
N-Terminal Peptide of PGLYRP1/Tag7 Is a Novel Ligand for TREM-1 Receptor

 ,
, 
Abstract
:1. Introduction
2. Results
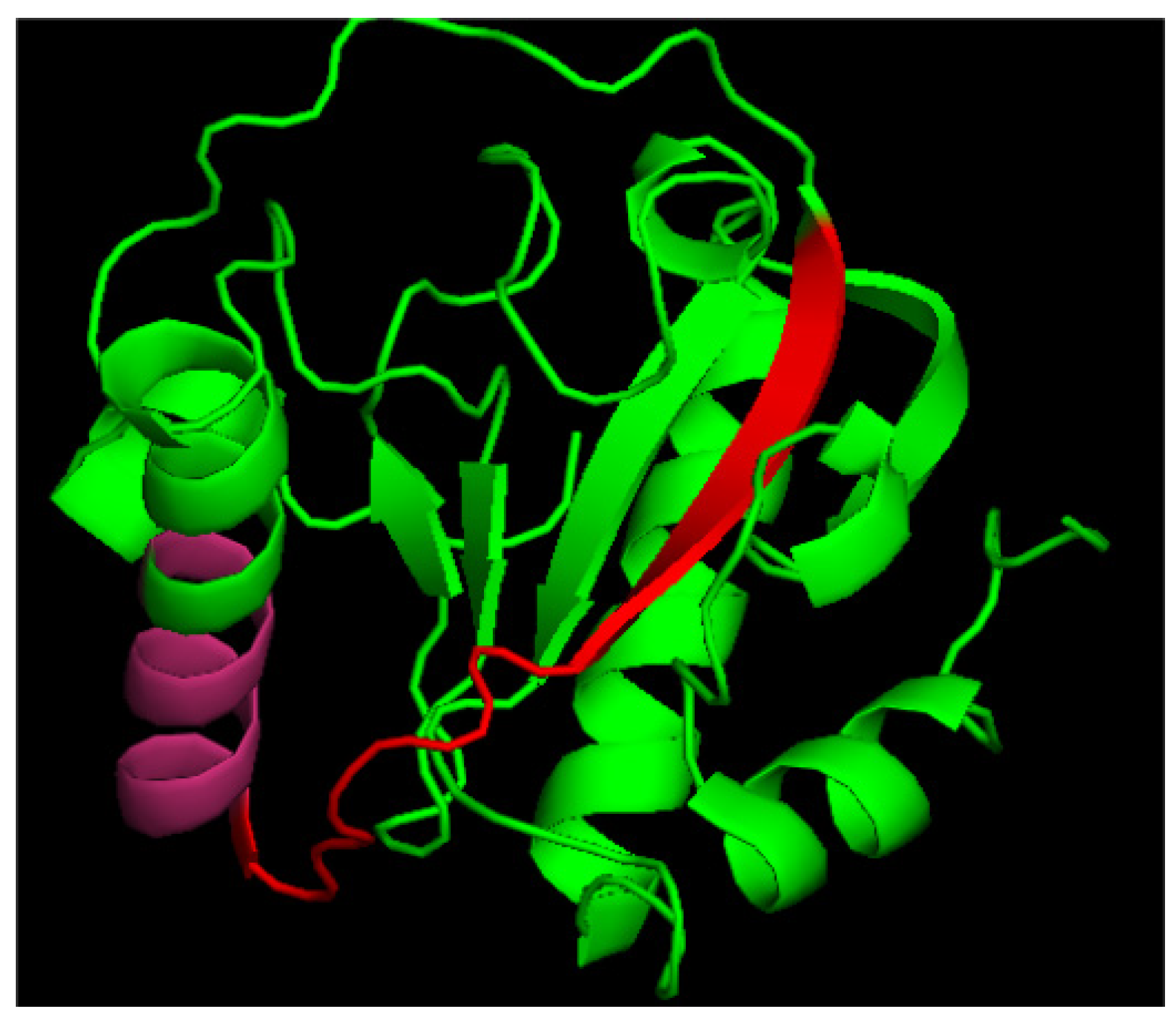
2.1. N-Terminal Peptide of Tag7 Is Responsible for Binding and Activation of TREM-1
2.2. N3 Leads to Expression and Release Cytokines in PBMCs
2.3. Incubation of N3 Peptide with PBMCs Reveals the Appearance of Cytotoxic Lymphocytes
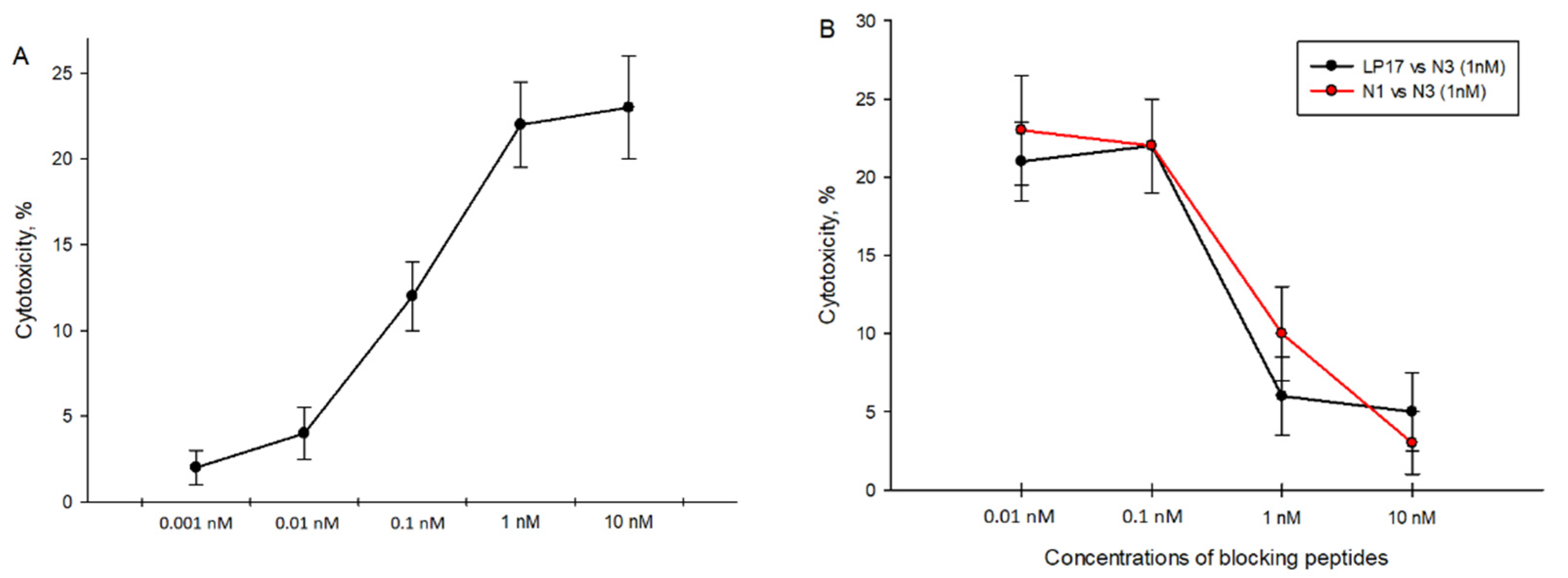
2.4. 9-Mer Peptide of Tag7 Binds to TREM-1 and Prevents Development of Cytotoxic Activity of PBMCs
3. Discussion
4. Materials and Methods
- Cell lines
- Proteins and Antibodies
- Peptides
- Mass Spectrometry
- Affinity Chromatography, Immunoadsorption, and Immunoblotting
- Cytotoxicity assays
- Flow cytometry
- ELISA
- RT-PCR
- Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Li, D.; Wu, M. Pattern Recognition Receptors in Health and Diseases. Signal. Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef] [PubMed]
- Amarante-Mendes, G.P.; Adjemian, S.; Branco, L.M.; Zanetti, L.C.; Weinlich, R.; Bortoluci, K.R. Pattern Recognition Receptors and the Host Cell Death Molecular Machinery. Front Immunol. 2018, 9, 2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafferty, E.I.; Qureshi, S.T.; Schnare, M. The Role of Toll-like Receptors in Acute and Chronic Lung Inflammation. J. Inflamm. 2010, 7, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef] [PubMed]
- Korneev, K.V.; Atretkhany, K.-S.N.; Drutskaya, M.S.; Grivennikov, S.I.; Kuprash, D.V.; Nedospasov, S.A. TLR-Signaling and Proinflammatory Cytokines as Drivers of Tumorigenesis. Cytokine 2017, 89, 127–135. [Google Scholar] [CrossRef]
- Ozato, K.; Tsujimura, H.; Tamura, T. Toll-like Receptor Signaling and Regulation of Cytokine Gene Expression in the Immune System. Biotechniques 2002, (Suppl. 66–68, 70, 72 passim). [Google Scholar] [CrossRef] [Green Version]
- Grassin-Delyle, S.; Abrial, C.; Salvator, H.; Brollo, M.; Naline, E.; Devillier, P. The Role of Toll-Like Receptors in the Production of Cytokines by Human Lung Macrophages. JIN 2020, 12, 63–73. [Google Scholar] [CrossRef]
