
TRPM3 Is Expressed in Afferent Bladder Neurons and Is Upregulated during Bladder Inflammation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
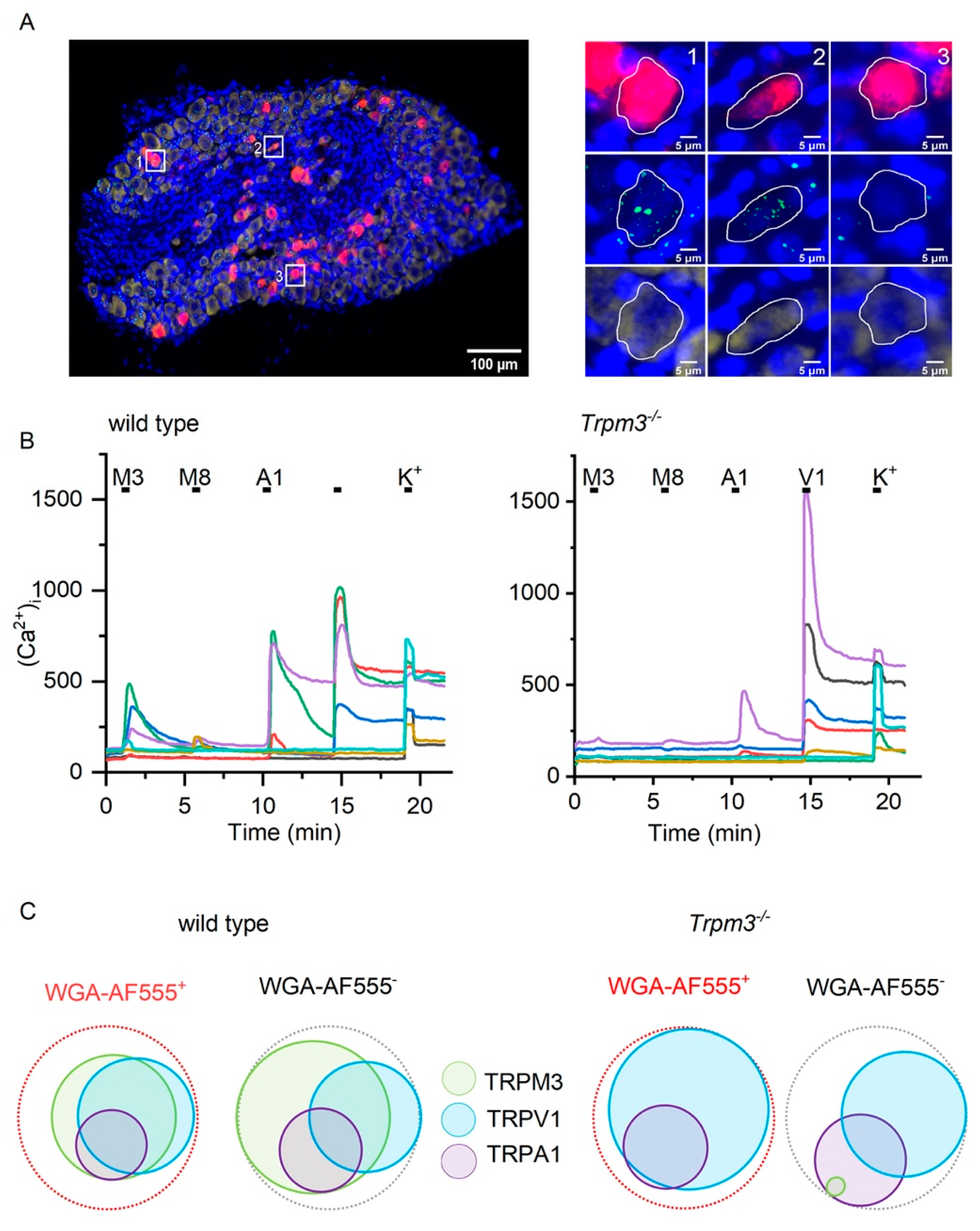
2.1. TRPM3 Is Expressed in Sensory Neurons Innervating the Bladder
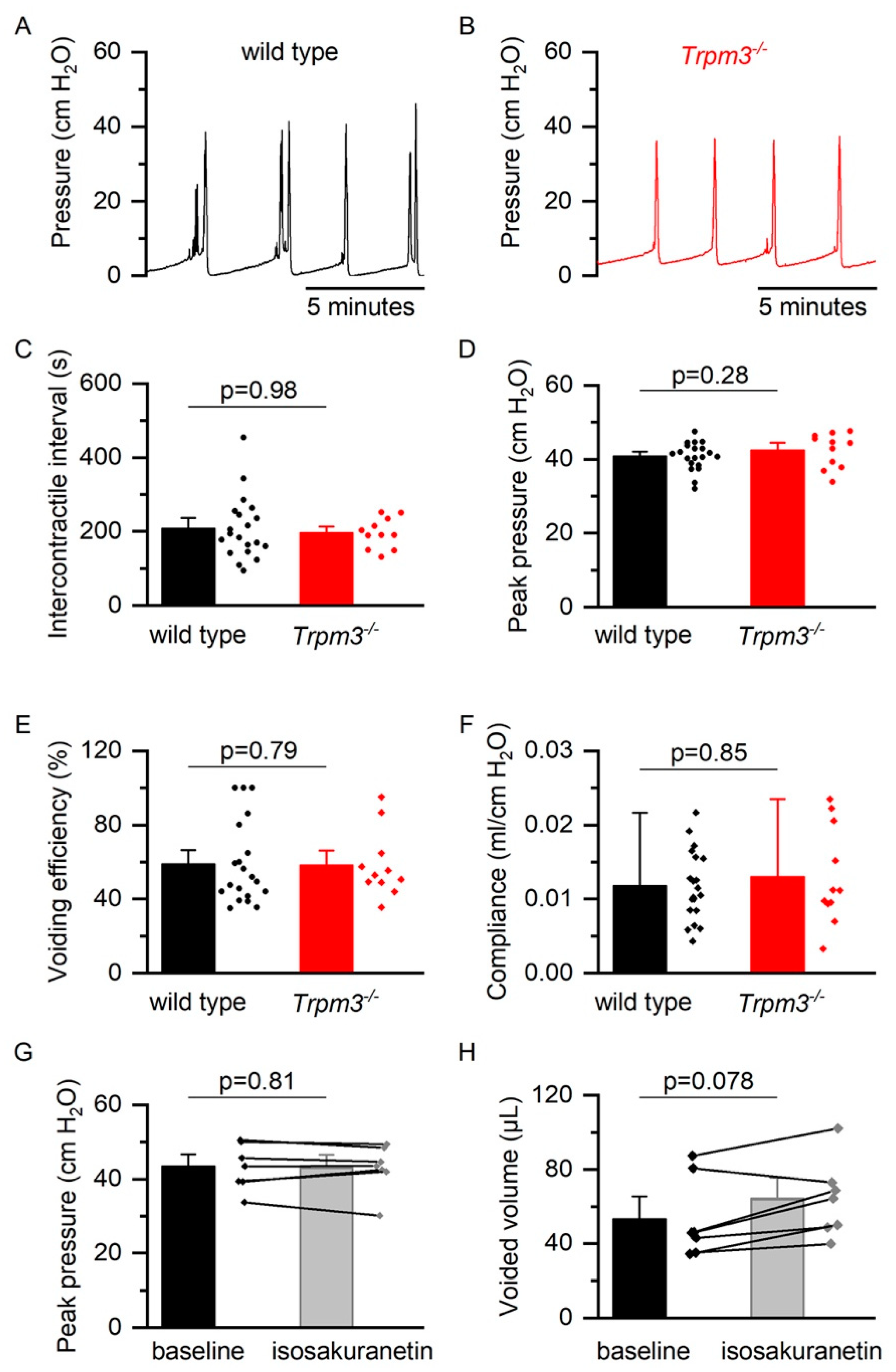
2.2. TRPM3 in Normal Bladder Function
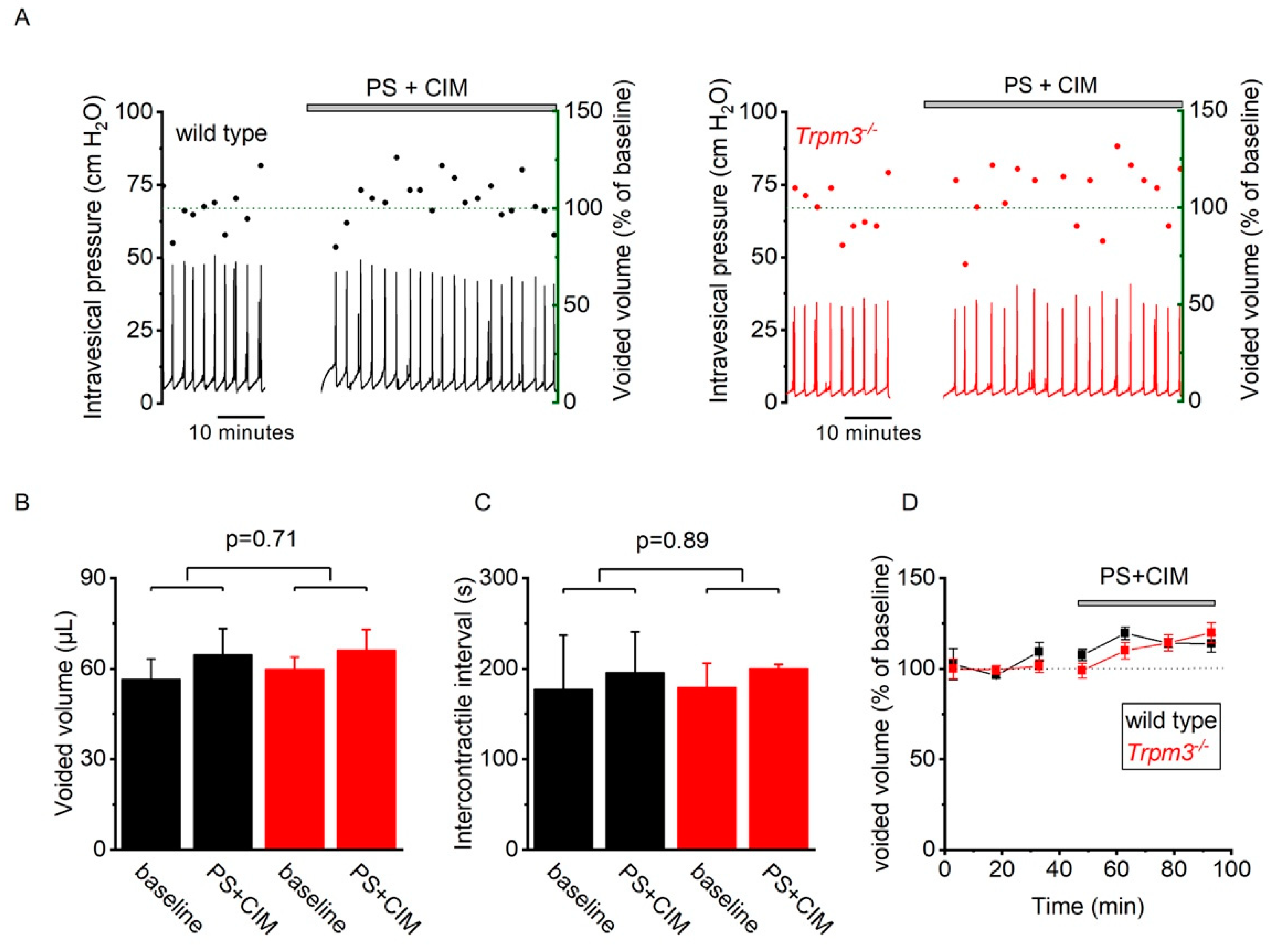
2.3. TRP Channel Function Is Upregulated during Bladder Inflammation
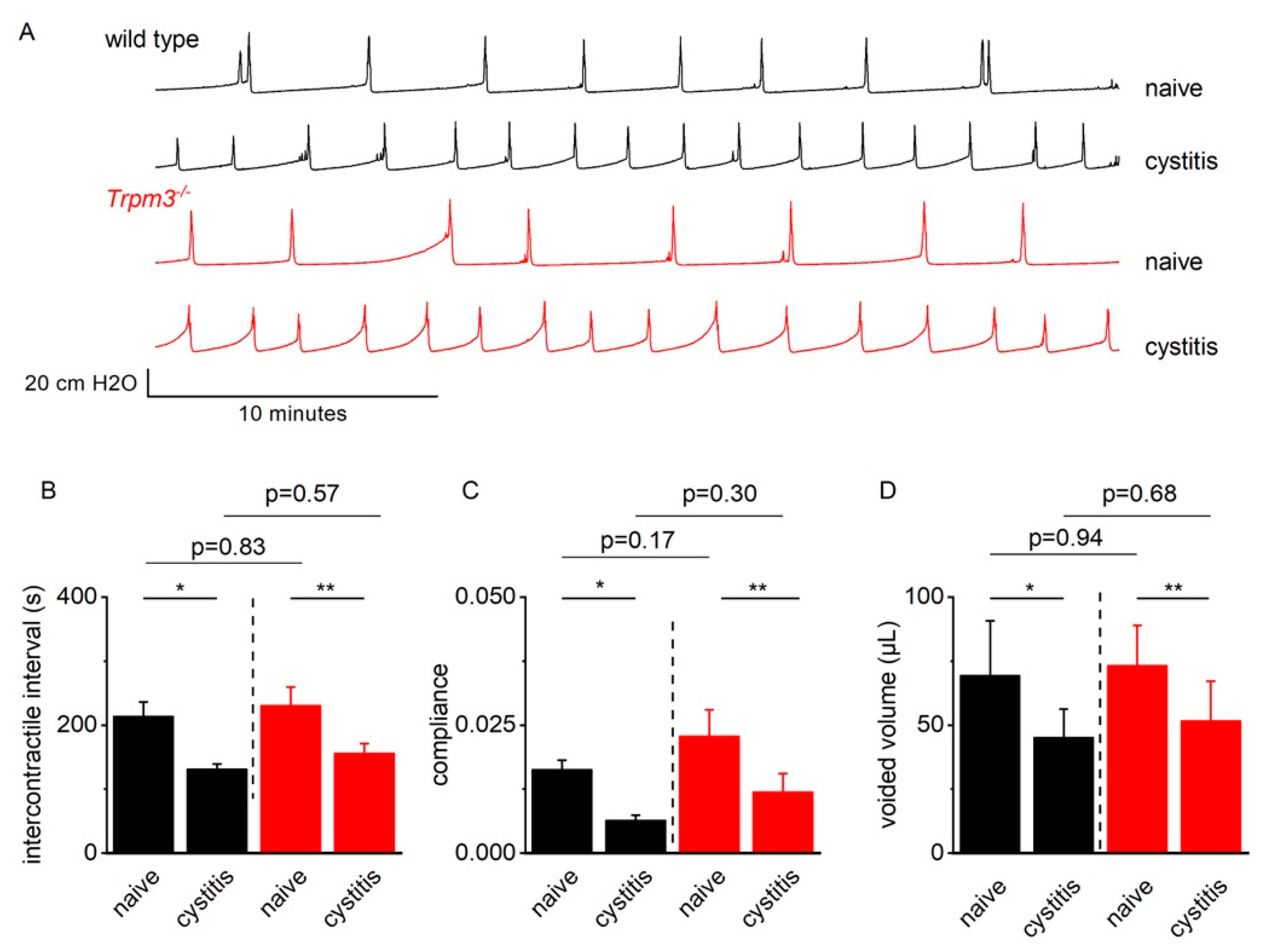
2.4. TRPM3 Does Not Contribute to Detrusor Hyperreflexia in Mice with CYP-Induced Cystitis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Retrograde Labeling of Bladder Neurons
4.3. Cyclophosphamide-Induced Cystitis
4.4. RNA Scope
4.5. Calcium Imaging of Bladder Afferent Neurons
4.6. Cystometry
4.7. Spotting Experiments
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Irwin, D.E.; Kopp, Z.S.; Agatep, B.; Milsom, I.; Abrams, P. Worldwide prevalence estimates of lower urinary tract symptoms, overactive bladder, urinary incontinence and bladder outlet obstruction. BJU Int. 2011, 108, 1132–1138. [Google Scholar] [CrossRef]
- Irwin, D.E.; Mungapen, L.; Milsom, I.; Kopp, Z.; Reeves, P.; Kelleher, C. The economic impact of overactive bladder syndrome in six Western countries. BJU Int. 2009, 103, 202–209. [Google Scholar] [CrossRef]
