
Reciprocal Homer1a and Homer2 Isoform Expression Is a Key Mechanism for Muscle Soleus Atrophy in Spaceflown Mice

 , , , ,
, , , ,  , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Microgravity Exposure Increases Homer1a and Decreases Homer2a/b Transcription in Antigravity Postural Mouse SOL Muscle
2.2. Microgravity Exposure Decreases Homer Protein at the NMJ Postsynaptic Microdomain in Mouse SOL Muscle
2.3. Microgravity Exposure Decreases nAChRs at the NMJ Level in Mouse SOL Muscle
2.4. One to 15 Days of Hindlimb Unloading Caused Transient Increase in Homer1a and a Long-Lasting Decrease in Homer2a/b mRNA Transcripts in Rat SOL Muscle
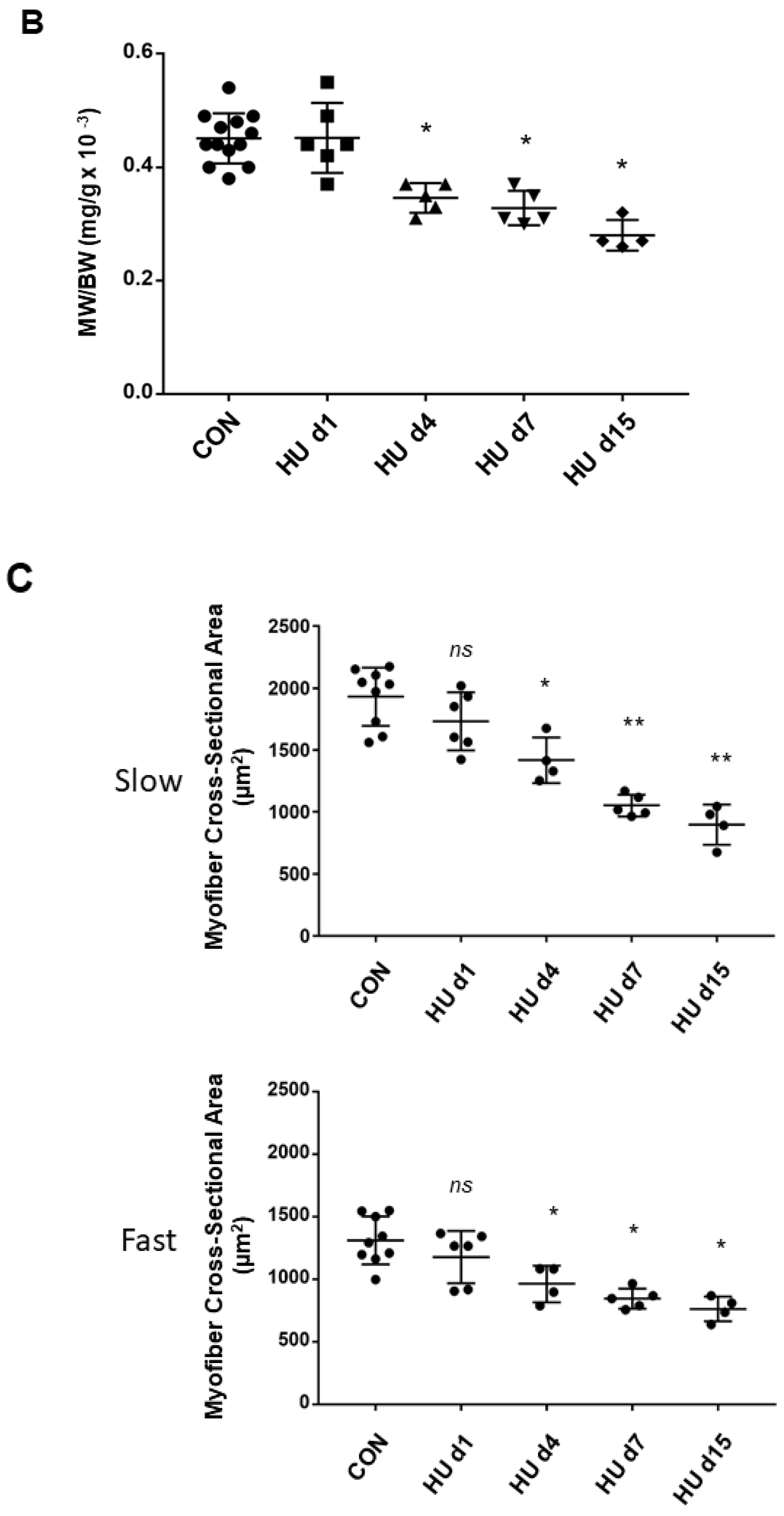
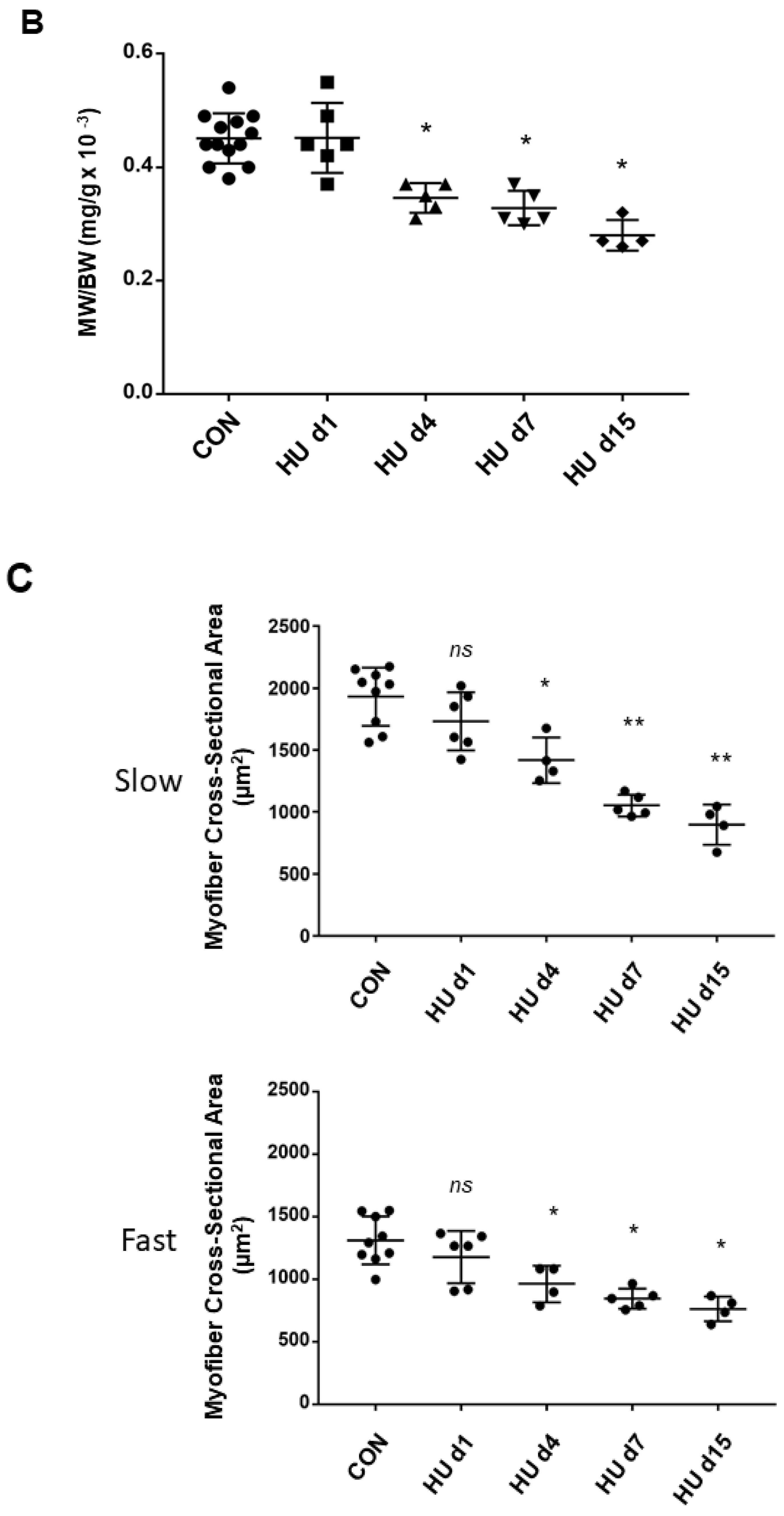
2.5. Muscle Weight/body Weight Ratio Data Changes during HU (rat)
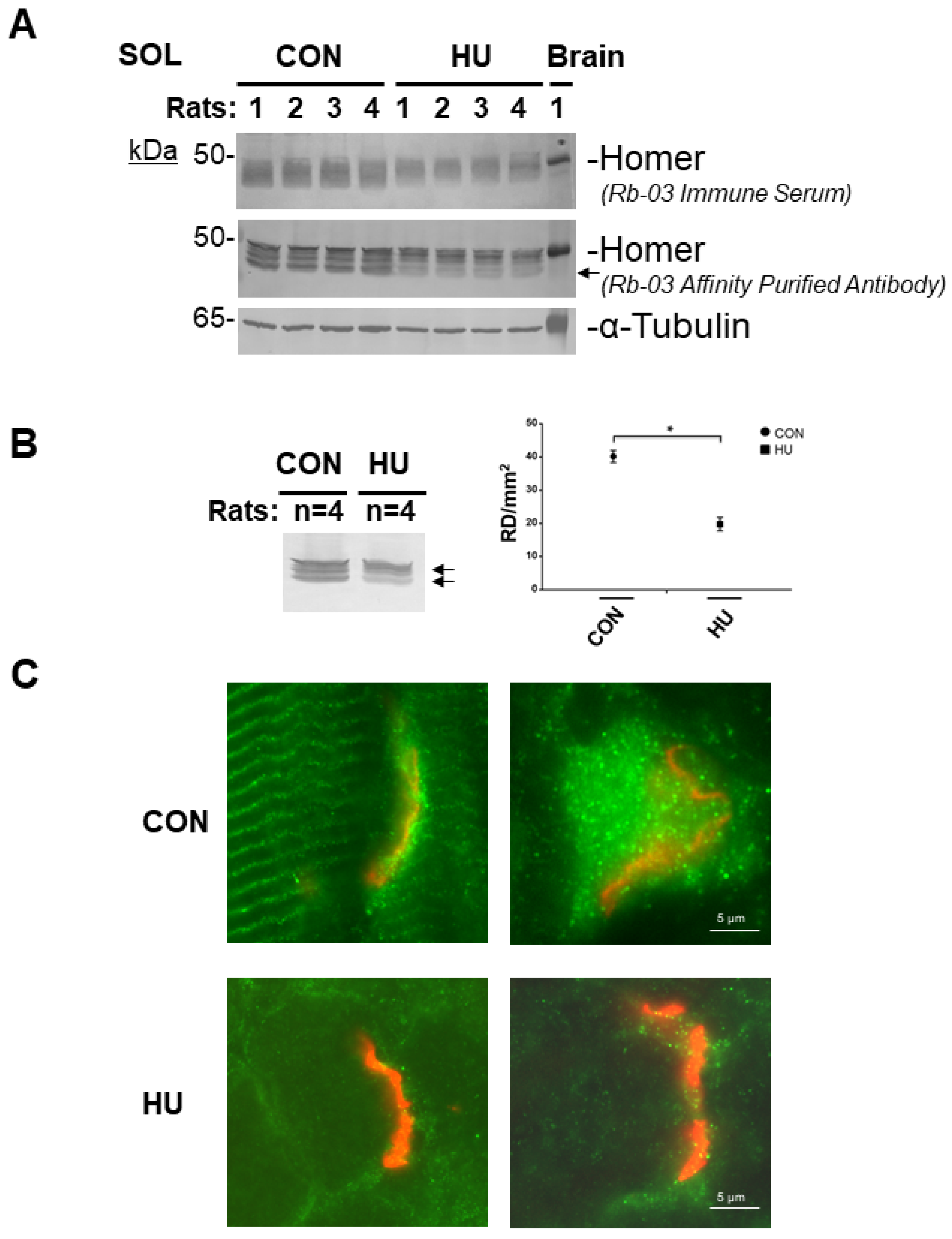
2.6. 3 Weeks of Hindlimb Unloading Decreased Homer Protein Expression and Subcellular Localization at the NMJ of rat SOL Muscle
2.7. Homer Protein Expression and Subcellular Localization in Homer2−/− Mice
3. Discussion
4. Materials and Methods
4.1. Space Flown Animals
4.2. Sample Preparation and Transportation
4.3. Hind Limb Unloading (HU) Experimental Animals and Animal Care
4.4. Homer2−/− Mice
4.5. Homer2 Mice Genotyping
4.6. Immunohistochemistry
4.7. Preparation of Muscle Homogenates
4.8. Sodium Dodecyl Sulphate (SDS)-Polyacrylamide Gel Electrophoresis (PAGE) and Western Blot Analysis
4.9. RNA Extraction and Sample Target Preparation
4.10. Quantitative Polymerase Chain Reaction (qPCR) Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parry, S.M.; Puthucheary, Z.A. The impact of extended bed rest on the musculoskeletal system in the critical care environment. Extrem Physiol. Med. 2015, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Baehr, L.M.; West, D.W.; Marcotte, G.; Marshall, A.G.; De Sousa, L.G.; Baar, K.; Bodine, S.C. Age-related deficits in skeletal muscle recovery following disuse are associated with neuromuscular junction instability and ER stress, not impaired protein synthesis. Aging 2016, 8, 127–146. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Freire, M.; de Cabo, R.; Studenski, S.A.; Ferrucci, L. The Neuromuscular Junction: Aging at the Crossroad between Nerves and Muscle. Front. Aging Neurosci. 2014, 6, 208. [Google Scholar] [CrossRef] [Green Version]