- Pelham, C.J.; Pandya, A.N.; Agrawal, D.K. Triggering Receptor Expressed on Myeloid Cells (TREM) Receptor Family Modulators: A Patent Review. Expert. Opin. Ther. Pat. 2014, 24, 1383–1395. [Google Scholar] [CrossRef] [Green Version]
- Moriyama, M.; Hirano, T.; Kawano, T.; Kadowaki, Y.; Kodama, S.; Suzuki, M. Toll-like Receptor 4 Plays an Important Role to Enhance Bacterial Clearance from the Nose in Synergy with Triggering Receptor Expressed on Myeloid Cells (TREM)-1 Expression on Polymorphonuclear Neutrophils. Int. J. Pediatr. Otorhinolaryngol. 2018, 112, 27–33. [Google Scholar] [CrossRef]
- Carrasco, K.; Boufenzer, A.; Jolly, L.; Le Cordier, H.; Wang, G.; Heck, A.J.; Cerwenka, A.; Vinolo, E.; Nazabal, A.; Kriznik, A.; et al. TREM-1 Multimerization Is Essential for Its Activation on Monocytes and Neutrophils. Cell Mol. Immunol. 2019, 16, 460–472. [Google Scholar] [CrossRef]
- Knapp, S.; Gibot, S.; de Vos, A.; Versteeg, H.H.; Colonna, M.; van der Poll, T. Cutting Edge: Expression Patterns of Surface and Soluble Triggering Receptor Expressed on Myeloid Cells-1 in Human Endotoxemia. J. Immunol. 2004, 173, 7131–7134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibot, S. Clinical Review: Role of Triggering Receptor Expressed on Myeloid Cells-1 during Sepsis. Crit. Care 2005, 9, 485–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haselmayer, P.; Daniel, M.; Tertilt, C.; Salih, H.R.; Stassen, M.; Schild, H.; Radsak, M.P. Signaling Pathways of the TREM-1- and TLR4-Mediated Neutrophil Oxidative Burst. J. Innate. Immun. 2009, 1, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Tammaro, A.; Derive, M.; Gibot, S.; Leemans, J.C.; Florquin, S.; Dessing, M.C. TREM-1 and Its Potential Ligands in Non-Infectious Diseases: From Biology to Clinical Perspectives. Pharmacology. Ther. 2017, 177, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, Q.; Huang, J.; Jin, Q.; Xu, B.; Chen, F.; Tu, C. Morin Hydrate Inhibits TREM-1/TLR4-Mediated Inflammatory Response in Macrophages and Protects against Carbon Tetrachloride-Induced Acute Liver Injury in Mice. