
How Cold Shock Affects Ploidy Level and Early Ontogenetic Development of the Sterlet, A. ruthenus L.


Abstract
:1. Introduction
2. Results
3. Discussion
3.1. Embryonic and Larval Developments, the Plasticity of Ploidy Level
3.2. Segregation of Alleles and Parentage Analysis
4. Materials and Methods
4.1. Controlled Reproduction with Cold Shock
4.2. Embryo Incubation and Experimental Larval Rearing
4.3. Ploidy Analysis of Experimental Larvae
4.4. Survival of Embryos and Larvae in Experimental Rearing
4.5. Molecular Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Billard, R.; Lecointre, G. Biology and conservation of sturgeon and paddlefish. Rev. Fish Biol. Fish. 2001, 10, 355–392. [Google Scholar] [CrossRef]
- De la Herrán, R.; Robles, F.; Martínez-Espín, E.; Lorente, J.; Rejón, C.R.; Garrido-Ramos, M.A.; Ruiz Rejón, M. Genetic Identification of Western Mediterranean Sturgeons and its Implication for Conservation. Conserv. Genet. 2004, 5, 545–551. [Google Scholar] [CrossRef]
- Chandra, G.; Fopp-Bayat, D. Trends in aquaculture and conservation of sturgeons: A review of molecular and cytogenetic tools. Rev. Aquac. 2021, 13, 119–137. [Google Scholar] [CrossRef]
- Dettlaff, T.A.; Ginsburg, A.S.; Schmalhausen, O.I. Sturgeon Fishes: Developmental Biology and Aquaculture; Springer: Berlin, Germany, 1993; p. 300. [Google Scholar] [CrossRef]
- Havelka, M.; Hulák, M.; Bailie, D.A.; Prodöhl, P.A.; Flajšhans, M. Extensive genome duplications in sturgeons: New evidence from microsatellite data. J. Appl. Ichthyol. 2013, 29, 704–708. [Google Scholar] [CrossRef]
- Ludwig, A. Identification of Acipenseriformes species in trade. J. Appl. Ichthyol. 2008, 24 (Suppl. 1), 2–19. [Google Scholar] [CrossRef]
- Bronzi, P.; Chebanov, M.; Michaels, J.T.; Wei, Q.; Rosenthal, H.; Gessner, J. Sturgeon meat and caviar production: Global update 2017. J. Appl. Ichthyol. 2019, 35, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Brett, J.R. Energetic responses of salmon to temperature. A study of some thermal relations in the physiology and freshwater ecology of sockeye salmon (Oncorhynchus nerka). Am. Zool. 1971, 11, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Van Eenennaam, J.P.; Linares-Casenave, J.; Deng, X.; Doroshov, S.I. Effect of incubation temperature on green sturgeon embryos, Acipenser medirostris. Environ. Biol. Fishes. 2005, 72, 145–154. [Google Scholar] [CrossRef]
- Güralp, H.; Pocherniaieva, K.; Blecha, M.; Policar, T.; Pšenička, M.; Saito, T. Development, and effect of water temperature on development rate, of pikeperch Sander lucioperca embryos. Theriogenology 2017, 104, 94–104. [Google Scholar] [CrossRef]