- Van der Pijl, E.M.; van Putten, M.; Niks, E.H.; Verschuuren, J.J.; Aartsma-Rus, A.; Plomp, J.J. Characterization of neuromuscular synapse function abnormalities in multiple Duchenne muscular dystrophy mouse models. Eur. J. Neurosci. 2016, 43, 1623–1635. [Google Scholar] [CrossRef]
- Edgerton, V.R.; Roy, R.R. Neuromuscular adaptation to actual and simulated weightlessness. Adv. Space Biol. Med. 1994, 4, 33–67. [Google Scholar]
- Antonutto, G.; Capelli, C.; Girardis, M.; Zamparo, P.; di Prampero, P.E. Effects of microgravity on maximal power of lower limbs during very short efforts in humans. J. Appl. Physiol. 1999, 86, 85–92. [Google Scholar] [CrossRef]
- McDonald, J.T.; Stainforth, R.; Miller, J.; Cahill, T.; da Silveira, W.A.; Rathi, K.S.; Hardiman, G.; Taylor, D.; Costes, S.V.; Chauhan, V.; et al. NASA GeneLab Platform Utilized for Biological Response to Space Radiation in Animal Models. Cancers 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Scofield, D.C.; Rytlewski, J.D.; Childress, P.; Shah, K.; Tucker, A.; Khan, F.; Peveler, J.; Li, D.; McKinley, T.O.; Chu, T.G.; et al. Development of a step-down method for altering male C57BL/6 mouse housing density and hierarchical structure: Preparations for spaceflight studies. Life Sci. Space Res. 2018, 17, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Radugina, E.A.; Almeida, E.A.C.; Blaber, E.; Poplinskaya, V.A.; Markitantova, Y.V.; Grigoryan, E.N. Exposure to microgravity for 30 days onboard Bion M1 caused muscle atrophy and impaired regeneration in murine femoral Quadriceps. Life Sci. Space Res. 2018, 16, 18–25. [Google Scholar] [CrossRef]
- Baranski, S.; Baranska, W.; Marciniak, M.; Ilyina-Kakueva, E.I. Ultrasonic investigations of the soleus muscle after space flight on the Biosputnik 936. Aviat. Space Environ. Med. 1979, 50, 930–934. [Google Scholar]
