
Oral Administration of Omega-3 Fatty Acids Attenuates Lung Injury Caused by PM2.5 Respiratory Inhalation Simply and Feasibly In Vivo

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Omega-3 Fatty Acids Attenuated PM2.5-Induced Injury on HUVEC and RAW264.7 Cells
2.2. Omega-3 Fatty Acids Attenuated Lung Injury in Mice Exposed to PM2.5
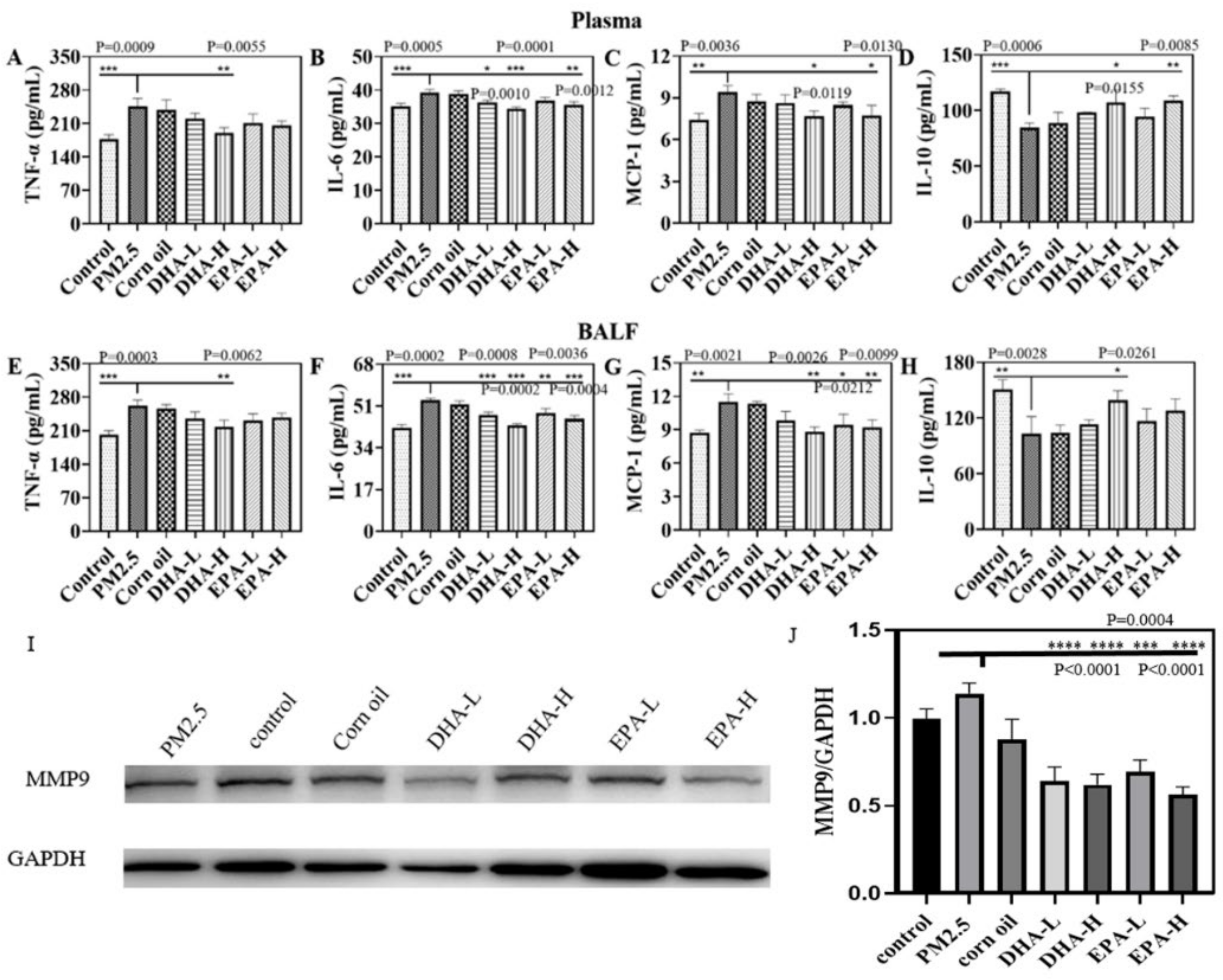
2.3. DHA and EPA Regulated PM2.5-Induced Inflammatory Factors in Mice
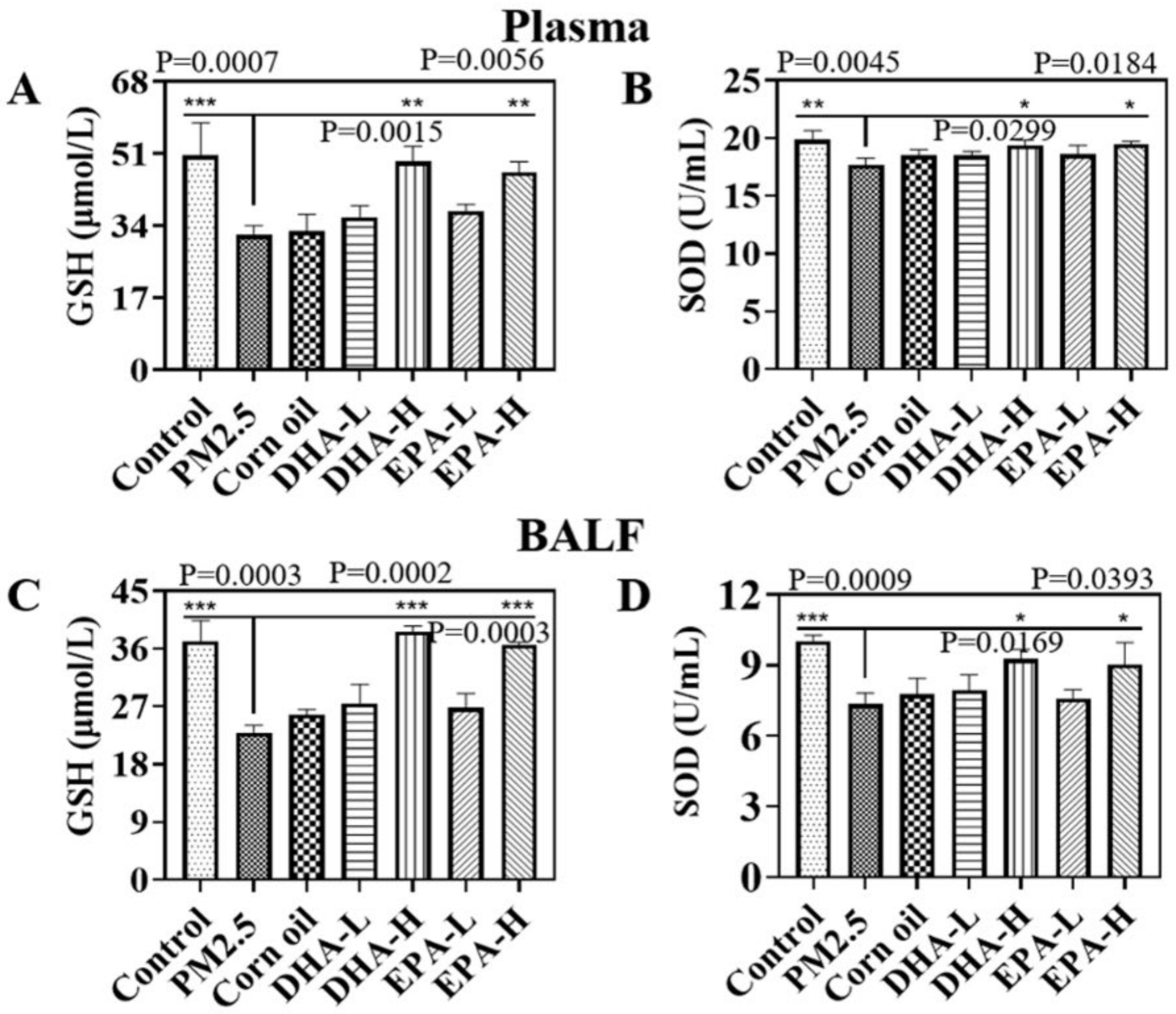
2.4. Omega-3 Fatty Acids Reduced PM2.5-Induced Oxidative Stress Level in Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Collection and Characterization of PM2.5
4.3. Cell Viability Assay
4.4. Endocytosis and Distribution of PM2.5 In Vitro
4.5. Distribution of PM2.5 In Vivo
4.6. Animal Study
4.7. Micro-CT Imaging for the Mice Models
4.8. Biomarkers Detection
4.9. Lung Histopathology
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Q.; Wang, J.; Zhou, J.; Ban, J.; Li, T. Estimation of PM2.5-associated disease burden in China in 2020 and 2030 using population and air quality scenarios: A modelling study. Lancet Planet. Health 2019, 3, e71–e80. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Wang, D.; Fu, Q.; Qiao, L.; Wang, H.; Li, L.; Sun, W.; Li, Q.; Wang, L.; Yang, X.; et al. The effects of firework regulation on air quality and public health during the Chinese Spring Festival from 2013 to 2017 in a Chinese megacity. Environ. Int. 2019, 126, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, J.-N.; Ma, G.-X.; Zhang, Y.-S. China tackles the health effects of air pollution. Lancet 2013, 382, 1959–1960. [Google Scholar] [CrossRef]
- Carvalho-Sousa, C.E.; Pereira, E.P.; Kinker, G.S.; Veras, M.; Ferreira, Z.S.; Barbosa-Nunes, F.P.; Martins, J.O.; Saldiva, P.H.; Reiter, R.J.; Fernandes, P.A.; et al. Immune-pineal axis protects rat lungs exposed to polluted air. J. Pineal Res. 2020, 68, e12636. [Google Scholar] [CrossRef]
- Song, Y.; Qi, Z.; Zhang, Y.; Wei, J.; Liao, X.; Li, R.; Dong, C.; Zhu, L.; Yang, Z.; Cai, Z. Effects of exposure to ambient fine particulate matter on the heart of diet-induced obesity mouse model. Sci. Total Environ. 2020, 732, 139304. [Google Scholar] [CrossRef]