- Beitinger, T.L.; Bennett, W.A.; McCauley, R.W. Temperature tolerances of North American freshwater fishes exposed to dynamic changes in temperature. Environ. Biol. Fishes 2000, 58, 237–275. [Google Scholar] [CrossRef]
- Somero, G.N. Linking biogeography to physiology: Evolutionary and acclamatory adjustments of thermal limits. Front. Zool. 2005, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kasimov, R. Ecological and Physiological Peculiarities of Development of Valuable and Commercial Fish in Azerbaijan; Elm: Baku, Azerbaijan, 1987; p. 171. (In Russian) [Google Scholar]
- Wang, Y.L.; Binkowski, F.P.; Doroshov, S.I. Effect of temperature on early development of white and lake sturgeon, Acipenser transmontanus and A. fulvescens. Environ. Biol. Fishes 1985, 14, 43–50. [Google Scholar] [CrossRef]
- Bolker, J.A. Embryology. In Sturgeons and Paddle Fish of North America; LeBreton, G., Beamishand, F., MacKindley, R., Eds.; Kluwer Academic Publishers: Boston, MA, USA, 2004; pp. 134–146. [Google Scholar] [CrossRef]
- Güralp, H.; Pocherniaieva, K.; Blecha, M.; Policar, T.; Pšenička, M.; Saito, T. Early embryonic development in pikeperch (Sander lucioperca) related to micromanipulation. Czech J. Anim. Sci. 2016, 61, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Nam, Y.K.; Kim, D.S. Ploidy status of progeny from the crosses between tetraploid males and diploid females in mud loach (Misgurnus mizolepis). Aquaculture 2004, 236, 575–582. [Google Scholar] [CrossRef]
- Takle, H.; Baeverfjord, G.; Lunde, M.; Kolstad, K.; Andersen, O. The effect of heat and cold exposure on HSP70 expression and development of deformities during embryogenesis of Atlantic salmon (Salmo salar). Aquaculture 2005, 249, 515–524. [Google Scholar] [CrossRef]
- Kim, E.J.; Park, C.; Nam, Y.K. Effects of incubation temperature on the embryonic viability and hatching time in Russian sturgeon (Acipenser gueldenstaedtii). Fish. Aquatic. Sci. 2018, 21, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tiwary, B.K.; Kirubagaran, R.; Ray, A.K. The biology of triploid fish. Rev. Fish Biol. Fish. 2004, 14, 391–402. [Google Scholar] [CrossRef]
- Valenti, R.J. Induced polyploidy in Tilapia aurea (Steindachner) by means of temperature shock treatment. J. Fish Biol. 1975, 7, 519–528. [Google Scholar] [CrossRef]
- Lemoine, H.L., Jr.; Smith, L.T. Polyploidy induced in brook trout by cold shock. Trans. Am. Fish. Soc. 1980, 9, 626–631. [Google Scholar] [CrossRef]