- Baranski, S.; Marciniak, M. Morphometric ultrastructural analysis of muscle fibers and neuromuscular junction in rats kept under conditions of weightlessness for 21 days. Mater. Med. Pol. 1990, 22, 258–262. [Google Scholar]
- Pozdniakov, O.M.; Babakova, L.L.; Demorzhi, M.S.; Il’ina-Kakueva, E.I. Changes in the ultrastructure of striated muscles and neuromuscular synapses of rats under the effects of a 13-day space flight. Kosm. Biol. Aviakosm. Med. 1990, 24, 38–42. [Google Scholar]
- Riley, D.A.; Ilyina-Kakueva, E.I.; Ellis, S.; Bain, J.L.; Slocum, G.R.; Sedlak, F.R. Skeletal muscle fiber, nerve, and blood vessel breakdown in space-flown rats. FASEB J. 1990, 4, 84–91. [Google Scholar] [CrossRef]
- D’Amelio, F.; Daunton, N.G. Effects of spaceflight in the adductor longus muscle of rats flown in the Soviet Biosatellite COSMOS 2044. A study employing neural cell adhesion molecule (N-CAM) immunocytochemistry and conventional morphological techniques (light and electron microscopy). J. Neuropathol. Exp. Neurol. 1992, 51, 415–431. [Google Scholar] [CrossRef]
- Deschenes, M.R.; Britt, A.A.; Gomes, R.R.; Booth, F.W.; Gordon, S.E. Recovery of neuromuscular junction morphology following 16 days of spaceflight. Synapse 2001, 42, 177–184. [Google Scholar] [CrossRef]
- Salanova, M.; Bortoloso, E.; Schiffl, G.; Gutsmann, M.; Belavy, D.L.; Felsenberg, D.; Furlan, S.; Volpe, P.; Blottner, D. Expression and regulation of Homer in human skeletal muscle during neuromuscular junction adaptation to disuse and exercise. FASEB J. 2011, 25, 4312–4325. [Google Scholar] [CrossRef]
- Bortoloso, E.; Megighian, A.; Furlan, S.; Gorza, L.; Volpe, P. Homer 2 antagonizes protein degradation in slow-twitch skeletal muscles. Am. J. Physiol. Cell Physiol. 2013, 304, C68–C77. [Google Scholar] [CrossRef] [Green Version]
- Brakeman, P.R.; Lanahan, A.A.; O’Brien, R.; Roche, K.; Barnes, C.A.; Huganir, R.L.; Worley, P.F. Homer: A protein that selectively binds metabotropic glutamate receptors. Nature 1997, 386, 284–288. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, L.; Shi, C.; Wei, R.; Yin, C. Interaction of the Homer1 EVH1 domain and skeletal muscle ryanodine receptor. Biochem. Biophys. Res. Commun. 2019, 514, 720–725. [Google Scholar] [CrossRef]
