
Infections, Reactions of Natural Killer T Cells and Natural Killer Cells, and Kidney Injury

Abstract
:1. Introduction
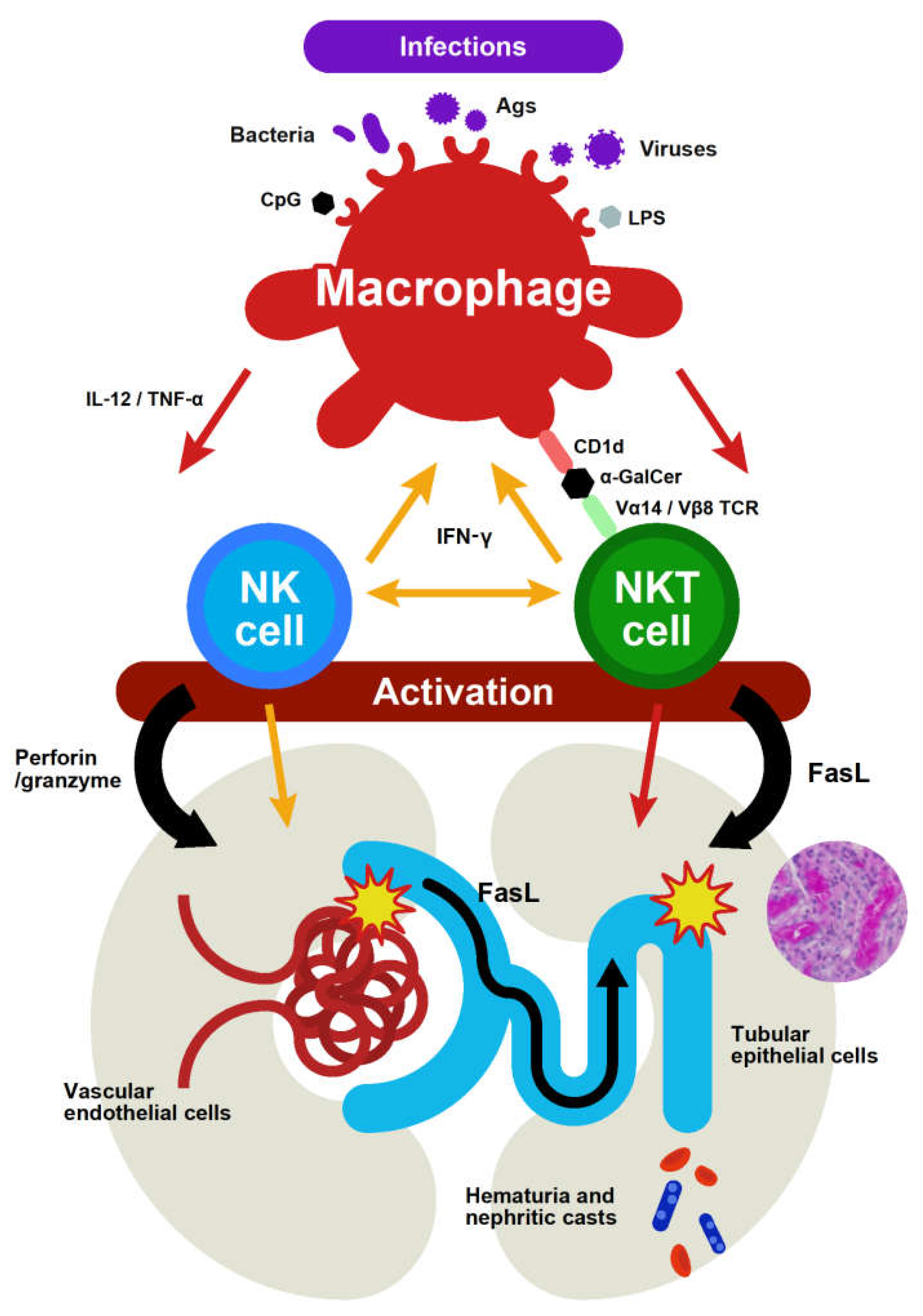
2. Roles of Mouse NKT Cells and NK Cells in Infection-Associated Kidney Injury
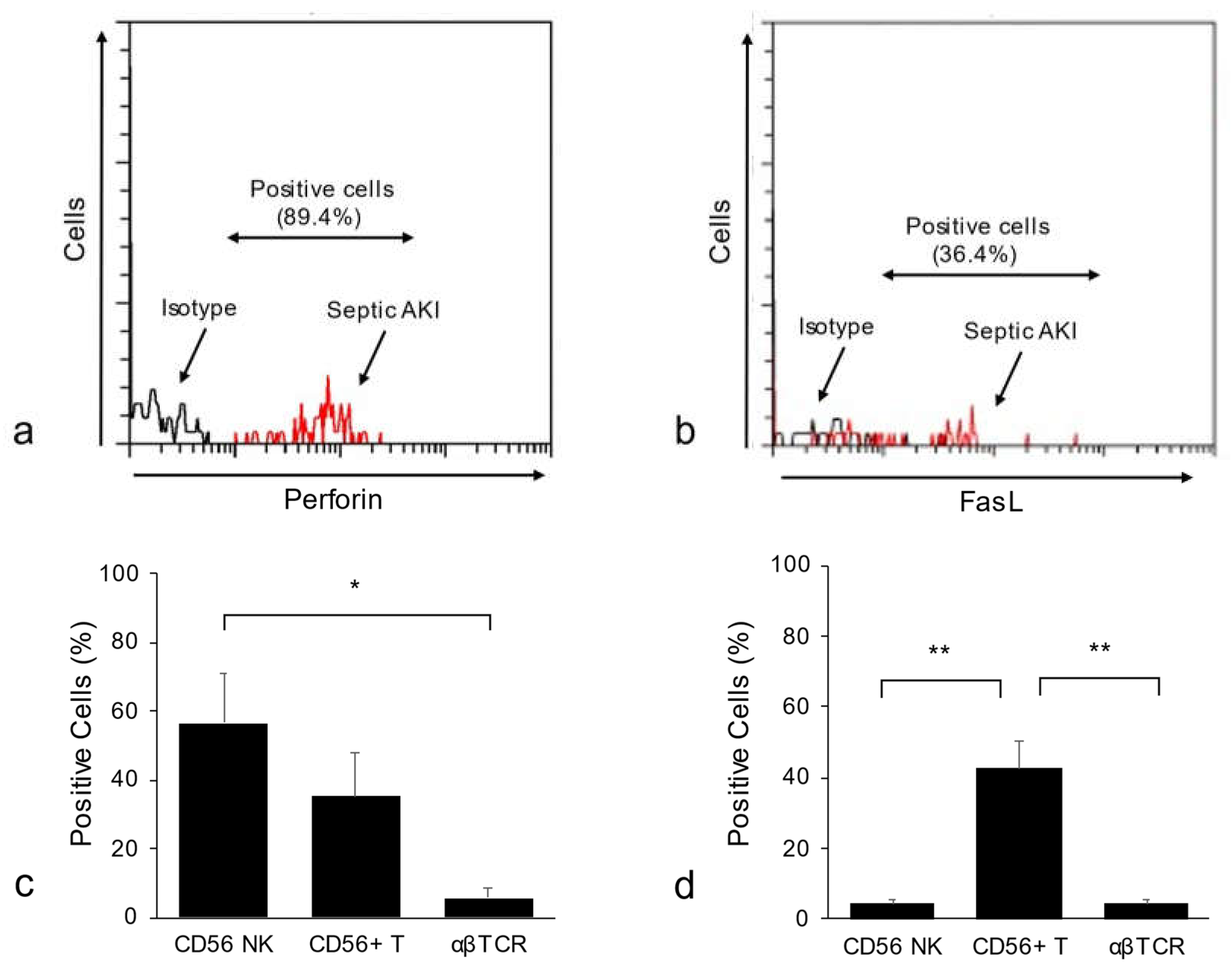
3. Associations among Infections, Human CD56+ T Cells and CD56+ NK Cells, and Renal Diseases
4. Two Distinct Macrophage Subsets in the Liver and Kidney and Their Interactions with NKT Cells
5. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prasad, N.; Patel, M.R. Infection-Induced Kidney Diseases. Front. Med. 2018, 5, 327. [Google Scholar] [CrossRef] [PubMed]
- Ludes, P.O.; de Roquetaillade, C.; Chousterman, B.G.; Pottecher, J.; Mebazaa, A. Role of Damage-Associated Molecular Patterns in Septic Acute Kidney Injury, from Injury to Recovery. Front. Immunol. 2021, 12, 606622. [Google Scholar] [CrossRef]
- Peters, E.; Masereeuw, R.; Pickkers, P. The potential of alkaline phosphatase as a treatment for sepsis-associated acute kidney injury. Nephron Clin. Pract. 2014, 127, 144–148. [Google Scholar] [CrossRef]
- Oda, T.; Yoshizawa, N. Factors Affecting the Progression of Infection-Related Glomerulonephritis to Chronic Kidney Disease. Int. J. Mol. Sci. 2021, 22, 905. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zeng, M.; Li, M.; Kan, Y.; Li, B.; Xu, R.; Wu, Y.; Wang, S.; Zheng, X.; Feng, W. Protopine Protects Mice against LPS-Induced Acute Kidney Injury by Inhibiting Apoptosis and Inflammation via the TLR4 Signaling Pathway. Molecules 2019, 25, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, H.J.; Wilkens, L.; Schraml, B.; Marschner, J. One concept does not fit all: The immune system in different forms of acute kidney injury. Nephrol. Dial. Transplant. 2021, 36, 29–38. [Google Scholar] [CrossRef]
- Uchida, T.; Nakashima, H.; Ito, S.; Ishikiriyama, T.; Nakashima, M.; Seki, S.; Kumagai, H.; Oshima, N. Activated natural killer T cells in mice induce acute kidney injury with hematuria through possibly common mechanisms shared by human CD56(+) T cells. Am. J. Physiol. Ren. Physiol. 2018, 315, F618–F627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, T.; Ito, S.; Kumagai, H.; Oda, T.; Nakashima, H.; Seki, S. Roles of Natural Killer T Cells and Natural Killer Cells in Kidney Injury. Int. J. Mol. Sci. 2019, 20, 2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.E.; Becker, M.; Mittrucker, H.W.; Panzer, U. Tissue-Resident Lymphocytes in the Kidney. J. Am. Soc. Nephrol. 2018, 29, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Seki, S.; Habu, Y.; Kawamura, T.; Takeda, K.; Dobashi, H.; Ohkawa, T.; Hiraide, H. The liver as a crucial organ in the first line of host defense: The roles of Kupffer cells, natural killer (NK) cells and NK1.1 Ag+ T cells in T helper 1 immune responses. Immunol. Rev. 2000, 174, 35–46. [Google Scholar] [CrossRef]