- Wolters, W.R.; Libey, G.S.; Chrisman, C.L. Effect of triploidy on growth and gonad development of channel catfish. Trans. Am. Fish. Soc. 1982, 111, 102–105. [Google Scholar] [CrossRef]
- Arai, K.; Taniura, K.; Zhang, Q. Production of second generation progeny of hexaploid loach. Fish. Sci. 1999, 65, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Devlin, R.H.; Biagi, C.A.; Yesaki, T.Y. Growth, viability and genetic characteristics of GH transgenic coho salmon strains. Aquaculture 2004, 236, 607–632. [Google Scholar] [CrossRef]
- Johnson, R.M.; Shrimpton, J.M.; Heath, J.W.; Heath, D.D. Family, induction methodology and interaction effects on the performance of diploid and triploid Chinook salmon (Oncorhynchus tshawytscha). Aquaculture 2004, 234, 123–142. [Google Scholar] [CrossRef]
- Werner, I.; Viant, M.R.; Rosenblum, E.S.; Gantner, A.S.; Tjeerdema, R.S.; Johnson, M.L. Cellular responses to temperature stress in steelhead trout (Onchorynchus mykiss) parr with different rearing histories. Fish Physiol. Biochem. 2006, 32, 261–273. [Google Scholar] [CrossRef]
- Van Eenennaam, A.L.; Van Eenennaam, J.P.; Medrano, J.F.; Doroshov, S.I. Rapid verification of meiotic gynogenesis and polyploidy in white sturgeon (Acipenser transmontanus Richardson). Aquaculture 1996, 147, 177–189. [Google Scholar] [CrossRef]
- Mims, S.D.; Shelton, W.L. Induced meiotic gynogenesis in shovelnose sturgeon. Aquac. Int. 1998, 6, 323–329. [Google Scholar] [CrossRef]
- Omoto, N.; Maebayashi, M.; Adachi, S.; Arai, K.; Yamauchi, K. Sex ratios of triploids and gynogenetic diploids induced in the hybrid sturgeon, the bester (Huso huso female × Acipenser ruthenus male). Aquaculture 2005, 245, 39–47. [Google Scholar] [CrossRef]
- Fopp-Bayat, D. Verification of meiotic gynogenesis in Siberian sturgeon (Acipenser baeri Brandt) using microsatellite DNA and cytogenetical markers. J. Fish Biol. 2007, 71, 478–485. [Google Scholar] [CrossRef]
- Fopp-Bayat, D. Meiotic gynogenesis revealed not homogametic female sex determination system in Siberian sturgeon (Acipenser baeri Brandt). Aquaculture 2010, 305, 174–177. [Google Scholar] [CrossRef]
- Fopp-Bayat, D.; Kolman, R.; Woznicki, P. Induction of meiotic gynogenesis in sterlet (Acipenser ruthenus) using UV-irradiated bester sperm. Aquaculture 2007, 264, 54–58. [Google Scholar] [CrossRef]
- Fopp-Bayat, D.; Jankun, M.; Woznicki, P.; Kolman, R. Viability of diploid and triploid larvae of Siberian sturgeon and bester hybrids. Aquac. Res. 2007, 38, 1301–1304. [Google Scholar] [CrossRef]