- Pouliquin, P.; Pace, S.M.; Dulhunty, A.F. In Vitro modulation of the cardiac ryanodine receptor activity by Homer1. Pflugers Arch. 2009, 458, 723–732. [Google Scholar] [CrossRef]
- Tu, J.C.; Xiao, B.; Yuan, J.P.; Lanahan, A.A.; Leoffert, K.; Li, M.; Linden, D.J.; Worley, P.F. Homer binds a novel proline-rich motif and links group 1 metabotropic glutamate receptors with IP3 receptors. Neuron 1998, 21, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Worley, P.F.; Zeng, W.; Huang, G.; Kim, J.Y.; Shin, D.M.; Kim, M.S.; Yuan, J.P.; Kiselyov, K.; Muallem, S. Homer proteins in Ca2+ signaling by excitable and non-excitable cells. Cell Calcium 2007, 42, 363–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kammermeier, P.J.; Worley, P.F. Homer 1a uncouples metabotropic glutamate receptor 5 from postsynaptic effectors. Proc. Natl. Acad. Sci. USA 2007, 104, 6055–6060. [Google Scholar] [CrossRef] [Green Version]
- Clifton, N.E.; Trent, S.; Thomas, K.L.; Hall, J. Regulation and Function of Activity-Dependent Homer in Synaptic Plasticity. Mol. Neuropsychiatry 2019, 5, 147–161. [Google Scholar] [CrossRef]
- Nepliouev, I.; Zhang, Z.S.; Stiber, J.A. Effect of oxidative stress on homer scaffolding proteins. PLoS ONE 2011, 6, e26128. [Google Scholar] [CrossRef] [Green Version]
- Goulding, S.P.; Szumlinski, K.K.; Contet, C.; MacCoss, M.J.; Wu, C.C. A mass spectrometry-based proteomic analysis of Homer2-interacting proteins in the mouse brain. J. Proteomics 2017, 166, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Bottai, D.; Guzowski, J.F.; Schwarz, M.K.; Kang, S.H.; Xiao, B.; Lanahan, A.; Worley, P.F.; Seeburg, P.H. Synaptic activity-induced conversion of intronic to exonic sequence in Homer 1 immediate early gene expression. J. Neurosci. 2002, 22, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Andreev-Andrievskiy, A.; Popova, A.; Boyle, R.; Alberts, J.; Shenkman, B.; Vinogradova, O.; Dolgov, O.; Anokhin, K.; Tsvirkun, D.; Soldatov, P.; et al. Mice in Bion-M 1 space mission: Training and selection. PLoS ONE 2014, 9, e104830. [Google Scholar] [CrossRef] [PubMed]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef]