- Flahou, C.; Morishima, T.; Takizawa, H.; Sugimoto, N. Fit-For-All iPSC-Derived Cell Therapies and Their Evaluation in Humanized Mice with NK Cell Immunity. Front. Immunol. 2021, 12, 662360. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.E.; Rickassel, C.; Healy, H.; Kassianos, A.J. Natural Killer Cells in Kidney Health and Disease. Front. Immunol. 2019, 10, 587. [Google Scholar] [CrossRef] [Green Version]
- Habu, Y.; Uchida, T.; Inui, T.; Nakashima, H.; Fukasawa, M.; Seki, S. Enhancement of the synthetic ligand-mediated function of liver NK1.1Ag+ T cells in mice by interleukin-12 pretreatment. Immunology 2004, 113, 35–43. [Google Scholar] [CrossRef]
- Kaminski, H.; Couzi, L.; Eberl, M. Unconventional T cells and kidney disease. Nat. Rev. Nephrol. 2021, 17, 795–813. [Google Scholar] [CrossRef]
- Inui, T.; Nakashima, H.; Habu, Y.; Nakagawa, R.; Fukasawa, M.; Kinoshita, M.; Shinomiya, N.; Seki, S. Neutralization of tumor necrosis factor abrogates hepatic failure induced by alpha-galactosylceramide without attenuating its antitumor effect in aged mice. J. Hepatol. 2005, 43, 670–678. [Google Scholar] [CrossRef]
- Seki, S.; Nakashima, H.; Nakashima, M.; Kinoshita, M. Antitumor immunity produced by the liver Kupffer cells, NK cells, NKT cells, and CD8 CD122 T cells. Clin. Dev. Immunol. 2011, 2011, 868345. [Google Scholar] [CrossRef] [Green Version]
- Takayama, E.; Koike, Y.; Ohkawa, T.; Majima, T.; Fukasawa, M.; Shinomiya, N.; Yamaguchi, T.; Konishi, M.; Hiraide, H.; Tadakuma, T.; et al. Functional and Vbeta repertoire characterization of human CD8+ T-cell subsets with natural killer cell markers, CD56+ CD57- T cells, CD56+ CD57+ T cells and CD56- CD57+ T cells. Immunology 2003, 108, 211–219. [Google Scholar] [CrossRef]
- Kronenberg, M. Toward an understanding of NKT cell biology: Progress and paradoxes. Annu. Rev. Immunol. 2005, 23, 877–900. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, Y.; Sonoda, T.; Shida, H.; Kusunoki, Y.; Hattanda, F.; Tanimura, S.; Uozumi, R.; Yamada, M.; Nishibata, Y.; Masuda, S.; et al. Detection of Autoreactive Type II NKT Cells: A Pilot Study of Comparison between Healthy Individuals and Patients with Vasculitis. Cytom. Part A 2018, 93, 1157–1164. [Google Scholar] [CrossRef] [Green Version]
