
The Functions of PCNA in Tumor Stemness and Invasion

Abstract
:1. Introduction
2. Results
2.1. Enhanced Mesenchymal Phenotype in Spontaneous Tumors Derived from WT/PyMT Mice but Not in Tumors Developed in 211F/PyMT Mice
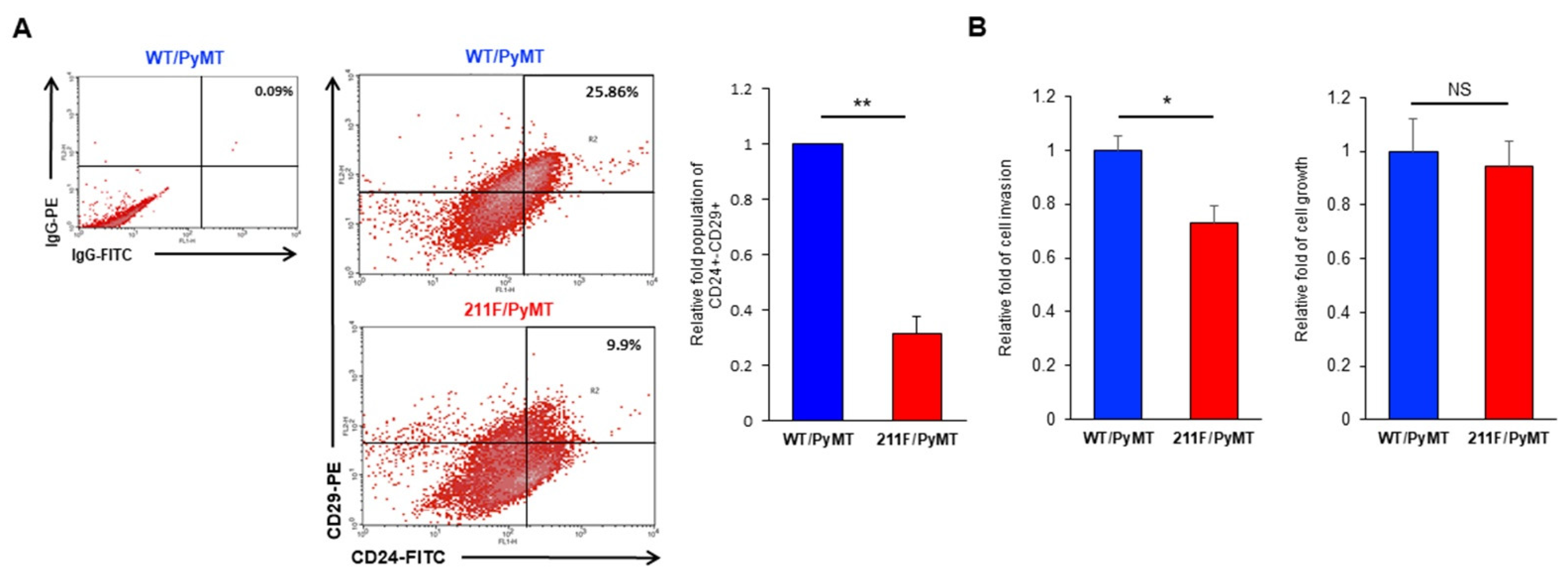
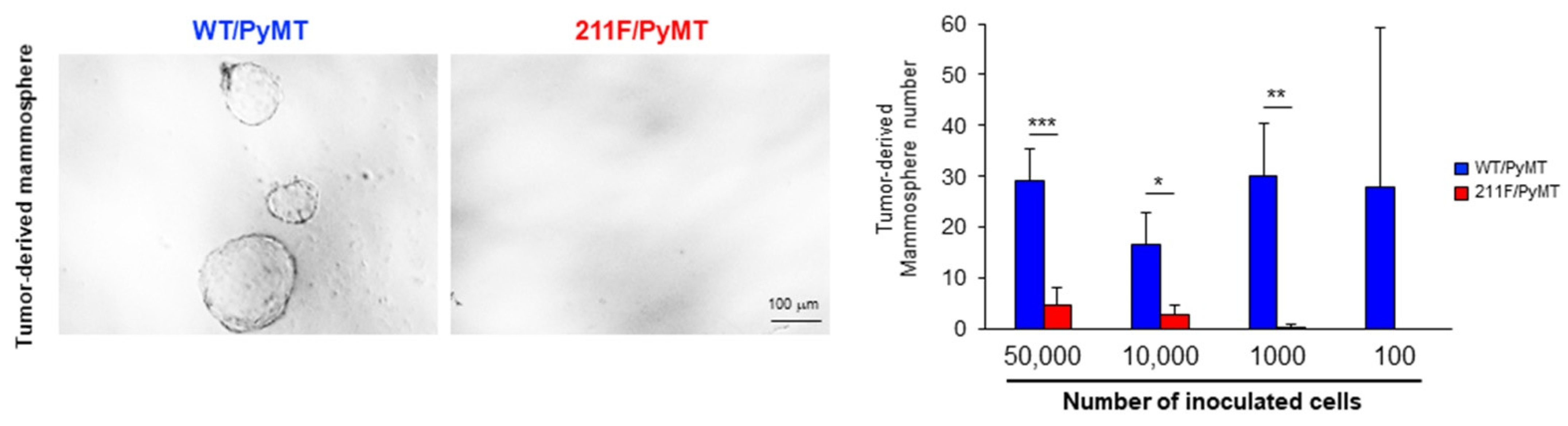
2.2. Y211 Phosphorylation Is Important for Maintaining the Stemness of Cancer Cells
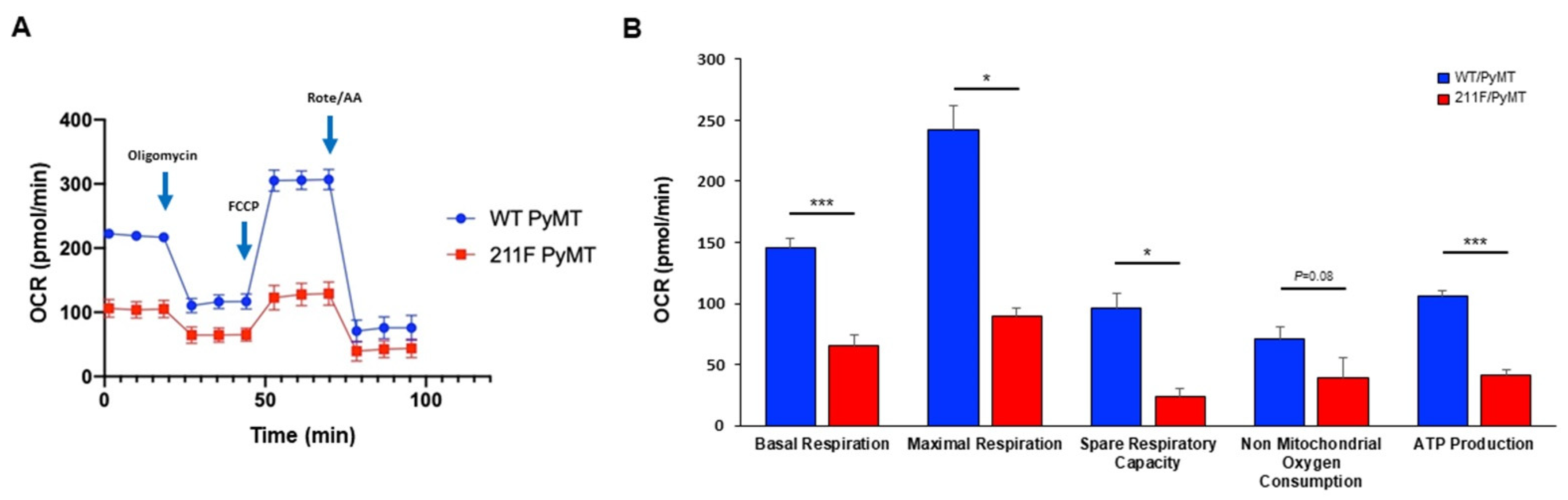
2.3. Metabolism Reprogramming Mediated by Y211 Phosphorylation in PCNA
2.4. MEFs Derived from WT Mice (MFE/WT) Promote Cancer Cell Growth Compared to MEFs Derived from 211F Mice (MEF/211F)
3. Discussion
4. Materials and Methods
4.1. Cell Lines, Chemicals and Antibodies
4.2. Boyden Chamber Assay
4.3. Immunofluorescence
4.4. Limiting Dilution Assay
4.5. ALDH Assay
4.6. Mammosphere Formation Assay
4.7. Oxygen Consumption Rate (OCR) Assay
4.8. Fibroblast Co-Culture Assay
4.9. qRT-PCR
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.X.; Bos, P.D.; Massagué, J. Metastasis: From dissemination to organ-specific colonization. Nat. Rev. Cancer 2009, 9, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.J.; Nieto, M.A. Epithelial-Mesenchymal Transitions in Development and Disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.A.; Guo, W.; Liao, M.-J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The Epithelial-Mesenchymal Transition Generates Cells with Properties of Stem Cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [Green Version]
- Melo, F.d.S.e.; Kurtova, A.V.; Harnoss, J.M.; Kljavin, N.; Hoeck, J.D.; Hung, J.; Anderson, J.E.; Storm, E.E.; Modrusan, Z.; Koeppen, H.; et al. A distinct role for Lgr5+ stem cells in primary and metastatic colon cancer. Nature 2017, 543, 676–680. [Google Scholar] [CrossRef]
- Blanpain, C.; Lowry, W.E.; Geoghegan, A.; Polak, L.; Fuchs, E. Self-Renewal, Multipotency, and the Existence of Two Cell Populations within an Epithelial Stem Cell Niche. Cell 2004, 118, 635–648. [Google Scholar] [CrossRef]
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the maestro of the replication fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef] [Green Version]