- Dahlem, C.; Kado, S.; He, Y.; Bein, K.; Wu, D.; Haarmann-Stemmann, T.; Kado, N.; Vogel, C. AHR Signaling Interacting with Nutritional Factors Regulating the Expression of Markers in Vascular Inflammation and Atherogenesis. Int. J. Mol. Sci. 2020, 21, 8287. [Google Scholar] [CrossRef]
- Li, Q.; Sun, J.; Chen, X.; Li, S.; Wang, Y.; Xu, C.; Zhao, J.; Zhu, Z.; Tian, L. Integrative characterization of fine particulate matter-induced chronic obstructive pulmonary disease in mice. Sci. Total Environ. 2019, 706, 135687. [Google Scholar] [CrossRef]
- Li, J.; Li, H.; Li, H.; Guo, W.; An, Z.; Zeng, X.; Li, W.; Li, H.; Song, J.; Wu, W. Amelioration of PM2.5-induced lung toxicity in rats by nutritional supplementation with fish oil and Vitamin E. Respir. Res. 2019, 20, 76. [Google Scholar] [CrossRef]
- Xue, Z.; Li, A.; Zhang, X.; Yu, W.; Wang, J.; Li, Y.; Chen, K.; Wang, Z.; Kou, X. Amelioration of PM2.5-induced lung toxicity in rats by nutritional supplementation with biochanin A. Ecotoxicol. Environ. Saf. 2020, 202, 110878. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, N.; Huang, X.; Hu, W.; Ma, Y.; Liang, Y.; Xie, W.; Tang, M. Atmospheric particulate matter impedes autophagic flux by impairing lysosomal milieu and integrity in human umbilical vein endothelial cells (HUVECs). Sci. Total Environ. 2020, 761, 143290. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, H.; Yang, T.; Rui, W.; Liu, F.; Zhang, F.; Zhao, Y.; Ding, W. Direct effects of airborne PM2.5 exposure on macrophage polarizations. Biochim. Biophys. Acta 2016, 1860, 2835–2843. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Tong, L.; Liu, X.; Weng, X.; Chen, X.; Wang, D.; Dudley, S.C.; Weir, E.K.; Ding, W.; Lu, Z.; et al. Short term Pm2.5 exposure caused a robust lung inflammation, vascular remodeling, and exacerbated transition from left ventricular failure to right ventricular hypertrophy. Redox Biol. 2019, 22, 101161. [Google Scholar] [CrossRef] [PubMed]