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Dantas, P.H.D.S.; Matos, A.d.O.; da Silva Filho, E.; Silva-Sales, M.; Sales-Campos, H. Triggering Receptor Expressed on Myeloid Cells-1 (TREM-1) as a Therapeutic Target in Infectious and Noninfectious Disease: A Critical Review. Int. Rev. Immunol. 2020, 39, 188–202. [Google Scholar] [CrossRef]
- Palazzo, S.J.; Simpson, T.; Schnapp, L.M. Triggering Receptor Expressed on Myeloid Cells Type 1 as a Potential Therapeutic Target in Sepsis. Dimens. Crit. Care Nurs. 2012, 31, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Jolly, L.; Carrasco, K.; Salcedo-Magguilli, M.; Garaud, J.-J.; Lambden, S.; van der Poll, T.; Mebazaa, A.; Laterre, P.-F.; Gibot, S.; Boufenzer, A.; et al. STREM-1 Is a Specific Biomarker of TREM-1 Pathway Activation. Cell Mol. Immunol. 2021, 18, 2054–2056. [Google Scholar] [CrossRef]
- Weiss, G.; Lai, C.; Fife, M.E.; Grabiec, A.M.; Tildy, B.; Snelgrove, R.J.; Xin, G.; Lloyd, C.M.; Hussell, T. Reversal of TREM-1 Ectodomain Shedding and Improved Bacterial Clearance by Intranasal Metalloproteinase Inhibitors. Mucosal. Immunol. 2017, 10, 1021–1030. [Google Scholar] [CrossRef]
- De Sá Resende, A.; Matos de Oliveira, Y.L.; Rodrigues de Moura, T.; Martins-Filho, P.R. Potential Role of Triggering Receptor Expressed on Myeloid Cells-1 (TREM-1) in SARS-CoV-2 Infection: First Insights. EXCLI J. 2021, 20, 722–723. [Google Scholar] [CrossRef]
- Van Singer, M.; Brahier, T.; Ngai, M.; Wright, J.; Weckman, A.M.; Erice, C.; Meuwly, J.-Y.; Hugli, O.; Kain, K.C.; Boillat-Blanco, N. COVID-19 Risk Stratification Algorithms Based on STREM-1 and IL-6 in Emergency Department. J. Allergy. Clin. Immunol. 2021, 147, 99–106.e4. [Google Scholar] [CrossRef] [PubMed]
- De Nooijer, A.H.; Grondman, I.; Lambden, S.; Kooistra, E.J.; Janssen, N.A.F.; Kox, M.; Pickkers, P.; Joosten, L.A.B.; van de Veerdonk, F.L.; Derive, M.; et al. Increased STREM-1 Plasma Concentrations Are Associated with Poor Clinical Outcomes in Patients with COVID-19. Biosci. Rep. 2021, 41, BSR20210940. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, J.; Salcedo, R.; Mivechi, N.F.; Trinchieri, G.; Horuzsko, A. The Proinflammatory Myeloid Cell Receptor TREM-1 Controls Kupffer Cell Activation and Development of Hepatocellular Carcinoma. Cancer Res. 2012, 72, 3977–3986. [Google Scholar] [CrossRef] [Green Version]