- Roa, S.; Avdievich, E.; Peled, J.U.; MacCarthy, T.; Werling, U.; Kuang, F.L.; Kan, R.; Zhao, C.; Bergman, A.; Cohen, P.E.; et al. Ubiquitylated PCNA plays a role in somatic hypermutation and class-switch recombination and is required for meiotic progression. Proc. Natl. Acad. Sci. USA 2008, 105, 16248–16253. [Google Scholar] [CrossRef] [Green Version]
- Choe, K.N.; Moldovan, G.-L. Forging Ahead through Darkness: PCNA, Still the Principal Conductor at the Replication Fork. Mol. Cell 2017, 65, 380–392. [Google Scholar] [CrossRef] [Green Version]
- Maison, C.; Almouzni, G. HP1 and the dynamics of heterochromatin maintenance. Nat. Rev. Mol. Cell Biol. 2004, 5, 296–304. [Google Scholar] [CrossRef]
- Wang, S.-C. PCNA: A silent housekeeper or a potential therapeutic target? Trends Pharmacol. Sci. 2014, 35, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-C.; Nakajima, Y.; Yu, Y.-L.; Xia, W.; Chen, C.-T.; Yang, C.-C.; McIntush, E.W.; Li, L.-Y.; Hawke, D.H.; Kobayashi, R.; et al. Tyrosine phosphorylation controls PCNA function through protein stability. Nat. Cell Biol. 2006, 8, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Lo, Y.-H.; Ma, L.; Waltz, S.E.; Gray, J.K.; Hung, M.-C.; Wang, S.-C. Targeting tyrosine phosphorylation of PCNA inhibits prostate cancer growth. Mol. Cancer Ther. 2011, 10, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Lo, Y.H.; Ho, P.C.; Wang, S.C. Epidermal growth factor receptor protects proliferating cell nuclear antigen from cullin 4A protein-mediated proteolysis. J. Biol. Chem. 2012, 287, 27148–27157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Ho, P.-C.; Lo, Y.-H.; Espejo, A.; Bedford, M.T.; Hung, M.-C.; Wang, S.-C. Interaction of proliferation cell nuclear antigen (PCNA) with c-Abl in cell proliferation and response to DNA damages in breast cancer. PLoS ONE 2012, 7, e29416. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Chen, M.-S.; Lo, Y.-H.; Waltz, S.; Wang, J.; Ho, P.-C.; Vasiliauskas, J.; Plattner, R.; Wang, Y.-L.; Wang, S.-C. The Ron receptor tyrosine kinase activates c-Abl to promote cell proliferation through tyrosine phosphorylation of PCNA in breast cancer. Oncogene 2014, 33, 1429–1437. [Google Scholar] [CrossRef] [Green Version]