- Jaspers, S.R.; Tischler, M.E. Atrophy and growth failure of rat hindlimb muscles in tail-cast suspension. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1984, 57, 1472–1479. [Google Scholar] [CrossRef]
- Gambara, G.; Salanova, M.; Ciciliot, S.; Furlan, S.; Gutsmann, M.; Schiffl, G.; Ungethuem, U.; Volpe, P.; Gunga, H.C.; Blottner, D. Gene Expression Profiling in Slow-Type Calf Soleus Muscle of 30 Days Space-Flown Mice. PLoS ONE 2017, 12, e0169314. [Google Scholar] [CrossRef]
- Salanova, M.; Priori, G.; Barone, V.; Intravaia, E.; Flucher, B.; Ciruela, F.; McIlhinney, R.A.; Parys, J.B.; Mikoshiba, K.; Sorrentino, V. Homer proteins and InsP(3) receptors co-localise in the longitudinal sarcoplasmic reticulum of skeletal muscle fibres. Cell Calcium 2002, 32, 193–200. [Google Scholar] [CrossRef]
- Pratt, S.J.P.; Iyer, S.R.; Shah, S.B.; Lovering, R.M. Imaging Analysis of the Neuromuscular Junction in Dystrophic Muscle. Methods Mol. Biol. 2018, 1687, 57–72. [Google Scholar] [PubMed]
- Chiarello, C.; Bortoloso, E.; Carpi, A.; Furlan, S.; Volpe, P. Negative feedback regulation of Homer 1a on norepinephrine-dependent cardiac hypertrophy. Exp. Cell Res. 2013, 319, 1804–1814. [Google Scholar] [CrossRef] [PubMed]
- Lechado, I.T.A.; Vitadello, M.; Traini, L.; Namuduri, A.V.; Gastaldello, S.; Gorza, L. Sarcolemmal loss of active nNOS (Nos1) is an oxidative stress-dependent, early event driving disuse atrophy. J. Pathol. 2018, 246, 433–446. [Google Scholar] [CrossRef]
- Vitadello, M.; Germinario, E.; Ravara, B.; Libera, L.D.; Danieli-Betto, D.; Gorza, L. Curcumin counteracts loss of force and atrophy of hindlimb unloaded rat soleus by hampering neuronal nitric oxide synthase untethering from sarcolemma. J. Physiol. 2014, 592, 2637–2652. [Google Scholar] [CrossRef]
- Sun, L.W.; Blottner, D.; Luan, H.Q.; Salanova, M.; Wang, C.; Niu, H.J.; Felsenberg, D.; Fan, Y.B. Bone and muscle structure and quality preserved by active versus passive muscle exercise on a new stepper device in 21 days tail-suspended rats. J. Musculoskelet Neuronal Interact. 2013, 13, 166–177. [Google Scholar]
