Mechanism Affecting Premature Ovarian Insufficiency. J. Immunol. Res. 2019, 2019, 8069898. [Google Scholar] [CrossRef]
- Umehara, T.; Richards, J.S.; Shimada, M. The stromal fibrosis in aging ovary. Aging 2018, 10, 9–10. [Google Scholar] [CrossRef]
- Edwards, R. Follicular Fluid. Reproduction 1974, 37, 189–219. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, R.J.; Irving-Rodgers, H.F. Formation of the Ovarian Follicular Antrum and Follicular Fluid1. Biol. Reprod. 2010, 82, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Revelli, A.; Piane, L.D.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular fluid content and oocyte quality: From single biochemical markers to metabolomics. Reprod. Biol. Endocrinol. 2009, 7, 40. [Google Scholar] [CrossRef] [PubMed]
- Emori, M.M.; Drapkin, R. The hormonal composition of follicular fluid and its implications for ovarian cancer pathogenesis. Reprod. Biol. Endocrinol. 2014, 12, 60. [Google Scholar] [CrossRef]
- Bouet, P.-E.; Boueilh, T.; de la Barca, J.M.C.; Boucret, L.; Blanchard, S.; Ferré-L’Hotellier, V.; Jeannin, P.; Descamps, P.; Procaccio, V.; Reynier, P.; et al. The cytokine profile of follicular fluid changes during ovarian ageing. J. Gynecol. Obstet. Hum. Reprod. 2020, 49, 101704. [Google Scholar] [CrossRef]
- de Vet, A.; Laven, J.S.; de Jong, F.H.; Themmen, A.P.; Fauser, B.C. Antimüllerian hormone serum levels: A putative marker for ovarian aging. Fertil. Steril. 2002, 77, 357–362. [Google Scholar] [CrossRef]
- Bentzen, J.G.; Forman, J.L.; Johannsen, T.H.; Pinborg, A.; Larsen, E.C.; Andersen, A.N. Ovarian Antral Follicle Subclasses and Anti-Müllerian Hormone During Normal Reproductive Aging. J. Clin. Endocrinol. Metab. 2013, 98, 1602–1611. [Google Scholar] [CrossRef] [PubMed]
- Van Rooij, I.; Broekmans, F.; Velde, E.T.; Fauser, B.; Bancsi, L.; Jong, F.; Themmen, A. Serum anti-Mullerian hormone levels: A novel measure of ovarian reserve. Hum. Reprod. 2002, 17, 3065–3071. [Google Scholar] [CrossRef]
- Welt, C.K.; Schneyer, A.L.; Heist, K.; Mantzoros, C.S. Leptin and Soluble Leptin Receptor in Follicular Fluid. J. Assist. Reprod. Genet. 2003, 20, 495–501. [Google Scholar] [CrossRef]
- Llaneza-Suarez, D.; Llaneza, P.; González, C.; De-La-Fuente, P.; García-Ochoa, C.; Garrido, P.; Castañón, V.; Pérez-López, F.R. Assessment of follicular fluid leptin levels and insulin resistance as outcome predictors in women undergoing in vitro fertilization–intracytoplasmic sperm injection. Fertil. Steril. 2014, 102, 1619–1625. [Google Scholar] [CrossRef]
- Letterio, J.J.; Roberts, A.B. Regulation of Immune Responses by TGF-β. Annu. Rev. Immunol. 1998, 16, 137–161. [Google Scholar] [CrossRef]
- Border, W.A.; Ruoslahti, E. Transforming growth factor-beta in disease: The dark side of tissue repair. J. Clin. Investig. 1992, 90, 1–7. [Google Scholar] [CrossRef]
- Raja-Khan, N.; Urbanek, M.; Rodgers, R.J.; Legro, R.S. The role of TGF-beta in polycystic ovary syndrome. Reprod. Sci. 2014, 21, 20–31. [Google Scholar] [CrossRef]