- Fopp-Bayat, D.; Hliwa, P.; Ocalewicz, K. Presence of gynogenetic males suggests a female heterogamety in sterlet Acipenser ruthenus L. Anim. Reprod. Sci. 2018, 189, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Fopp-Bayat, D.; Wlasow, T.; Ziomek, E. Haematology of gynogenetic diploids of Siberien sturgeon Acipenser baerii Brandt. Acta Vet. Brno 2013, 82, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Wlasow, T.; Fopp-Bayat, D. The effect of thermal shock on morphological characteristics of blood cells in Siberian sturgeon (Acipenser baerii) triploids. Acta Vet. Brno 2011, 80, 215–218. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.C.; Wei, Q.W.; Pan, G.B. Induction of meiotic gynogenesis in paddlefish (Polyodon spathula) and its confirmation using microsatellite markers. J. Appl. Ichthyol. 2011, 27, 496–500. [Google Scholar] [CrossRef]
- Saber, M.H.; Noveiri, S.B.; Pourkazemi, M.; Yarmohammadi, M. Induction of gynogenesis in stellate sturgeon (Acipenser stellatus Pallas 1771) and its verification using microsatellite markers. Aquac. Res. 2008, 39, 1483–1487. [Google Scholar] [CrossRef]
- Saber, M.H.; Noveiri, S.B.; Pourkazemi, M.; Yazdani, M.; Ghoroghi, A.; Bahmani, M.; Pourdehghani, M.; Chakmehdouz, F.; Yarmohammadi, M.; Nowruzfashkhami, M. Induction of meiotic gynogenesis in ship sturgeon Acipenser nudiventris using UV-irradiated heterologous sperm. J. Appl. Genet. 2014, 55, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Saber, M.H.; Hallajian, A. Study of sex determination system in ship sturgeon, Acipenser nudiventris using meiotic gynogenesis. Aquac. Int. 2014, 22, 273–279. [Google Scholar] [CrossRef]
- Donaldson, M.R.; Arlinghaus, R.; Hanson, K.C.; Cooke, S.J. Enhancing catch-and-release science with biotelemetry. Fish Fish. 2008, 9, 79–105. [Google Scholar] [CrossRef]
- Hovda, J.; Linley, T.J. The potential application of hypothermia for anesthesia in adult Pacific salmon. N. Am. J. Aquac. 2000, 62, 67–72. [Google Scholar] [CrossRef]
- Craig, J.K.; Foote, C.J.; Wood, C.C. Evidence for temperature-dependent sex determination in sockeye salmon (Oncorhynchus nerka). Can. J. Fish. Aquat. Sci. 1996, 53, 141–147. [Google Scholar] [CrossRef]
- Peruzzi, S.; Kettunen, A.; Primicerio, R.; Kauric, G. Thermal shock induction of triploidy in Atlantic cod (Gadus morhua L.). Aquac. Res. 2007, 38, 926–932. [Google Scholar] [CrossRef]
- Pandian, T.J.; Koteeswaran, R. Ploidy induction and sex control in fish. Hydrobiologia 1998, 384, 167–243. [Google Scholar] [CrossRef]