- Bass, A.J.; Thorsson, V.; Shmulevich, I.; Reynolds, S.M.; Miller, M.; Bernard, B.; Hinoue, T.; Laird, P.W.; Curtis, C.; Shen, H.; et al. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-L.; Lee, C.-C.; Shen, Y.-C.; Lin, P.-L.; Wu, W.-R.; Lin, Y.-Z.; Cheng, W.-C.; Chang, H.; Hung, Y.; Cho, Y.-C.; et al. Evading immune surveillance via tyrosine phosphorylation of nuclear PCNA. Cell Rep. 2021, 36, 109537. [Google Scholar] [CrossRef]
- Duursma, A.M.; Driscoll, R.; Elias, J.E.; Cimprich, K.A. A role for the MRN complex in ATR activation via TOPBP1 recruitment. Mol. Cell 2013, 2013, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Clarke, M.F.; Dick, J.E.; Dirks, P.B.; Eaves, C.J.; Jamieson, C.H.; Jones, D.L.; Visvader, J.; Weissman, I.L.; Wahl, G.M. Cancer stem cells--perspectives on current status and future directions: AACR Workshop on cancer stem cells. Cancer Res. 2006, 66, 9339–9344. [Google Scholar] [CrossRef] [Green Version]
- Ginestier, C.; Hur, M.H.; Charafe-Jauffret, E.; Monville, F.; Dutcher, J.; Brown, M.; Jacquemier, J.; Viens, P.; Kleer, C.G.; Liu, S.; et al. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell 2007, 1, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dontu, G.; Al-Hajj, M.; Abdallah, W.M.; Clarke, M.F.; Wicha, M.S. Stem cells in normal breast development and breast cancer. Cell Prolif. 2003, 36, 59–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, F.L.; Harrison, H.; Spence, K.; Ablett, M.P.; Simões, B.M.; Farnie, G.; Clarke, R.B. A Detailed Mammosphere Assay Protocol for the Quantification of Breast Stem Cell Activity. J. Mammary Gland. Biol. Neoplasia 2012, 17, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Sancho, P.; Burgos-Ramos, E.; Tavera, A.; Bou Kheir, T.; Jagust, P.; Schoenhals, M.; Barneda, D.; Sellers, K.; Campos-Olivas, R.; Graña, O.; et al. MYC/PGC-1α Balance Determines the Metabolic Phenotype and Plasticity of Pancreatic Cancer Stem Cells. Cell Metab. 2015, 22, 590–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancho, P.; Barneda, D.; Heeschen, C. Hallmarks of cancer stem cell metabolism. Br. J. Cancer 2016, 114, 1305–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, B.; Ortega, J.; Gu, L.; Chang, Z.; Li, G.M. Phosphorylation of proliferating cell nuclear antigen promotes cancer