- Van Acker, H.H.; Capsomidis, A.; Smits, E.L.; Van Tendeloo, V.F. CD56 in the Immune System: More Than a Marker for Cytotoxicity? Front. Immunol. 2017, 8, 892. [Google Scholar] [CrossRef]
- Uchida, T.; Nakashima, H.; Yamagata, A.; Ito, S.; Ishikiriyama, T.; Nakashima, M.; Seki, S.; Kumagai, H.; Oshima, N. Repeated administration of alpha-galactosylceramide ameliorates experimental lupus nephritis in mice. Sci. Rep. 2018, 8, 8225. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, T.; Seki, S.; Dobashi, H.; Koike, Y.; Habu, Y.; Ami, K.; Hiraide, H.; Sekine, I. Systematic characterization of human CD8+ T cells with natural killer cell markers in comparison with natural killer cells and normal CD8+ T cells. Immunology 2001, 103, 281–290. [Google Scholar] [CrossRef]
- Kawabata, T.; Kinoshita, M.; Inatsu, A.; Habu, Y.; Nakashima, H.; Shinomiya, N.; Seki, S. Functional alterations of liver innate immunity of mice with aging in response to CpG-oligodeoxynucleotide. Hepatology 2008, 48, 1586–1597. [Google Scholar] [CrossRef]
- Ferhat, M.; Robin, A.; Giraud, S.; Sena, S.; Goujon, J.M.; Touchard, G.; Hauet, T.; Girard, J.P.; Gombert, J.M.; Herbelin, A.; et al. Endogenous IL-33 Contributes to Kidney Ischemia-Reperfusion Injury as an Alarmin. J. Am. Soc. Nephrol. 2018, 29, 1272–1288. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Han, C.; Dai, H.; Hou, J.; Dong, Y.; Cui, X.; Xu, L.; Zhang, M.; Xia, Q. Hypoxia-Inducible Factor-2alpha Limits Natural Killer T Cell Cytotoxicity in Renal Ischemia/Reperfusion Injury. J. Am. Soc. Nephrol. 2016, 27, 92–106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.X.; Wang, S.; Huang, X.; Min, W.P.; Sun, H.; Liu, W.; Garcia, B.; Jevnikar, A.M. NK cells induce apoptosis in tubular epithelial cells and contribute to renal ischemia-reperfusion injury. J. Immunol. 2008, 181, 7489–7498. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.H.; Lee, J.P.; Jang, H.R.; Cha, R.H.; Han, S.S.; Jeon, U.S.; Kim, D.K.; Song, J.; Lee, D.S.; Kim, Y.S. Sulfatide-reactive natural killer T cells abrogate ischemia-reperfusion injury. J. Am. Soc. Nephrol. 2011, 22, 1305–1314. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, H.; Inui, T.; Habu, Y.; Kinoshita, M.; Nagao, S.; Kawaguchi, A.; Miura, S.; Shinomiya, N.; Yagita, H.; Seki, S. Activation of mouse natural killer T cells accelerates liver regeneration after partial hepatectomy. Gastroenterology 2006, 131, 1573–1583. [Google Scholar] [CrossRef]