- Morishima, K.; Oshima, K.; Horie, S.; Fujimoto, T.; Yamaha, E.; Arai, K. Clonal diploid sperm of the diploid-triploid mosaic loach, Misgurnus anguillicaudatus (Teleostei: Cobitidae). J. Exp. Zool. A Comp. Exp. Biol. 2004, 301, 502–511. [Google Scholar] [CrossRef]
- Yoshikawa, H.; Morishima, K.; Fujimoto, T.; Yamaha, E.; Arai, K. Simultaneous formation of haploid, diploid and triploid eggs in diploid-triploid mosaic loaches. J. Fish Biol. 2007, 71 (Suppl. B), 250–263. [Google Scholar] [CrossRef] [Green Version]
- Morishima, K.; Fujimoto, T.; Sato, M.; Kawae, A.; Zhao, Y.; Yamaha, E.; Arai, K. Cold-shock eliminates female nucleus in fertilized eggs to induce androgenesis in the loach (Misgurnus anguillicaudatus), a teleost fish. BMC Biotechnol. 2011, 11, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Teplitz, R.L.; Joyce, J.E.; Doroshov, S.I.; Min, B.H. A preliminary ploidy analysis of diploid and triploid salmonids. Can. J. Fish. Aquat. Sci. 1994, 51, 38–41. [Google Scholar] [CrossRef]
- Hussain, M.G. Suppression of meiotic and mitotic cell divisions in Nile tilapia, Oreochromis niloticus L., and induction of triploids and two types of gynogenetic diploids. Asian Fish. Sci. 1995, 8, 133–142. [Google Scholar]
- Iegorova, V.; Psenicka, M.; Lebeda, I.; Rodina, M.; Saito, T. Polyspermy produces viable haploid/diploid mosaics in sturgeon. Biol. Reprod. 2018, 99, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Gheyas, A.A.; Hussain, M.G. Triploidy Induction in Stinging Catfish Heteropneustes fossilis Using Cold Shock. Reprod. Nutr. Dev. 2001, 14, 323–332. [Google Scholar] [CrossRef]
- Wiggins, T.A.; Bender, T.R.J.R.; Mudrack, V.A.; Coll, J.A. The development, feeding, growth, and survival of cultured American shad larvae through the transition from endogenous to exogenous nutrition. Prog. Fish-Cult. 1985, 47, 87–93. [Google Scholar] [CrossRef]
- Elliott, J.M. The critical-period concept for juvenile survival and its relevance for population regulation in young sea trout. Salmotrutta. J. Fish. Biol. 1989, 35, 91–98. [Google Scholar] [CrossRef]
- Chourrout, D.; Chevassus, B.; Krieg, F.; Happe, A.; Burger, G.; Renard, P. Production of second generation tetraploid males and diploid females—Potential of tetraploid fishes. Theor. Appl. Genet. 1986, 72, 193–206. [Google Scholar] [CrossRef]
- Fopp-Bayat, D.; Nitkiewicz, A.; Chandra, G. First report on embryonic and larval development of 2n/3n mosaic sterlet. Animal 2021, 15, 100317. [Google Scholar] [CrossRef]