- Dewidar, B.; Meyer, C.; Dooley, S.; Meindl-Beinker, A.N. TGF-β in Hepatic Stellate Cell Activation and Liver Fibrogenesis—Updated 2019. Cells 2019, 8, 1419. [Google Scholar] [CrossRef]
- Monsivais, D.; Matzuk, M.M.; Pangas, S.A. The TGF-beta Family in the Reproductive Tract. Cold Spring Harb. Perspect Biol. 2017, 9, 10. [Google Scholar] [CrossRef]
- Hachim, M.Y.; Hachim, I.Y.; Dai, M.; Ali, S.; Lebrun, J.J. Differential expression of TGFbeta isoforms in breast cancer highlights different roles during breast cancer progression. Tumour Biol. 2018, 40, 1010428317748254. [Google Scholar] [CrossRef] [PubMed]
- Janny, L.; Menezo, Y.J.R. Maternal age effect on early human embryonic development and blastocyst formation. Mol. Reprod. Dev. 1996, 45, 31–37. [Google Scholar] [CrossRef]
- Steuerwald, N.M.; Bermúdez, M.G.; Wells, D.; Munné, S.; Cohen, J. Maternal age-related differential global expression profiles observed in human oocytes. Reprod. Biomed. Online 2007, 14, 700–708. [Google Scholar] [CrossRef]
- Munné, S.; Sandalinas, M.; Escudero, T.; Márquez, C.; Cohen, J. Chromosome mosaicism in cleavage-stage human embryos: Evidence of a maternal age effect. Reprod. Biomed. Online 2002, 4, 223–232. [Google Scholar] [CrossRef]
- Hou, Y.-J.; Zhu, C.-C.; Duan, X.; Liu, H.-L.; Wang, Q.; Sun, S.-C. Both diet and gene mutation induced obesity affect oocyte quality in mice. Sci. Rep. 2016, 6, 18858. [Google Scholar] [CrossRef] [PubMed]
- Sneed, M.L.; Uhler, M.L.; Grotjan, H.E.; Rapisarda, J.J.; Lederer, K.J.; Beltsos, A.N. Body mass index: Impact on IVF success appears age-related. Hum. Reprod. 2008, 23, 1835–1839. [Google Scholar] [CrossRef]
- Roubenoff, R.; Harris, T.B.; Abad, L.W.; Wilson, P.W.F.; Dallal, G.E.; Dinarello, C.A. Monocyte cytokine production in an elderly population: Effect of age and inflammation. J. Gerontol. Ser. A Boil. Sci. Med Sci. 1998, 53, M20–M26. [Google Scholar] [CrossRef] [PubMed]
- Kiecolt-Glaser, J.K.; Preacher, K.J.; Maccallum, R.C.; Atkinson, C.; Malarkey, W.B.; Glaser, R. Chronic stress and age-related increases in the proinflammatory cytokine IL-6. Proc. Natl. Acad. Sci. USA 2003, 100, 9090–9095. [Google Scholar] [CrossRef]
- Khaodhiar, L.; Ling, P.-R.; Blackburn, G.L.; Bistrian, B.R. Serum Levels of Interleukin-6 and C-Reactive Protein Correlate With Body Mass Index Across the Broad Range of Obesity. J. Parenter. Enter. Nutr. 2004, 28, 410–415. [Google Scholar] [CrossRef]
- Cioffi, J.A.; Van Blerkom, J.; Antczak, M.; Shafer, A.; Wittmer, S.; Snodgrass, H.R. The expression of leptin and its receptors in pre-ovulatory human follicles. Mol. Hum. Reprod. 1997, 3, 467–472. [Google Scholar] [CrossRef]
- Duggal, P.S.; Van der Hoek, K.H.; Milner, C.R.; Ryan, N.K.; Armstrong, D.T.; Magoffin, D.A.; Norman, R.J. The in vivo and in vitro effects of exogenous leptin on ovulation in the rat. Endocrinology 2000, 141, 1971–1976. [Google Scholar] [CrossRef]