- Hwang, S.Y.; Wei, J.; Westhoff, J.H.; Duncan, R.S.; Ozawa, F.; Volpe, P.; Inokuchi, K.; Koulen, P. Differential functional interaction of two Vesl/Homer protein isoforms with ryanodine receptor type 1: A novel mechanism for control of intracellular calcium signaling. Cell Calcium 2003, 34, 177–184. [Google Scholar] [CrossRef]
- Feng, W.; Tu, J.; Yang, T.; Vernon, P.S.; Allen, P.D.; Worley, P.F.; Pessah, I.N. Homer regulates gain of ryanodine receptor type 1 channel complex. J. Biol. Chem. 2002, 277, 44722–44730. [Google Scholar] [CrossRef] [Green Version]
- Stiber, J.A.; Tabatabaei, N.; Hawkins, A.F.; Hawke, T.; Worley, P.F.; Williams, R.S.; Rosenberg, P. Homer modulates NFAT-dependent signaling during muscle differentiation. Dev. Biol. 2005, 287, 213–224. [Google Scholar] [CrossRef]
- Roy, R.R.; Baldwin, K.M.; Edgerton, V.R. Response of the neuromuscular unit to spaceflight: What has been learned from the rat model. Exerc. Sport Sci. Rev. 1996, 24, 399–425. [Google Scholar] [CrossRef]
- Deschenes, M.R.; Wilson, M.H.; Kraemer, W.J. Neuromuscular adaptations to spaceflight are specific to postural muscles. Muscle Nerve 2005, 31, 468–474. [Google Scholar] [CrossRef]
- Fitts, R.H.; Trappe, S.W.; Costill, D.L.; Gallagher, P.M.; Creer, A.C.; Colloton, P.A.; Peters, J.R.; Romatowski, J.G.; Bain, J.L.; Riley, D.A. Prolonged space flight-induced alterations in the structure and function of human skeletal muscle fibres. J. Physiol. 2010, 588, 3567–3592. [Google Scholar] [CrossRef]
- Fitts, R.H.; Riley, D.R.; Widrick, J.J. Functional and structural adaptations of skeletal muscle to microgravity. J. Exp. Biol. 2001, 204, 3201–3208. [Google Scholar] [CrossRef]
- Porseva, V.V.; Shilkin, V.V.; Strelkov, A.A.; Krasnov, I.B.; Masliukov, P.M. Changes in the Neurochemical Composition of Motor Neurons of the Spinal Cord in Mice under Conditions of Space Flight. Bull. Exp. Biol. Med. 2017, 162, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, A.; Nagatomo, F.; Fujino, H.; Kondo, H.; Ohira, Y. Decreased succinate dehydrogenase activity of gamma and alpha motoneurons in mouse spinal cords following 13 weeks of exposure to microgravity. Neurochem. Res. 2013, 38, 2160–2167. [Google Scholar] [CrossRef]