- Piferrer, F.; Beaumont, A.; Falguiere, J.C.; Flajšhans, M.; Haffray, P.; Colombo, L. Polyploid fish and shellfish: Production, biology and applications to aquaculture for performance improvement and genetic containment. Aquaculture 2009, 239, 125–156. [Google Scholar] [CrossRef] [Green Version]
- Flajšhans, M.; Havelka, M.; Lebeda, I.; Rodina, M.; Gela, D.; Hubálek, M. Application of hydrostatic pressure shock for retention of the second polar body in sterlet (Acipenser ruthenus). Aquaculture 2020, 520, 734947. [Google Scholar] [CrossRef]
- Nagler, J.J. Polyploidy production in salmonidae. In Sex Control in Aquaculture; Wang, H.-P., Piferrer, F., Chen, S.-L., Eds.; First John Wiley & Sons.: Hoboken, NJ, USA, 2019; pp. 297–304. [Google Scholar]
- Alcantar-Vazquez, J.P. Effects of cold-shock triploidy induction treatment on cell cleavage, hatching rate, and survival of triploid larvae of spotted sand bass (Paralabrax maculatofasciatus). Cienc. Mar. 2016, 42, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Sultana, R.M. Prediction of Larval Viability Based on Egg Quality Parameters and Early Cleavage Patterns in the Experiments of Triploidy Induction in Atlantic cod Gadus morhua L. Master’s Thesis, University of Tromsø, Tromsø, Norway, 27 May 2005. [Google Scholar]
- Diaz, N.F.; Iturra, P.; Veloso, A.; Estay, F.; Colihueque, N. Physiological factors affecting triploid production in rainbow trout, Oncorhynchus mykiss. Aquaculture 1993, 114, 33–40. [Google Scholar] [CrossRef]
- Zelazowska, M. Formation and structure of egg envelopes in Russian sturgeon Acipenser gueldenstaedtii (Acipenseriformes: Acipenseridae). J. Fish Biol. 2010, 76, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Dakin, E.E.; Avise, J.C. Microsatellite null alleles in parentage analysis. Heredity 2004, 93, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Horvath, L.; Szabo, T.; Burke, J. Hatchery testing of GnRH analogue-containing pellets on ovulation in four cyprinid species. Polskie Archiwum. Hydrobiologii 1997, 44, 221–226. [Google Scholar]
- McQuown, E.C.; Sloss, B.L.; Sheehan, R.J.; Rodzen, J.; Tranah, G.J.; May, B. Microsatellite analysis of genetic variation in sturgeon: New primer sequences for Scaphirhynchus and Acipenser. Trans. Am. Fish. Soc. 2000, 129, 1380–1388. [Google Scholar] [CrossRef]
- Fopp-Bayat, D. Application of DNA fingerprint analysis for identification of mixed groups of Siberian sturgeon (Acipenser baeri Brandt). Pol. J. Nat. Sci. 2009, 24, 169–176. [Google Scholar] [CrossRef]