- Vanneste, M.; Segal, A.; Voets, T.; Everaerts, W. Transient receptor potential channels in sensory mechanisms of the lower urinary tract. Nat. Rev. Urol. 2021, 18, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Everaerts, W.; Zhen, X.; Ghosh, D.; Vriens, J.; Gevaert, T.; Gilbert, J.P.; Hayward, N.J.; McNamara, C.R.; Xue, F.; Moran, M.M.; et al. Inhibition of the cation channel TRPV4 improves bladder function in mice and rats with cyclophosphamide-induced cystitis. Proc. Natl. Acad. Sci. USA 2010, 107, 19084–19089. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-Y.; Wang, P.; Voznika Merriam, F.; Bjorling, D.E. Lack of TRPV1 inhibits cystitis-induced increased mechanical sensitivity in mice. Pain 2008, 139, 158–167. [Google Scholar] [CrossRef]
- Deberry, J.J.; Schwartz, E.S.; Davis, B.M. TRPA1 mediates bladder hyperalgesia in a mouse model of cystitis. Pain 2014, 155, 1280–1287. [Google Scholar] [CrossRef] [PubMed]
- Held, K.; Kichko, T.; De Clercq, K.; Klaassen, H.; Van Bree, R.; Vanherck, J.-C.; Marchand, A.; Reeh, P.W.; Chaltin, P.; Voets, T.; et al. Activation of TRPM3 by a potent synthetic ligand reveals a role in peptide release. Proc. Natl. Acad. Sci. USA 2015, 112, E1363–E1372. [Google Scholar] [CrossRef] [PubMed]
- Vandewauw, I.; De Clercq, K.; Mulier, M.; Held, K.; Pinto, S.; Van Ranst, N.; Segal, A.; Voet, T.; Vennekens, R.; Zimmermann, K.; et al. A TRP channel trio mediates acute noxious heat sensing. Nature 2018, 555, 662–666. [Google Scholar] [CrossRef]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S.; et al. TRPM3 Is a Nociceptor Channel Involved in the Detection of Noxious Heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef]