- Takahashi, E.; Kuranaga, N.; Satoh, K.; Habu, Y.; Shinomiya, N.; Asano, T.; Seki, S.; Hayakawa, M. Induction of CD16+ CD56bright NK cells with antitumour cytotoxicity not only from CD16- CD56bright NK Cells but also from CD16- CD56dim NK cells. Scand. J. Immunol. 2007, 65, 126–138. [Google Scholar] [CrossRef]
- Hirano, K.; Matsuzaki, K.; Yasuda, T.; Nishikawa, M.; Yasuda, Y.; Koike, K.; Maruyama, S.; Yokoo, T.; Matsuo, S.; Kawamura, T.; et al. Association Between Tonsillectomy and Outcomes in Patients with Immunoglobulin A Nephropathy. JAMA Netw. Open 2019, 2, e194772. [Google Scholar] [CrossRef] [Green Version]
- Hotta, O.; Taguma, Y.; Yusa, N.; Ooyama, M. Analysis of mononuclear cells in urine using flow cytometry in glomerular diseases. Kidney Int. Suppl. 1994, 47, S117–S121. [Google Scholar] [PubMed]
- Hotta, O.; Taguma, Y.; Ooyama, M.; Yusa, N.; Nagura, H. Analysis of CD14+ cells and CD56+ cells in urine using flow cytometry: A useful tool for monitoring disease activity of IgA nephropathy. Clin. Nephrol. 1993, 39, 289–294. [Google Scholar] [PubMed]
- Cox, S.N.; Sallustio, F.; Serino, G.; Loverre, A.; Pesce, F.; Gigante, M.; Zaza, G.; Stifanelli, P.F.; Ancona, N.; Schena, F.P. Activated innate immunity and the involvement of CX3CR1-fractalkine in promoting hematuria in patients with IgA nephropathy. Kidney Int. 2012, 82, 548–560. [Google Scholar] [CrossRef] [Green Version]
- Iwatani, H.; Nagasawa, Y.; Yamamoto, R.; Iio, K.; Mizui, M.; Horii, A.; Kitahara, T.; Inohara, H.; Kumanogoh, A.; Imai, E.; et al. CD16+CD56+ cells are a potential culprit for hematuria in IgA nephropathy. Clin. Exp. Nephrol. 2015, 19, 216–224. [Google Scholar] [CrossRef]
- Hotta, O.; Oda, T. The epipharynx-kidney axis triggers glomerular vasculitis in immunoglobulin A nephropathy. Immunol. Res. 2019, 67, 304–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kano, T.; Suzuki, H.; Makita, Y.; Fukao, Y.; Suzuki, Y. Nasal-associated lymphoid tissue is the major induction site for nephritogenic IgA in murine IgA nephropathy. Kidney Int. 2021, 100, 364–376. [Google Scholar] [CrossRef]
- Nasr, S.H.; Fidler, M.E.; Valeri, A.M.; Cornell, L.D.; Sethi, S.; Zoller, A.; Stokes, M.B.; Markowitz, G.S.; D’Agati, V.D. Postinfectious glomerulonephritis in the elderly. J. Am. Soc. Nephrol. 2011, 22, 187–195. [Google Scholar] [CrossRef]
- Nasr, S.H.; Radhakrishnan, J.; D’Agati, V.D. Bacterial infection-related glomerulonephritis in adults. Kidney Int. 2013, 83, 792–803. [Google Scholar] [CrossRef] [Green Version]