- Pozdniakov, O.M.; Babakova, L.L.; Demorzhi, M.S.; Il’ina-Kakueva, E.I. Changes in the ultrastructure of neuromuscular synapses in rats under the effects of space flight factors. Bull. Exp. Biol. Med. 1988, 105, 752–755. [Google Scholar]
- Willadt, S.; Nash, M.; Slater, C. Age-related changes in the structure and function of mammalian neuromuscular junctions. Ann. N. Y. Acad. Sci. 2018, 1412, 41–53. [Google Scholar] [CrossRef]
- Deschenes, M.R. Motor unit and neuromuscular junction remodeling with aging. Curr. Aging Sci. 2011, 4, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Vitadello, M.; Sorge, M.; Percivalle, E.; Germinario, E.; Danieli-Betto, D.; Turco, E.; Tarone, G.; Brancaccio, M.; Gorza, L. Loss of melusin is a novel, neuronal NO synthase/FoxO3-independent master switch of unloading-induced muscle atrophy. J. Cachexia Sarcopenia Muscle 2020, 11, 802–819. [Google Scholar] [CrossRef]
- Gerbaix, M.; White, H.; Courbon, G.; Shenkman, B.; Gauquelin-Koch, G.; Vico, L. Eight Days of Earth Reambulation Worsen Bone Loss Induced by 1-Month Spaceflight in the Major Weight-Bearing Ankle Bones of Mature Mice. Front. Physiol. 2018, 9, 746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalivas, P.W.; Szumlinski, K.K.; Worley, P. Homer2 gene deletion in mice produces a phenotype similar to chronic cocaine treated rats. Neurotox. Res. 2004, 6, 385–387. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence |
|---|---|---|
| Homer1a | forward reverse | 5′-GAAGTCGCAGGAGAAGATGG-3′ 5′-GAACTTCCATATTTATCCA-3’ |
| Homer2a/b | forward reverse | 5′-CAGTGTATGTGACTCTCCAGCAG-3′ 5′-CTTTGTGGTTGACAATGTCATG-3′ |
| PPIA | forward reverse | 5′-AGCATGTGGTCTTTGGGAAGGTG-3′ 5′-CTTCTTGCTGGTCTTGCCATTCC-3′ |
| PCX | forward reverse | 5′-CCTCTCAGAGCGAGCAGACT-3′ 5′-TAGGGAAGCCGTAGGTGTTG-3′ |
| GAPDH | forward reverse | 5′-CACCATCTTCCAGGAGCGAG-3′ 5′-CCTTCTCCATGGTGGTGAAGAC-3′ |
| ACTB | forward reverse | 5′-CAAACATCCCCCAAAGTTCTAC-3′ 5′-TGAGGGACTTCCTCTAACCACT-3′ |