- Mulier, M.; Van Ranst, N.; Corthout, N.; Munck, S.; Vanden Berghe, P.; Vriens, J.; Voets, T.; Moilanen, L. Upregulation of TRPM3 in nociceptors innervating inflamed tissue. eLife 2020, 9, e61103. [Google Scholar] [CrossRef]
- Su, S.; Yudin, Y.; Kim, N.; Tao, Y.X.; Rohacs, T. TRPM3 Channels Play Roles in Heat Hypersensitivity and Spontaneous Pain after Nerve Injury. J. Neurosci. 2021, 41, 2457–2474. [Google Scholar] [CrossRef]
- Alkhatib, O.; Da Costa, R.; Gentry, C.; Quallo, T.; Bevan, S.; Andersson, D.A. Promiscuous G-Protein-Coupled Receptor Inhibition of Transient Receptor Potential Melastatin 3 Ion Channels by Gβγ Subunits. J. Neurosci. 2019, 39, 7840–7852. [Google Scholar] [CrossRef] [PubMed]
- Dembla, S.; Behrendt, M.; Mohr, F.; Goecke, C.; Sondermann, J.; Schneider, F.M.; Schmidt, M.; Stab, J.; Enzeroth, R.; Leitner, M.G.; et al. Anti-nociceptive action of peripheral mu-opioid receptors by G-beta-gamma protein-mediated inhibition of TRPM3 channels. eLife 2017, 6, e26280. [Google Scholar] [CrossRef]
- Straub, I.; Krügel, U.; Mohr, F.; Teichert, J.; Rizun, O.; Konrad, M.; Oberwinkler, J.; Schaefer, M. Flavanones That Selectively Inhibit TRPM3 Attenuate Thermal Nociception In Vivo. Mol. Pharmacol. 2013, 84, 736–750. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Zhang, Y.; Yu, J. Antinociceptive Effects of Isosakuranetin in a Rat Model of Peripheral Neuropathy. Pharmacology 2017, 100, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Grundy, L.; Caldwell, A.; Garcia Caraballo, S.; Erickson, A.; Schober, G.; Castro, J.; Harrington, A.M.; Brierley, S.M.; Caraballo, G.S. Histamine induces peripheral and central hypersensitivity to bladder distension via the histamine H 1 receptor and TRPV1. Am. J. Physiol. Ren. Physiol. 2020, 318, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Araki, I.; Yoshiyama, M.; Nomura, T.; Takeda, M. Transient Receptor Potential Channel A1 Involved in Sensory Transduction of Rat Urinary Bladder Through C-Fiber Pathway. Urology 2007, 70, 826–831. [Google Scholar] [CrossRef]
- O’Mullane, L.M.; Keast, J.R.; Osborne, P.B. Co-Cultures Provide a New Tool to Probe Communication Between Adult Sensory Neurons and Urothelium. J. Urol. 2013, 190, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Charrua, A.; Cruz, C.D.; Cruz, F.; Avelino, A. Transient Receptor Potential Vanilloid Subfamily 1 is Essential for the Generation of Noxious Bladder Input and Bladder Overactivity in Cystitis. J. Urol. 2007, 177, 1537–1541. [Google Scholar] [CrossRef]
- Kamei, J.; Aizawa, N.; Nakagawa, T.; Kaneko, S.; Kume, H.; Homma, Y.; Igawa, Y. Attenuated lipopolysaccharide-induced inflammatory bladder hypersensitivity in mice deficient of transient receptor potential ankilin1. Sci. Rep. 2018, 8, 15622. [Google Scholar] [CrossRef] [PubMed]