progression by activating the ATM/AKT/GSK3beta/Snail signaling pathway. J. Biol. Chem. 2019, 294, 7037–7045. [Google Scholar] [CrossRef]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.X.; Guan, X.Y.; Fu, L. Therapeutic targeting of the crosstalk between cancer-associated fibroblasts and cancer stem cells. Am. J. Cancer Res. 2019, 9, 1889–1904. [Google Scholar]
- Iwamoto, H.; Izumi, K.; Mizokami, A. Is the C-C Motif Ligand 2–C-C Chemokine Receptor 2 Axis a Promising Target for Cancer Therapy and Diagnosis? Int. J. Mol. Sci. 2020, 21, 9328. [Google Scholar] [CrossRef]
- Wang, X.; Spandidos, A.; Wang, H.; Seed, B. PrimerBank: A PCR primer database for quantitative gene expression analysis, 2012 update. Nucleic Acids Res. 2012, 40, D1144–D1149. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Confidence Intervals for Stem Cell Frequency | ||||||
|---|---|---|---|---|---|---|
| Genotype | Cell # | Tumor Incidence (Response/Test) | Estimate | Lower | Upper | p-Value |
| WT/PyMT | 50,000 10,000 1000 100 | 14/16 8/14 2/16 0/18 | 1/17,228 | 28,660 | 10,356 | 0.000583 |
| 211F/PyMT | 50,000 10,000 1000 100 | 10/16 1/16 0/18 0/18 | 1/62,899 | 113,621 | 34,820 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-L.; Wu, W.-R.; Lin, P.-L.; Shen, Y.-C.; Lin, Y.-Z.; Li, H.-W.; Hsu, K.-W.; Wang, S.-C. The Functions of PCNA in Tumor Stemness and Invasion. Int. J. Mol. Sci. 2022, 23, 5679. https://doi.org/10.3390/ijms23105679
Wang Y-L, Wu W-R, Lin P-L, Shen Y-C, Lin Y-Z, Li H-W, Hsu K-W, Wang S-C. The Functions of PCNA in Tumor Stemness and Invasion. International Journal of Molecular Sciences. 2022; 23(10):5679. https://doi.org/10.3390/ijms23105679
Chicago/Turabian StyleWang, Yuan-Liang, Wan-Rong Wu, Pei-Le Lin, Yi-Chun Shen, You-Zhe Lin, Hong-Wei Li, Kai-Wen Hsu, and Shao-Chun Wang. 2022. "The Functions of PCNA in Tumor Stemness and Invasion" International Journal of Molecular Sciences 23, no. 10: 5679. https://doi.org/10.3390/ijms23105679
APA StyleWang, Y.-L., Wu, W.-R., Lin, P.-L., Shen, Y.-C., Lin, Y.-Z., Li, H.-W., Hsu, K.-W., & Wang, S.-C. (2022). The Functions of PCNA in Tumor Stemness and Invasion. International Journal of Molecular Sciences, 23(10), 5679. https://doi.org/10.3390/ijms23105679