- Swain, J.E.; Dunn, R.L.; McConnell, D.; Gonzalez-Martinez, J.; Smith, G.D. Direct Effects of Leptin on Mouse Reproductive Function: Regulation of Follicular, Oocyte, and Embryo Development. Biol. Reprod. 2004, 71, 1446–1452. [Google Scholar] [CrossRef]
- Sharabiani, M.T.A.; Vermeulen, R.; Scoccianti, C.; Hosnijeh, F.S.; Minelli, L.; Sacerdote, C.; Palli, D.; Krogh, V.; Tumino, R.; Chiodini, P.; et al. Immunologic profile of excessive body weight. Biomarkers 2011, 16, 243–251. [Google Scholar] [CrossRef]
- Zhang, Z.; Schlamp, F.; Huang, L.; Clark, H.; Brayboy, L.M. Inflammaging is associated with shifted macrophage ontogeny and polarization in the aging mouse ovary. Reproduction 2020, 159, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. 1), S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Zhang, X.; Zheng, S.; Khanabdali, R.; Kalionis, B.; Wu, J.; Wan, W.; Tai, X. An Update on Inflamm-Aging: Mechanisms, Prevention, and Treatment. J. Immunol. Res. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sanada, F.; Taniyama, Y.; Muratsu, J.; Otsu, R.; Shimizu, H.; Rakugi, H.; Morishita, R. Source of Chronic Inflammation in Aging. Front. Cardiovasc. Med. 2018, 5, 8426874. [Google Scholar] [CrossRef]
- van Deursen, J.M. The role of senescent cells in ageing. Nature 2014, 509, 439–446. [Google Scholar] [CrossRef]
- Chung, H.Y.; Ki, W.C.; Lee, E.K.; Chung, K.W.; Chung, S.; Lee, B.; Seo, A.Y.; Chung, J.H.; Jung, Y.S.; Im, E.; et al. Redefining Chronic Inflammation in Aging and Age-Related Diseases: Proposal of the Senoinflammation Concept. Aging Dis. 2019, 10, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.-P.; Desprez, P.-Y.; Krtolica, A.; Campisi, J. The Senescence-Associated Secretory Phenotype: The Dark Side of Tumor Suppression. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 99–118. [Google Scholar] [CrossRef]
- Schluns, K.S.; Kieper, W.C.; Jameson, S.C.; Lefrançois, L. Interleukin-7 mediates the homeostasis of naïve and memory CD8 T cells in vivo. Nat. Immunol. 2000, 1, 426–432. [Google Scholar] [CrossRef]
- Seddon, B.; Tomlinson, P.; Zamoyska, R. Interleukin 7 and T cell receptor signals regulate homeostasis of CD4 memory cells. Nat. Immunol. 2003, 4, 680–686. [Google Scholar] [CrossRef]
- Vujisi, S.; Lepej, S.I.; Emedi, I.; Bauman, R.; Remenar, A.; Tiljak, M.K.; Vujisić, S. Ovarian follicular concentration of IL-12, IL-15, IL-18 and p40 subunit of IL-12 and IL-23. Hum. Reprod. 2006, 21, 2650–2655. [Google Scholar] [CrossRef][Green Version]
- Morrisette-Thomas, V.; Cohen, A.A.; Fülöp, T.; Riesco, É.; Legault, V.; Li, Q.; Milot, E.; Dusseault-Bélanger, F.; Ferrucci, L. Inflamm-aging does not simply reflect increases in pro-inflammatory markers. Mech. Ageing Dev. 2014, 139, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Freund, A.; Orjalo, A.V.; Desprez, P.-Y.; Campisi, J. Inflammatory networks during cellular senescence: Causes and consequences. Trends Mol. Med. 2010, 16, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Faget, D.V.; Ren, Q.; Stewart, S.A. Unmasking senescence: Context-dependent effects of SASP in cancer. Nat. Rev. Cancer 2019, 19, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Ingman, W.V.; Robker, R.L.; Woittiez, K.; Robertson, S.A. Null mutation in transforming growth factor beta1 disrupts ovarian function and causes oocyte incompetence and early embryo arrest. Endocrinology 2006, 147, 835–845. [Google Scholar] [CrossRef]