- Oda, T.; Yoshizawa, N.; Yamakami, K.; Sakurai, Y.; Takechi, H.; Yamamoto, K.; Oshima, N.; Kumagai, H. The role of nephritis-associated plasmin receptor (NAPlr) in glomerulonephritis associated with streptococcal infection. J. Biomed. Biotechnol. 2012, 2012, 417675. [Google Scholar] [CrossRef] [PubMed]
- Uchida, T.; Oda, T. Glomerular Deposition of Nephritis-Associated Plasmin Receptor (NAPlr) and Related Plasmin Activity: Key Diagnostic Biomarkers of Bacterial Infection-related Glomerulonephritis. Int. J. Mol. Sci. 2020, 21, 2595. [Google Scholar] [CrossRef] [Green Version]
- Halloran, P.F.; Famulski, K.S.; Reeve, J. Molecular assessment of disease states in kidney transplant biopsy samples. Nat. Rev. Nephrol. 2016, 12, 534–548. [Google Scholar] [CrossRef] [PubMed]
- Calvani, J.; Terada, M.; Lesaffre, C.; Eloudzeri, M.; Lamarthee, B.; Burger, C.; Tinel, C.; Anglicheau, D.; Vermorel, A.; Couzi, L.; et al. In situ multiplex immunofluorescence analysis of the inflammatory burden in kidney allograft rejection: A new tool to characterize the alloimmune response. Am. J. Transplant. 2020, 20, 942–953. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Ito, S.; Ishikiriyama, T.; Sekiguchi, K.; Yamaguchi, R.; Tsuruhara, R.; Matsuda, A.; Koiwa, K.; Nakashima, M.; Nakashima, H.; et al. The Efficacy of Posttreatment with Synthetic C-Reactive Protein in Murine Bacterial Peritonitis via Activation of FcgammaRI-Expressing Kupffer Cells. J. Innate Immun. 2021, 13, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Inatsu, A.; Kinoshita, M.; Nakashima, H.; Shimizu, J.; Saitoh, D.; Tamai, S.; Seki, S. Novel mechanism of C-reactive protein for enhancing mouse liver innate immunity. Hepatology 2009, 49, 2044–2054. [Google Scholar] [CrossRef]
- Sato, A.; Nakashima, H.; Kinoshita, M.; Nakashima, M.; Ogawa, Y.; Shono, S.; Ikarashi, M.; Seki, S. The effect of synthetic C-reactive protein on the in vitro immune response of human PBMCs stimulated with bacterial reagents. Inflammation 2013, 36, 781–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickkers, P.; Mehta, R.L.; Murray, P.T.; Joannidis, M.; Molitoris, B.A.; Kellum, J.A.; Bachler, M.; Hoste, E.A.J.; Hoiting, O.; Krell, K.; et al. Effect of Human Recombinant Alkaline Phosphatase on 7-Day Creatinine Clearance in Patients With Sepsis-Associated Acute Kidney Injury: A Randomized Clinical Trial. JAMA 2018, 320, 1998–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.; Huang, J.; Huang, X.; Han, X.; Tang, W.; Ke, G.; Xu, Q. Effect of alkaline phosphatase on sepsis-associated acute kidney injury patients: A systematic review and meta-analysis. Medicine 2020, 99, e18788. [Google Scholar] [CrossRef] [PubMed]