- Sharapova, T.N.; Romanova, E.A.; Ivanova, O.K.; Yashin, D.V.; Sashchenko, L.P. Hsp70 Interacts with the TREM-1 Receptor Expressed on Monocytes and Thereby Stimulates Generation of Cytotoxic Lymphocytes Active against MHC-Negative Tumor Cells. Int. J. Mol. Sci. 2021, 22, 6889. [Google Scholar] [CrossRef] [PubMed]
- Read, C.B.; Kuijper, J.L.; Hjorth, S.A.; Heipel, M.D.; Tang, X.; Fleetwood, A.J.; Dantzler, J.L.; Grell, S.N.; Kastrup, J.; Wang, C.; et al. Cutting Edge: Identification of Neutrophil PGLYRP1 as a Ligand for TREM-1. J. Immunol. 2015, 194, 1417–1421. [Google Scholar] [CrossRef] [PubMed]
- El Mezayen, R.; El Gazzar, M.; Seeds, M.C.; McCall, C.E.; Dreskin, S.C.; Nicolls, M.R. Endogenous Signals Released from Necrotic Cells Augment Inflammatory Responses to Bacterial Endotoxin. Immunol. Lett. 2007, 111, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Kustikova, O.S.; Kiselev, S.L.; Borodulina, O.R.; Senin, V.M.; Afanas’eva, A.V.; Kabishev, A.A. Cloning of the tag7 gene expressed in metastatic mouse tumors. Genetika 1996, 32, 621–628. [Google Scholar]
- Kiselev, S.L.; Kustikova, O.S.; Korobko, E.V.; Prokhortchouk, E.B.; Kabishev, A.A.; Lukanidin, E.M.; Georgiev, G.P. Molecular Cloning and Characterization of the Mouse Tag7 Gene Encoding a Novel Cytokine. J. Biol. Chem. 1998, 273, 18633–18639. [Google Scholar] [CrossRef] [Green Version]
- Sashchenko, L.P.; Dukhanina, E.A.; Yashin, D.V.; Shatalov, Y.V.; Romanova, E.A.; Korobko, E.V.; Demin, A.V.; Lukyanova, T.I.; Kabanova, O.D.; Khaidukov, S.V.; et al. Peptidoglycan Recognition Protein Tag7 Forms a Cytotoxic Complex with Heat Shock Protein 70 in Solution and in Lymphocytes. J. Biol. Chem. 2004, 279, 2117–2124. [Google Scholar] [CrossRef] [Green Version]