| Gene | Primer | Sequence |
|---|---|---|
| Homer1a | forward reverse | 5′-CCAGAAAGTATCAATGGGACAGATG-3′ 5′-TGCTGAATTGAATGTGTACCTATGTG-3′ [34] |
| Homer1b/c | forward reverse | 5′-GTGAAGCAGTGGAAGCAACA-3′ 5′-CAGCTCCTGCACTGTCTGAC-3′ [34] |
| Homer2a/b | forward reverse | 5′-TCTTGCTTCTCTGGCTTTGT-3′ 5′-CTGCGTAAACGGCTAAGGTA-3′ [17] |
| PPIA | forward reverse | 5′-AGCATGTGGTCTTTGGGAAGGTG-3′ 5′-CTTCTTGCTGGTCTTGCCATTCC-3′ |
| ACTB | forward reverse | 5′-CAAACATCCCCCAAAGTTCTAC-3′ 5′-TGAGGGACTTCCTCTAACCACT-3′ |
| TBP1 | forward reverse | 5′-TCAAACCCAGAATTGTTCTCC-3′ 5′-AACTATGTGGTCTTCCTGAATCC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blottner, D.; Trautmann, G.; Furlan, S.; Gambara, G.; Block, K.; Gutsmann, M.; Sun, L.-W.; Worley, P.F.; Gorza, L.; Scano, M.; et al. Reciprocal Homer1a and Homer2 Isoform Expression Is a Key Mechanism for Muscle Soleus Atrophy in Spaceflown Mice. Int. J. Mol. Sci. 2022, 23, 75. https://doi.org/10.3390/ijms23010075
Blottner D, Trautmann G, Furlan S, Gambara G, Block K, Gutsmann M, Sun L-W, Worley PF, Gorza L, Scano M, et al. Reciprocal Homer1a and Homer2 Isoform Expression Is a Key Mechanism for Muscle Soleus Atrophy in Spaceflown Mice. International Journal of Molecular Sciences. 2022; 23(1):75. https://doi.org/10.3390/ijms23010075
Chicago/Turabian StyleBlottner, Dieter, Gabor Trautmann, Sandra Furlan, Guido Gambara, Katharina Block, Martina Gutsmann, Lian-Wen Sun, Paul F. Worley, Luisa Gorza, Martina Scano, and et al. 2022. "Reciprocal Homer1a and Homer2 Isoform Expression Is a Key Mechanism for Muscle Soleus Atrophy in Spaceflown Mice" International Journal of Molecular Sciences 23, no. 1: 75. https://doi.org/10.3390/ijms23010075
APA StyleBlottner, D., Trautmann, G., Furlan, S., Gambara, G., Block, K., Gutsmann, M., Sun, L.-W., Worley, P. F., Gorza, L., Scano, M., Lorenzon, P., Vida, I., Volpe, P., & Salanova, M. (2022). Reciprocal Homer1a and Homer2 Isoform Expression Is a Key Mechanism for Muscle Soleus Atrophy in Spaceflown Mice. International Journal of Molecular Sciences, 23(1), 75. https://doi.org/10.3390/ijms23010075