- Uvin, P.; Franken, J.; Pinto, S.; Rietjens, R.; Grammet, L.; Deruyver, Y.; Alpizar, Y.A.; Talavera, K.; Vennekens, R.; Everaerts, W.; et al. Essential Role of Transient Receptor Potential M8 (TRPM8) in a Model of Acute Cold-induced Urinary Urgency. Eur. Urol. 2015, 68, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Izgi, K.; Altuntas, C.Z.; Bicer, F.; Ozer, A.; Sakalar, C.; Li, X.; Tuohy, V.K.; Daneshgari, F. Uroplakin Peptide-Specific Autoimmunity Initiates Interstitial Cystitis/Painful Bladder Syndrome in Mice. PLoS ONE 2013, 8, e72067. [Google Scholar] [CrossRef]
- Clodfelder-Miller, B.J.; Kanda, H.; Gu, J.G.; Creighton, J.R.; Ness, T.J.; DeBerry, J.J. Urothelial bladder afferent neurons in the rat are anatomically and neurochemically distinct from non-urothelial afferents. Brain Res. 2017, 1689, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Clodfelder-Miller, B.J.; Gu, J.G.; Ness, T.J.; DeBerry, J.J. Electrophysiological properties of lumbosacral primary afferent neurons innervating urothelial and non-urothelial layers of mouse urinary bladder. Brain Res. 2016, 1648, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.J. Cyclophosphamide cystitis—Identification of acrolein as the causative agent. Biochem. Pharmacol. 1979, 28, 2045–2049. [Google Scholar] [CrossRef]
- Uvin, P.; Everaerts, W.; Pinto, S.; Alpízar, Y.A.; Boudes, M.; Gevaert, T.; Voets, T.; Nilius, B.; Talavera, K.; De Ridder, D. The Use of Cystometry in Small Rodents: A Study of Bladder Chemosensation. J. Vis. Exp. 2012, 66, e3869. [Google Scholar] [CrossRef]