- Yashin, D.V.; Ivanova, O.K.; Soshnikova, N.V.; Sheludchenkov, A.A.; Romanova, E.A.; Dukhanina, E.A.; Tonevitsky, A.G.; Gnuchev, N.V.; Gabibov, A.G.; Georgiev, G.P.; et al. Tag7 (PGLYRP1) in Complex with Hsp70 Induces Alternative Cytotoxic Processes in Tumor Cells via TNFR1 Receptor*. J. Biological. Chem. 2015, 290, 21724–21731. [Google Scholar] [CrossRef] [Green Version]
- Sashchenko, L.P.; Dukhanina, E.A.; Shatalov, Y.V.; Yashin, D.V.; Lukyanova, T.I.; Kabanova, O.D.; Romanova, E.A.; Khaidukov, S.V.; Galkin, A.V.; Gnuchev, N.V.; et al. Cytotoxic T Lymphocytes Carrying a Pattern Recognition Protein Tag7 Can Detect Evasive, HLA-Negative but Hsp70-Exposing Tumor Cells, Thereby Ensuring FasL/Fas-Mediated Contact Killing. Blood 2007, 110, 1997–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharapova, T.N.; Romanova, E.A.; Ivanova, O.K.; Sashchenko, L.P.; Yashin, D.V. Cytokines TNFα, IFNγ and IL-2 Are Responsible for Signal Transmission from the Innate Immunity Protein Tag7 (PGLYRP1) to Cytotoxic Effector Lymphocytes. Cells 2020, 9, 2602. [Google Scholar] [CrossRef] [PubMed]
- Sharapova, T.N.; Ivanova, O.K.; Soshnikova, N.V.; Romanova, E.A.; Sashchenko, L.P.; Yashin, D.V. Innate Immunity Protein Tag7 Induces 3 Distinct Populations of Cytotoxic Cells That Use Different Mechanisms to Exhibit Their Antitumor Activity on Human Leukocyte Antigen-Deficient Cancer Cells. J. Innate. Immun. 2017, 9, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Sharapova, T.N.; Romanova, E.A.; Chernov, A.S.; Minakov, A.N.; Kazakov, V.A.; Kudriaeva, A.A.; Belogurov, A.A.; Ivanova, O.K.; Gabibov, A.G.; Telegin, G.B.; et al. Protein PGLYRP1/Tag7 Peptides Decrease the Proinflammatory Response in Human Blood Cells and Mouse Model of Diffuse Alveolar Damage of Lung through Blockage of the TREM-1 and TNFR1 Receptors. Int. J. Mol. Sci. 2021, 22, 11213. [Google Scholar] [CrossRef]
- Telegin, G.B.; Chernov, A.S.; Kazakov, V.A.; Romanova, E.A.; Sharapova, T.N.; Yashin, D.V.; Gabibov, A.G.; Sashchenko, L.P. A 8-Mer Peptide of PGLYRP1/Tag7 Innate Immunity Protein Binds to TNFR1 Receptor and Inhibits TNFα-Induced Cytotoxic Effect and Inflammation. Front. Immunol. 2021, 12, 622471. [Google Scholar] [CrossRef]