- Halder, S.; Al-Hendy, A. Hypovitaminosis D and high serum transforming growth factor beta-3: Important biomarkers for uterine fibroids risk. Fertil. Steril. 2016, 106, 1648–1649. [Google Scholar] [CrossRef]
- Klar, R.M.; Duarte, R.; Dix-Peek, T.; Ripamonti, U. The induction of bone formation by the recombinant human transforming growth factor-β3. Biomaterials 2014, 35, 2773–2788. [Google Scholar] [CrossRef]
- Ohno, S.; Hirano, S.; Kanemaru, S.-I.; Tateya, I.; Kitani, Y.; Kojima, T.; Nakamura, T.; Ito, J. Prevention of buccal mucosa scarring with transforming growth factor β3. Laryngoscope 2011, 121, 1404–1409. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.E.; Doraiswamy, V.; Skinner, M.K. Transforming growth factor-beta isoform expression during bovine ovarian antral follicle development. Mol. Reprod. Dev. 2003, 66, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, P.A.; McCarty, J.H. TGF-β Activation and Signaling in Angiogenesis. In Physiologic and Pathologic Angiogenesis; Simionescu, D., Simionescu, A., Eds.; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Friedman, C.I.; Danforth, D.R.; Herbosa-Encarnacion, C.; Arbogast, L.; Alak, B.M.; Seifer, D.B. Follicular fluid vascular endothelial growth factor concentrations are elevated in women of advanced reproductive age undergoing ovulation induction. Fertil. Steril. 1997, 68, 607–612. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant ID | Age (Years) | AMH (ng/mL) | BMI (kg/m2) |
|---|---|---|---|
| Participant 01 | 27.7 | 1.8 | 26.3 |
| Participant 02 | 27.7 | 3.2 | 25.8 |
| Participant 03 | 28.8 | 2.1 | 19.9 |
| Participant 04 | 30.5 | 5.9 | 20.5 |
| Participant 05 | 30.5 | 0.9 | 20.2 |
| Participant 06 | 31.6 | 5.2 | 19.8 |
| Participant 07 | 31.9 | 2.7 | 24.1 |
| Participant 08 | 32.4 | 4.6 | 22.7 |
| Participant 09 | 32.6 | 2.0 | 24.0 |
| Participant 10 | 32.7 | 6.2 | 22.9 |
| Participant 11 | 32.9 | 1.2 | 21.2 |
| Participant 12 | 32.9 | 1.9 | 29.6 |
| Participant 13 | 33.6 | 3.6 | 21.1 |
| Participant 14 | 33.7 | 4.3 | 17.5 |
| Participant 15 | 34.0 | 3.9 | 25.7 |
| Participant 16 | 35.5 | 4.1 | 22.9 |
| Participant 17 | 35.6 | 0.2 | 23.0 |
| Participant 18 | 36.0 | 3.7 | 29.2 |
| Participant 19 | 36.8 | 1.4 | 23.6 |
| Participant 20 | 39.0 | 0.4 | 25.5 |
| Participant 21 | 39.0 | 4.3 | 28.3 |
| Participant 22 | 39.1 | 1.1 | 19.7 |
| Participant 23 | 39.9 | 5.3 | 21.1 |
| Participant 24 | 40.0 | 2.1 | 24.0 |
| Participant 25 | 40.2 | 1.0 | 28.4 |
| Participant 26 | 42.1 | 3.5 | 21.3 |
| Participant 27 | 42.4 | 0.6 | 21.3 |