- Golanski, J.; Szymanska, P.; Rozalski, M. Effects of Omega-3 Polyunsaturated Fatty Acids and Their Metabolites on Haemostasis-Current Perspectives in Cardiovascular Disease. Int. J. Mol. Sci. 2021, 27, 2394. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-H.; Chen, N.-Y.; Tu, P.-H.; Wu, C.-T.; Chiu, S.-C.; Huang, Y.-C.; Lim, S.-N.; Yip, P.K. DHA Attenuates Cerebral Edema Following Traumatic Brain Injury via the Reduction in Blood-Brain Barrier Permeability. Int. J. Mol. Sci. 2020, 21, 6291. [Google Scholar] [CrossRef] [PubMed]
- Manley, B.J.; Makrides, M.; Collins, C.T.; McPhee, A.J.; Gibson, R.A.; Ryan, P.; Sullivan, T.R.; Davis, P.G.; Committee, D.S. High-dose docosahexaenoic acid supplementation of preterm infants: Respiratory and allergy outcomes. Pediatrics 2011, 128, e71–e77. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Hu, X.; Sayer, A.M.; Levy, R.; Zhang, Q.; Xue, Y.; Tong, S.; Bi, J.; Huang, L.; Liu, Y. Satellite-Based Spatiotemporal Trends in PM2.5 Concentrations: China, 2004-2013. Environ. Health Perspect. 2016, 124, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Choi, M.G.; Park, M.K.; Seo, Y.R. Predictive and Prognostic Biomarkers of Respiratory Diseases due to Particulate Matter Exposure. J. Cancer Prev. 2017, 22, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Pope, C.A., III; Burnett, R.T.; Thun, M.J.; Calle, E.E.; Krewski, D.; Ito, K.; Thurston, G.D. Lung cancer, cardiopulmonary mortality, and long-term exposure to fine particulate air pollution. J. Am. Med. Assoc. 2002, 287, 1132–1141. [Google Scholar] [CrossRef] [Green Version]
- Bowler, R.P.; Nicks, M.; Warnick, K.; Crapo, J.D. Role of extracellular superoxide dismutase in bleomycin-induced pulmonary fibrosis. Am. J. Physiol. 2002, 282, L719–L726. [Google Scholar] [CrossRef] [Green Version]
- Manzano-Leon, N.; Serrano-Lomelin, J.; Sanchez, B.N.; Quintana-Belmares, R.; Vega, E.; Vazquez-Lopez, I.; Rojas-Bracho, L.; Lopez-Villegas, M.T.; Vadillo-Ortega, F.; De Vizcaya-Ruiz, A.; et al. TNFalpha and IL-6 Responses to Particulate Matter in Vitro: Variation According to PM Size, Season, and Polycyclic Aromatic Hydrocarbon and Soil Content. Environ. Health Perspect. 2016, 124, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Khawar, M.B.; Abbasi, M.H.; Sheikh, N. IL-32: A Novel Pluripotent Inflammatory Interleukin, towards Gastric Inflammation, Gastric Cancer, and Chronic Rhino Sinusitis. Mediat. Inflamm. 