- Romanova, E.A.; Sharapova, T.N.; Telegin, G.B.; Minakov, A.N.; Chernov, A.S.; Ivanova, O.K.; Bychkov, M.L.; Sashchenko, L.P.; Yashin, D.V. A 12-Mer Peptide of Tag7 (PGLYRP1) Forms a Cytotoxic Complex with Hsp70 and Inhibits TNF-Alpha Induced Cell Death. Cells 2020, 9, 488. [Google Scholar] [CrossRef] [Green Version]
- Radaev, S.; Kattah, M.; Rostro, B.; Colonna, M.; Sun, P.D. Crystal Structure of the Human Myeloid Cell Activating Receptor TREM-1. Structure 2003, 11, 1527–1535. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fractions | Cytotoxicity % | Fractions | Cytotoxicity % |
|---|---|---|---|
| 1 | 2 ± 0.5 | 13 | 5 ± 1.7 |
| 2 | 3 ± 1.2 | 14 | 4 ± 1.4 |
| 3 | 5 ± 1.6 | 15 | 24 ± 3.1 |
| 4 | 2 ± 0.7 | 16 | 1 ± 0.6 |
| 5 | 4 ± 1.3 | 17 | 2 ± 1.2 |
| 6 | 1 ± 0.5 | 18 | 2 ± 1.3 |
| 7 | 6 ± 1.9 | 19 | 4 ± 1.8 |
| 8 | 5 ± 1.8 | 20 | 6 ± 2.1 |
| 9 | 2 ± 0.5 | 21 | 5 ± 2.0 |
| 10 | 2 ± 0.8 | 22 | 1 ± 0.6 |
| 11 | 3 ± 1.4 | 23 | 2 ± 0.5 |
| 12 | 2 ± 1.7 | 24 | 2 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharapova, T.N.; Ivanova, O.K.; Romanova, E.A.; Sashchenko, L.P.; Yashin, D.V. N-Terminal Peptide of PGLYRP1/Tag7 Is a Novel Ligand for TREM-1 Receptor. Int. J. Mol. Sci. 2022, 23, 5752. https://doi.org/10.3390/ijms23105752
Sharapova TN, Ivanova OK, Romanova EA, Sashchenko LP, Yashin DV. N-Terminal Peptide of PGLYRP1/Tag7 Is a Novel Ligand for TREM-1 Receptor. International Journal of Molecular Sciences. 2022; 23(10):5752. https://doi.org/10.3390/ijms23105752
Chicago/Turabian StyleSharapova, Tatiana N., Olga K. Ivanova, Elena A. Romanova, Lidia P. Sashchenko, and Denis V. Yashin. 2022. "N-Terminal Peptide of PGLYRP1/Tag7 Is a Novel Ligand for TREM-1 Receptor" International Journal of Molecular Sciences 23, no. 10: 5752. https://doi.org/10.3390/ijms23105752
APA StyleSharapova, T. N., Ivanova, O. K., Romanova, E. A., Sashchenko, L. P., & Yashin, D. V. (2022). N-Terminal Peptide of PGLYRP1/Tag7 Is a Novel Ligand for TREM-1 Receptor. International Journal of Molecular Sciences, 23(10), 5752. https://doi.org/10.3390/ijms23105752