- Soni, H.; Adebiyi, A. Early septic insult in neonatal pigs increases serum and urinary soluble Fas ligand and decreases kidney function without inducing significant renal apoptosis. Ren. Fail. 2017, 39, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Miao, H.; Shi, J.; Wang, C.; Lu, G.; Zhu, X.; Wang, Y.; Cui, Y.; Zhang, Y. Continuous Renal Replacement Therapy in Pediatric Severe Sepsis: A Propensity Score-Matched Prospective Multicenter Cohort Study in the PICU. Crit. Care Med. 2019, 47, e806–e813. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Uchida, T.; Sato, A.; Nakashima, M.; Nakashima, H.; Shono, S.; Habu, Y.; Miyazaki, H.; Hiroi, S.; Seki, S. Characterization of two F4/80-positive Kupffer cell subsets by their function and phenotype in mice. J. Hepatol. 2010, 53, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, M.; Nakashima, H.; Kinoshita, M.; Sato, A.; Nakashima, M.; Miyazaki, H.; Nishiyama, K.; Yamamoto, J.; Seki, S. Distinct development and functions of resident and recruited liver Kupffer cells/macrophages. J. Leukoc. Biol. 2013, 94, 1325–1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikiriyama, T.; Nakashima, H.; Endo-Umeda, K.; Nakashima, M.; Ito, S.; Kinoshita, M.; Ikarashi, M.; Makishima, M.; Seki, S. Contrasting functional responses of resident Kupffer cells and recruited liver macrophages to irradiation and liver X receptor stimulation. PLoS ONE 2021, 16, e0254886. [Google Scholar] [CrossRef]
- Ito, S.; Nakashima, H.; Ishikiriyama, T.; Nakashima, M.; Yamagata, A.; Imakiire, T.; Kinoshita, M.; Seki, S.; Kumagai, H.; Oshima, N. Effects of a CCR2 antagonist on macrophages and Toll-like receptor 9 expression in a mouse model of diabetic nephropathy. Am. J. Physiol. Ren. Physiol. 2021, 321, F757–F770. [Google Scholar] [CrossRef]
- Nakagawa, R.; Nagafune, I.; Tazunoki, Y.; Ehara, H.; Tomura, H.; Iijima, R.; Motoki, K.; Kamishohara, M.; Seki, S. Mechanisms of the antimetastatic effect in the liver and of the hepatocyte injury induced by alpha-galactosylceramide in mice. J. Immunol. 2001, 166, 6578–6584. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, K.; Nakashima, H.; Ikarashi, M.; Kinoshita, M.; Nakashima, M.; Aosasa, S.; Seki, S.; Yamamoto, J. Mouse CD11b+Kupffer Cells Recruited from Bone Marrow Accelerate Liver Regeneration after Partial Hepatectomy. PLoS ONE 2015, 10, e0136774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, H.; Kinoshita, M.; Nakashima, M.; Habu, Y.; Shono, S.; Uchida, T.; Shinomiya, N.; Seki, S. Superoxide produced by Kupffer cells is an essential effector in concanavalin A-induced hepatitis in mice. Hepatology 2008, 48, 1979–1988. [Google Scholar] [CrossRef] [PubMed]