- Andersson, K.E.; Soler, R.; Füllhase, C. Rodent models for urodynamic investigation. Neurourol. Urodyn. 2011, 30, 636–646. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanneste, M.; Mulier, M.; Nogueira Freitas, A.C.; Van Ranst, N.; Kerstens, A.; Voets, T.; Everaerts, W. TRPM3 Is Expressed in Afferent Bladder Neurons and Is Upregulated during Bladder Inflammation. Int. J. Mol. Sci. 2022, 23, 107. https://doi.org/10.3390/ijms23010107
Vanneste M, Mulier M, Nogueira Freitas AC, Van Ranst N, Kerstens A, Voets T, Everaerts W. TRPM3 Is Expressed in Afferent Bladder Neurons and Is Upregulated during Bladder Inflammation. International Journal of Molecular Sciences. 2022; 23(1):107. https://doi.org/10.3390/ijms23010107
Chicago/Turabian StyleVanneste, Matthias, Marie Mulier, Ana Cristina Nogueira Freitas, Nele Van Ranst, Axelle Kerstens, Thomas Voets, and Wouter Everaerts. 2022. "TRPM3 Is Expressed in Afferent Bladder Neurons and Is Upregulated during Bladder Inflammation" International Journal of Molecular Sciences 23, no. 1: 107. https://doi.org/10.3390/ijms23010107
APA StyleVanneste, M., Mulier, M., Nogueira Freitas, A. C., Van Ranst, N., Kerstens, A., Voets, T., & Everaerts, W. (2022). TRPM3 Is Expressed in Afferent Bladder Neurons and Is Upregulated during Bladder Inflammation. International Journal of Molecular Sciences, 23(1), 107. https://doi.org/10.3390/ijms23010107