- Fopp-Bayat, D.; Ocalewicz, K. Activation of the albino sterlet Acipenser ruthenus eggs by UV-irradiated bester hybrid spermatozoa to provide gynogenetic progeny. Reprod. Domest. Anim. 2015, 50, 554–559. [Google Scholar] [CrossRef] [PubMed]
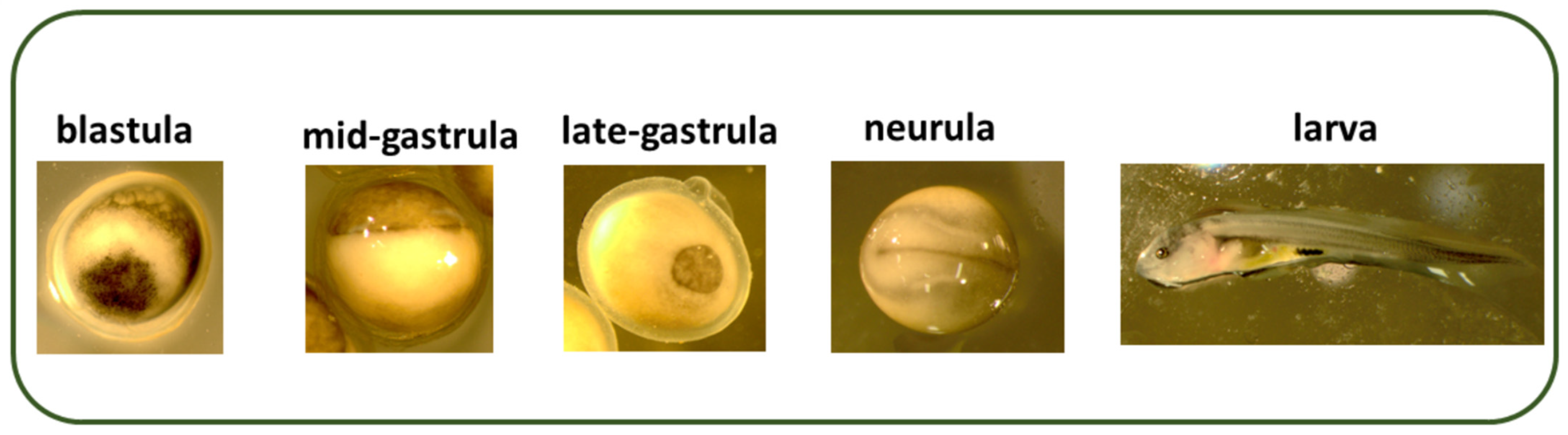
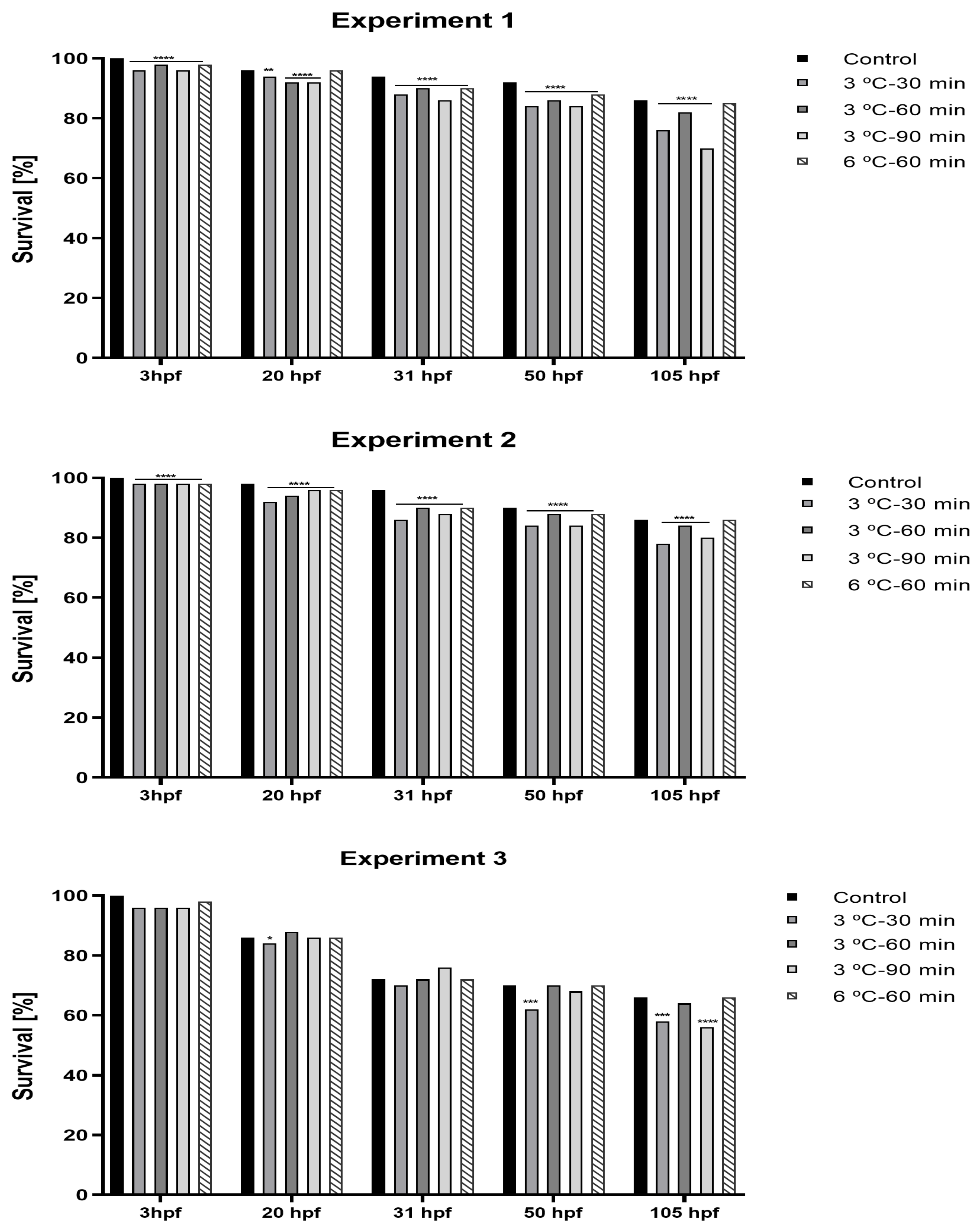
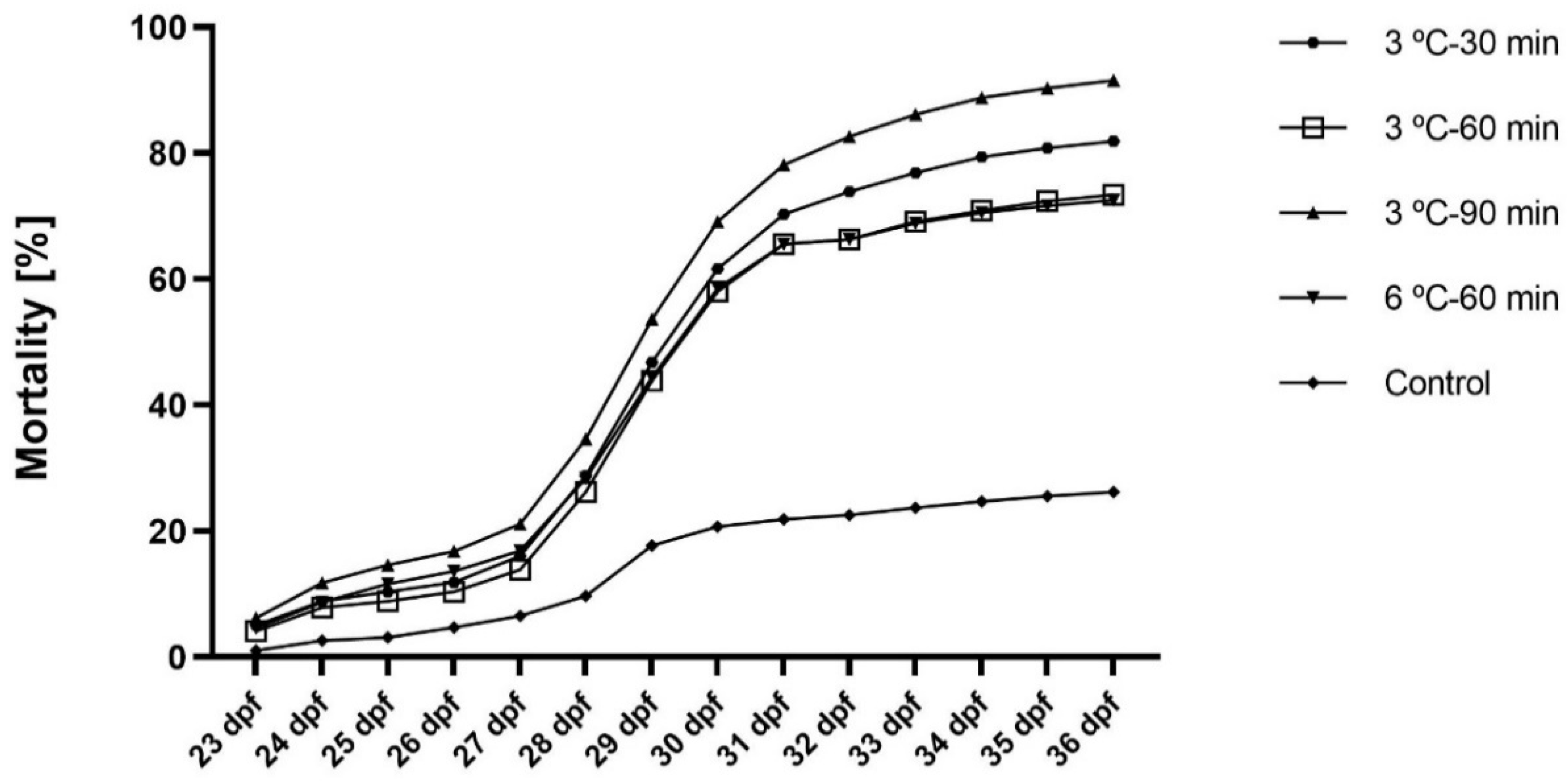




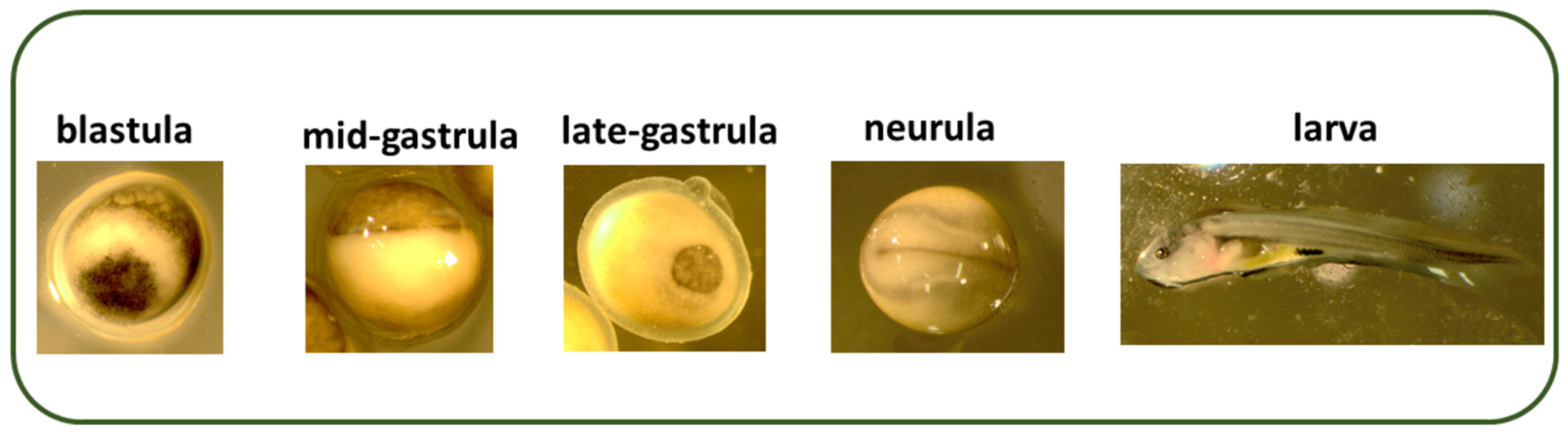{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Experimental Groups | Ploidy Variants (%) | ||||
|---|---|---|---|---|---|---|
| 1n/2n | 2n/3n | 2n | 3n | 1n/2n/3n | ||
| EI | 3 °C–30 min | 20 | - | - | - | 80 |
| 3 °C–60 min | - | 70 | 5 | 25 | - | |
| 3 °C–90 min | - | 80 | 9 | 11 | - | |
| 6 °C–60 min | - | 8 | 92 | - | - | |
| Control | - | - | 100 | - | - | |
| EII | 3 °C–30 min | 30 | - | - | - | 70 |
| 3 °C–60 min | - | 70 | 10 | 20 | - | |
| 3 °C–90 min | - | 80 | 10 | 10 | - | |
| 6 °C–60 min | - | 7 | 93 | - | - | |
| Control | - | - | 100 | - | - | |
| EIII | 3 °C–30 min | - | - | - | - | 100 |
| 3 °C–60 min | - | 100 | - | - | - | |
| 3 °C–90 min | - | 100 | - | - | - | |
| 6 °C–60 min | - | 8 | 92 | - | - | |
| Control | - | - | 100 | - | - | |
| Experimental Groups | Locus | Observed Genotype | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Males—Spermatozoa Donors | Females—Oocyte Donors | Offspring from Experimental Groups | ||||||||
| EI | EII | EIII | EI | EII | EIII | EI | EII | EIII | ||
| 3 °C–30 min | Spl-106 | 234/270 | 234/270 | 260 | 313 | 313/243 | 286/278 | 234/313 (12) | 234/313 (7) | 286/278/260 (30) |
| 270/313 (16) | 234/243 (4) | |||||||||
| 234/270/313 (2) | 270/313 (11) | |||||||||
| 270/243 (2) | ||||||||||
| 234/313/243 (2) | ||||||||||
| 270/313/243 (4) | ||||||||||
| Spl-163 | 188/204 | 188/204 | 188/196 | 188/204 | 188/204 | 204/212 | 188/204 (30) | 188/204 (30) | 212/204/196 (16) | |
| 212/204/188 (14) | ||||||||||
| 3 °C–60 min | Spl-106 | 234/270 | 234/270 | 260 | 313 | 313/243 | 286/278 | 234/313 (13) | 234/313 (5) | 286/278/260 (30) |
| 270/313 (15) | 234/243 (3) | |||||||||
| 234/270/313 (2) | 270/313 (8) | |||||||||
| 270/243 (1) | ||||||||||
| 234/313/243 (6) | ||||||||||
| 270/313/243 (7) | ||||||||||
| Spl-163 | 188/204 | 188/204 | 188/196 | 188/204 | 188/204 | 204/212 | 188/204 (30) | 188/204 (30) | 212/204/196 (15) | |
| 212/204/188 (15) | ||||||||||
| 3 °C–90 min | Spl-106 | 234/270 | 234/270 | 260 | 313 | 313/243 | 286/278 | 234/313 (13) | 234/313 (6) | 286/278/260 (30) |
| 270/313 (16) | 234/243 (2) | |||||||||