- Korf, H.; Vander Beken, S.; Romano, M.; Steffensen, K.R.; Stijlemans, B.; Gustafsson, J.A.; Grooten, J.; Huygen, K. Liver X receptors contribute to the protective immune response against Mycobacterium tuberculosis in mice. J. Clin. Invest. 2009, 119, 1626–1637. [Google Scholar] [CrossRef] [Green Version]
- Endo-Umeda, K.; Nakashima, H.; Uno, S.; Toyoshima, S.; Umeda, N.; Komine-Aizawa, S.; Seki, S.; Makishima, M. Liver X receptors regulate natural killer T cell population and antitumor activity in the liver of mice. Sci. Rep. 2021, 11, 22595. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Causative Pathogens | Roles of NKT and/or NK Cells | Type of Kidney Injury |
|---|---|---|
| CpG-ODN *1 [23] | Tubular epithelial cell injury via the TNF-α/FasL system | AKI in mice |
| LPS (the generalized Shwartzman reaction) [8] | Renal vascular endothelial cell injury via the perforin-mediated pathway | |
| Certain bacteria | Increase in expression of perforin and FasL | Sepsis-associated AKI in human |
| CpG-ODN *2 [36] | Mechanism remains to be solved | Exacerbation of murine IgAN |
| Certain bacteria | Injury of glomerular endothelial cells leading to rupture of capillary walls | Exacerbation of human IgAN |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uchida, T.; Seki, S.; Oda, T. Infections, Reactions of Natural Killer T Cells and Natural Killer Cells, and Kidney Injury. Int. J. Mol. Sci. 2022, 23, 479. https://doi.org/10.3390/ijms23010479
Uchida T, Seki S, Oda T. Infections, Reactions of Natural Killer T Cells and Natural Killer Cells, and Kidney Injury. International Journal of Molecular Sciences. 2022; 23(1):479. https://doi.org/10.3390/ijms23010479
Chicago/Turabian StyleUchida, Takahiro, Shuhji Seki, and Takashi Oda. 2022. "Infections, Reactions of Natural Killer T Cells and Natural Killer Cells, and Kidney Injury" International Journal of Molecular Sciences 23, no. 1: 479. https://doi.org/10.3390/ijms23010479
APA StyleUchida, T., Seki, S., & Oda, T. (2022). Infections, Reactions of Natural Killer T Cells and Natural Killer Cells, and Kidney Injury. International Journal of Molecular Sciences, 23(1), 479. https://doi.org/10.3390/ijms23010479