| Participant 28 | 43.3 | 0.1 | 21.0 |
| Participant 29 | 44.2 | 0.3 | 26.1 |
| Participant 30 | 44.8 | 0.8 | 22.3 |
| Average (± SE) | 35.7 ± 0.90 | 2.6 ± 0.33 | 23.3 ± 0.57 |
| Parameters | Fibroinflammatory Cytokines |
|---|---|
| Age and AMH | IL-3, IL-7, IL-15, MIP-1β, TGF-β1, TGF-β3 |
| Age | M-CSF, SDF-1, EGF, oncostatin M, VEGF, PDGF-BB, BLC, CK β 8-1, BDNF, FGF-9, GDNF, IGFBP-3, LIF, MIF, PIGF, TGF-β2 |
| Non-significant | GRO, GRO-α, IL-β, IL-8, IL-12 p40, MCP-1, MCP-2, MCP-3, MIP-1δ, RANTES, SCF, IGF-1, angiogenin, leptin, eotaxin 1, eotaxin 2, eotaxin 3, FGF-4, FGF-6, FGF-7, Flt-3 ligand, fractalkine, GCP-2, HGF, IGFBP-1, IGFBP-2, IGFBP-4, IL-16, IP-10, LIGHT, MIP-3α, NAP-2, NT-3, NT-4, osteopontin, osteoprotegerin, PARC, TIMP-1, TIMP-2 |
| Group | Participant ID | Age | Cumulus Mass ID | Embryo Outcome |
|---|---|---|---|---|
| 1 | Participant 32 | 32.5 | a | Blastocyst day 6 |
| b | Blastocyst day 6 | |||
| Participant 33 | 44.7 | a | Blastocyst day 6 | |
| b | Blastocyst day 5 | |||
| 2 | Participant 34 | 33.9 | a | Hatching blastocyst day 5 |
| b | Hatching blastocyst day 5 | |||
| c | Hatching blastocyst day 6 | |||
| Participant 35 | 41.5 | a | Blastocyst day 5 | |
| b | Blastocyst day 6 | |||
| c | Blastocyst day 6 | |||
| 3 | Participant 36 | 34.4 | a | Blastocyst day 6 |
| b | Blastocyst day 6 | |||
| Participant 37 | 42.3 | a | 6 cell day 6 | |
| b | Mature (not fert.) | |||
| 4 | Participant 38 | 34.8 | a | Blastocyst day 5 |
| Participant 39 | 40.1 | a | Blastocyst day 5 | |
| 5 | Participant 40 | 34.9 | b | Early blastocyst day 6 |
| c | Morula day 6 | |||
| Participant 41 | 43 | a | 10 cell day 6 | |
| b | 6 cell day 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machlin, J.H.; Barishansky, S.J.; Kelsh, J.; Larmore, M.J.; Johnson, B.W.; Pritchard, M.T.; Pavone, M.E.; Duncan, F.E. Fibroinflammatory Signatures Increase with Age in the Human Ovary and Follicular Fluid. Int. J. Mol. Sci. 2021, 22, 4902. https://doi.org/10.3390/ijms22094902
Machlin JH, Barishansky SJ, Kelsh J, Larmore MJ, Johnson BW, Pritchard MT, Pavone ME, Duncan FE. Fibroinflammatory Signatures Increase with Age in the Human Ovary and Follicular Fluid. International Journal of Molecular Sciences. 2021; 22(9):4902. https://doi.org/10.3390/ijms22094902
Chicago/Turabian StyleMachlin, Jordan H., Seth J. Barishansky, John Kelsh, Megan J. Larmore, Brian W. Johnson, Michele T. Pritchard, Mary Ellen Pavone, and Francesca E. Duncan. 2021. "Fibroinflammatory Signatures Increase with Age in the Human Ovary and Follicular Fluid" International Journal of Molecular Sciences 22, no. 9: 4902. https://doi.org/10.3390/ijms22094902
APA StyleMachlin, J. H., Barishansky, S. J., Kelsh, J., Larmore, M. J., Johnson, B. W., Pritchard, M. T., Pavone, M. E., & Duncan, F. E. (2021). Fibroinflammatory Signatures Increase with Age in the Human Ovary and Follicular Fluid. International Journal of Molecular Sciences, 22(9), 4902. https://doi.org/10.3390/ijms22094902