2016, 2016, 8413768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rancan, L.; Paredes, S.D.; Huerta, L.; Casanova, J.; Guzmán, J.; Garutti, I.; González-Aragoneses, F.; Simón, C.; Vara, E. Chemokine Involvement in Lung Injury Secondary to Ischaemia/Reperfusion. Lung 2017, 195, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Zhang, J.; Zeng, H.; Zhang, Y.; Zhang, Y.; Zhou, X.; Zhao, D.; Feng, Y.; Zhou, H. The impact of inflammation and cytokine expression of PM2.5 in AML. Oncol. Lett. 2018, 16, 2732–2740. [Google Scholar] [CrossRef] [Green Version]
- Van Eeden, S.F.; Tan, W.C.; Suwa, T.; Mukae, H.; Terashima, T.; Fujii, T.; Qui, D.; Vincent, R.; Hogg, J.C. Cytokines involved in the systemic inflammatory response induced by exposure to particulate matter air pollutants (PM(10)). Am. J. Respir. Crit. Care Med. 2001, 164, 826–830. [Google Scholar] [CrossRef] [PubMed]
- Pintha, K.; Chaiwangyen, W.; Yodkeeree, S.; Suttajit, M.; Tantipaiboonwong, P. Suppressive Effects of Rosmarinic Acid Rich Fraction from Perilla on Oxidative Stress, Inflammation and Metastasis Ability in A549 Cells Exposed to PM via C-Jun, P-65-Nf-Kappab and Akt Signaling Pathways. Biomolecules 2021, 11, 1090. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Mechanisms of action of (n-3) fatty acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [Green Version]
- Gorjão, R.; Azevedo-Martins, A.K.; Rodrigues, H.G.; Abdulkader, F.; Arcisio-Miranda, M.; Procopio, J.; Curi, R. Comparative effects of DHA and EPA on cell function. Pharmacol. Ther. 2009, 122, 56–64. [Google Scholar] [CrossRef]
- Bruno, M.J.; Koeppe, E.K., III; Andersen, O.S. Docosahexaenoic acid alters bilayer elastic properties. Proc. Natl. Acad. Sci. USA 2007, 104, 9638–9643. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Bhattacharya, A.; Fernandes, G. Docosahexaenoic acid is more potent inhibitor of osteoclast differentiation in RAW 264.7 cells than eicosapentaenoic acid. J. Cell. Physiol. 2007, 214, 201–209. [Google Scholar] [CrossRef]