| 234/270/313 (1) | 270/313 (8) | |||||||||
| 270/243 (1) | ||||||||||
| 234/313/243 (7) | ||||||||||
| 270/313/243 (6) | ||||||||||
| Spl-163 | 188/204 | 188/204 | 188/196 | 188/204 | 188/204 | 204/212 | 188/204 (11) | 188/204 (10) | 212/204/196 (16) | |
| 188/188 (9) | 188/188 (9) | 212/204/188 (14) | ||||||||
| 204/204 (10) | 204/204 (11) | |||||||||
| 6 °C–60 min | Spl-106 | 234/270 | 234/270 | 260 | 313 | 313/243 | 286/278 | 234/313 (13) | 234/313 (8) | 286/260 (15) |
| 270/313 (16) | 270/313 (12) | 278/260 (13) | ||||||||
| 234/270/313 (1) | 234/243 (4) | 286/278/260 (2) | ||||||||
| 270/243 (3) | ||||||||||
| 234/313/243 (1) | ||||||||||
| 270/313/243 (2) | ||||||||||
| Spl-163 | 188/204 | 188/204 | 188/196 | 188/204 | 188/204 | 204/212 | 188/204 (9) | 188/204 (9) | 188/204 (7) | |
| 188/188 (10) | 188/188 (9) | 188/212 (6) | ||||||||
| 204/204 (11) | 204/204 (12) | 196/204 (8) | ||||||||
| 196/212 (7) | ||||||||||
| 212/204/196 (1) | ||||||||||
| 212/204/188 (1) | ||||||||||
| Control -16 °C | Spl-106 | 234/270 | 234/270 | 260 | 313 | 313/243 | 286/278 | 270/313 (16) | 270/313 (14) | 286/260 (16) |
| 234/313 (14) | 234/313 (9) | 278/260 (14) | ||||||||
| 270/243 (4) | ||||||||||
| 234/243 (3) | ||||||||||
| Spl-163 | 188/204 | 188/204 | 188/196 | 188/204 | 188/204 | 204/212 | 188/204 (10) | 188/204 (10) | 188/204 (8) | |
| 188/188 (9) | 188/188 (9) | 188/212 (7) | ||||||||
| 204/204 (11) | 204/204 (11) | 196/204 (8) | ||||||||
| 196/212 (7) | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fopp-Bayat, D.; Chandra, G.; Nitkiewicz, A. How Cold Shock Affects Ploidy Level and Early Ontogenetic Development of the Sterlet, A. ruthenus L. Int. J. Mol. Sci. 2022, 23, 494. https://doi.org/10.3390/ijms23010494
Fopp-Bayat D, Chandra G, Nitkiewicz A. How Cold Shock Affects Ploidy Level and Early Ontogenetic Development of the Sterlet, A. ruthenus L. International Journal of Molecular Sciences. 2022; 23(1):494. https://doi.org/10.3390/ijms23010494
Chicago/Turabian StyleFopp-Bayat, Dorota, Gyan Chandra, and Anna Nitkiewicz. 2022. "How Cold Shock Affects Ploidy Level and Early Ontogenetic Development of the Sterlet, A. ruthenus L." International Journal of Molecular Sciences 23, no. 1: 494. https://doi.org/10.3390/ijms23010494
APA StyleFopp-Bayat, D., Chandra, G., & Nitkiewicz, A. (2022). How Cold Shock Affects Ploidy Level and Early Ontogenetic Development of the Sterlet, A. ruthenus L. International Journal of Molecular Sciences, 23(1), 494. https://doi.org/10.3390/ijms23010494