- Muller-Quernheim, J. MMPs are regulatory enzymes in pathways of inflammatory disorders, tissue injury, malignancies and remodelling of the lung. Eur. Respir. J. 2011, 38, 12–14. [Google Scholar] [CrossRef]
- Xue, H.; Wan, M.; Song, D.; Li, Y.; Li, J. Eicosapentaenoic acid and docosahexaenoic acid modulate mitogen-activated protein kinase activity in endothelium. Vasc. Pharmacol. 2006, 44, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Kong, J.; Chen, K.; Zhang, J.; Chen, Z.; Li, J.; Chang, Y.; Yuan, H.; Shi, X.; Chen, G.; et al. Evaluation of Nano-Particulate-Matter-Induced Lung Injury in Mice Using Quantitative Micro-Computed Tomography. J. Nanosci. Nanotechnol. 2022, 21, 6041. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Mao, M.; Li, J.; Chen, Z.; Ji, Y.; Kong, J.; Wang, Z.; Zhang, J.; Wang, Y.; Liang, W.; et al. Oral Administration of Omega-3 Fatty Acids Attenuates Lung Injury Caused by PM2.5 Respiratory Inhalation Simply and Feasibly In Vivo. Int. J. Mol. Sci. 2022, 23, 5323. https://doi.org/10.3390/ijms23105323
Li J, Mao M, Li J, Chen Z, Ji Y, Kong J, Wang Z, Zhang J, Wang Y, Liang W, et al. Oral Administration of Omega-3 Fatty Acids Attenuates Lung Injury Caused by PM2.5 Respiratory Inhalation Simply and Feasibly In Vivo. International Journal of Molecular Sciences. 2022; 23(10):5323. https://doi.org/10.3390/ijms23105323
Chicago/Turabian StyleLi, Juan, Meiru Mao, Jiacheng Li, Ziteng Chen, Ying Ji, Jianglong Kong, Zhijie Wang, Jiaxin Zhang, Yujiao Wang, Wei Liang, and et al. 2022. "Oral Administration of Omega-3 Fatty Acids Attenuates Lung Injury Caused by PM2.5 Respiratory Inhalation Simply and Feasibly In Vivo" International Journal of Molecular Sciences 23, no. 10: 5323. https://doi.org/10.3390/ijms23105323
APA StyleLi, J., Mao, M., Li, J., Chen, Z., Ji, Y., Kong, J., Wang, Z., Zhang, J., Wang, Y., Liang, W., Liang, H., Lv, L., Liu, Q., Yan, R., Yuan, H., Chen, K., Chang, Y., Chen, G., & Xing, G. (2022). Oral Administration of Omega-3 Fatty Acids Attenuates Lung Injury Caused by PM2.5 Respiratory Inhalation Simply and Feasibly In Vivo. International Journal of Molecular Sciences, 23(10), 5323. https://doi.org/10.3390/ijms23105323