
A Possible Way to Relate the Effects of SARS-CoV-2-Induced Changes in Transferrin to Severe COVID-19-Associated Diseases

 ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
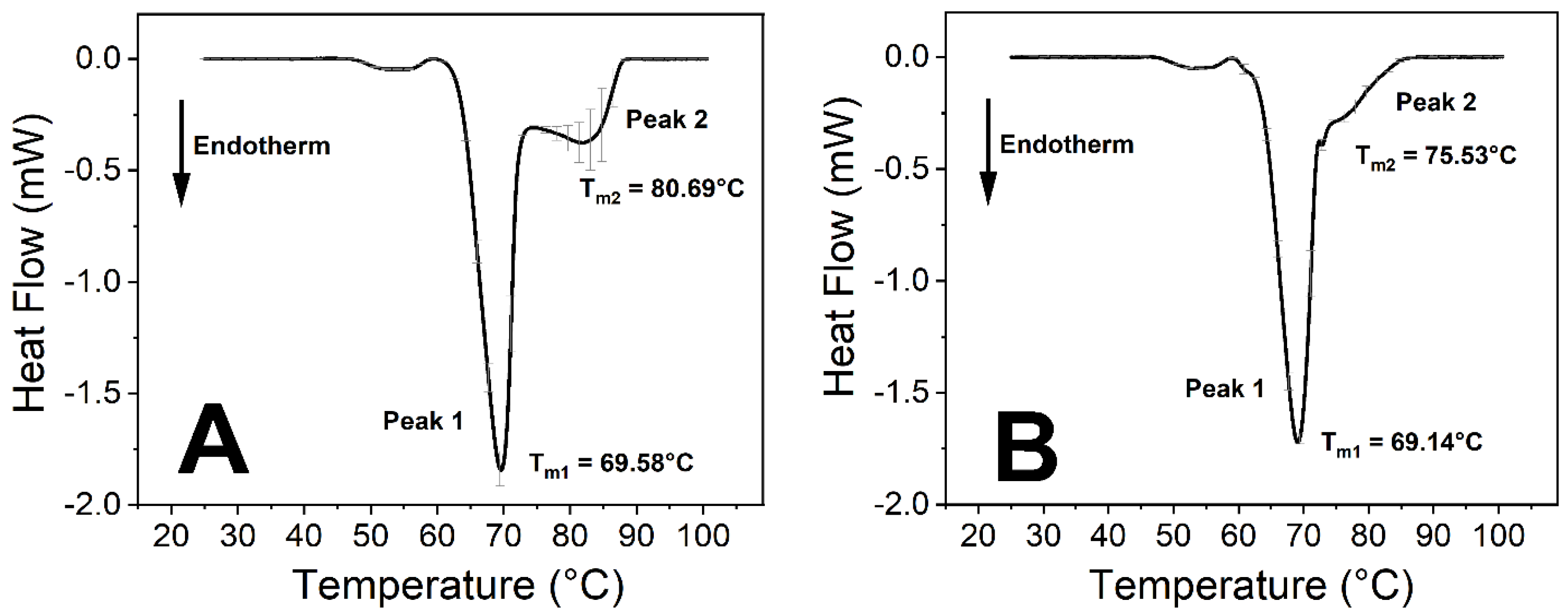
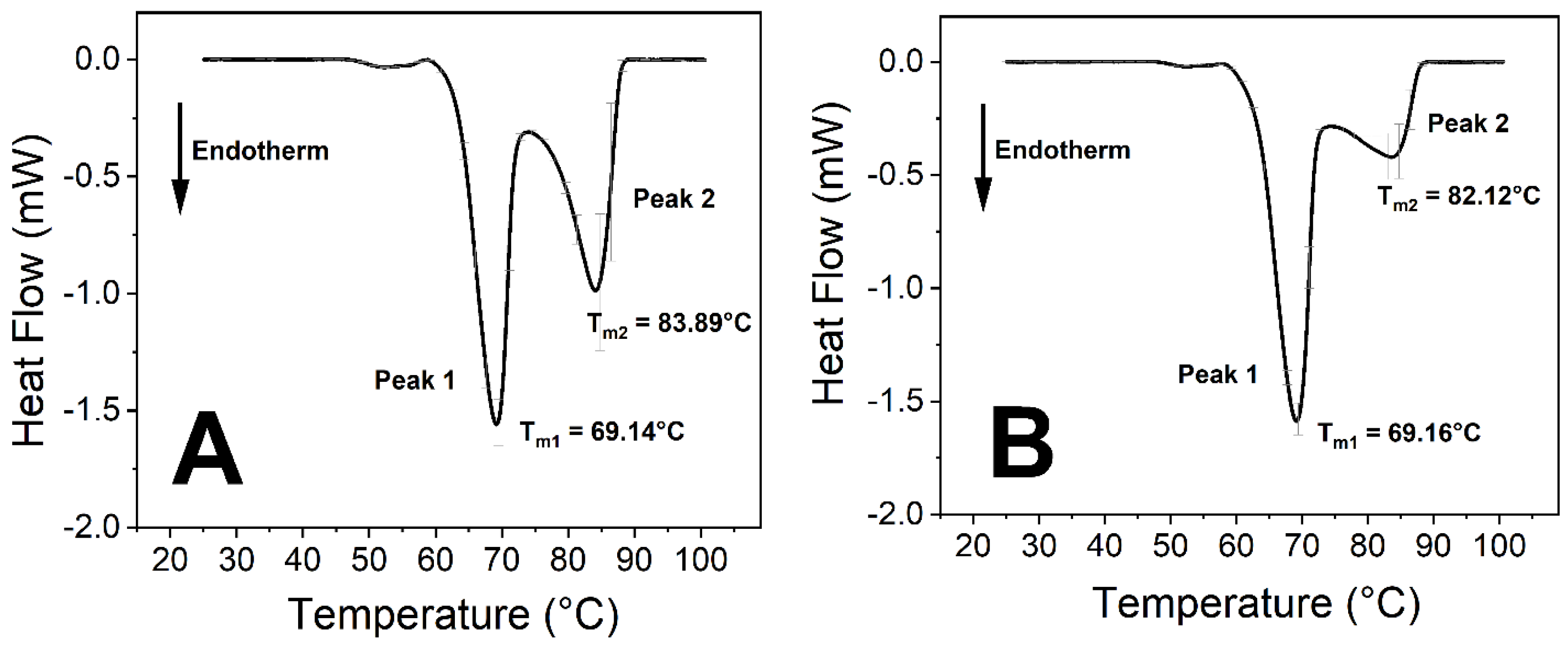
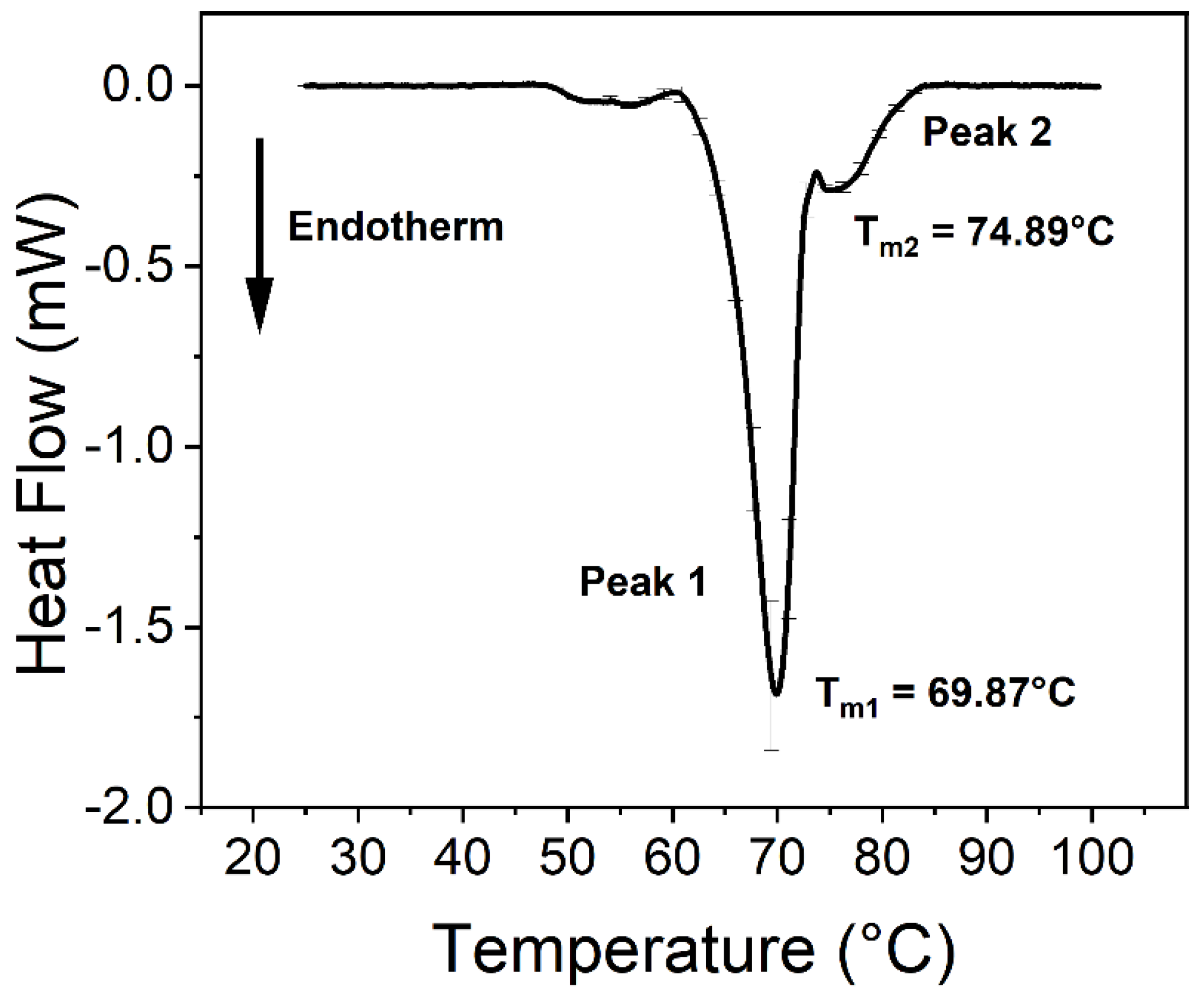
SARS-CoV-2 Influences the Thermodynamic Properties of Transferrin
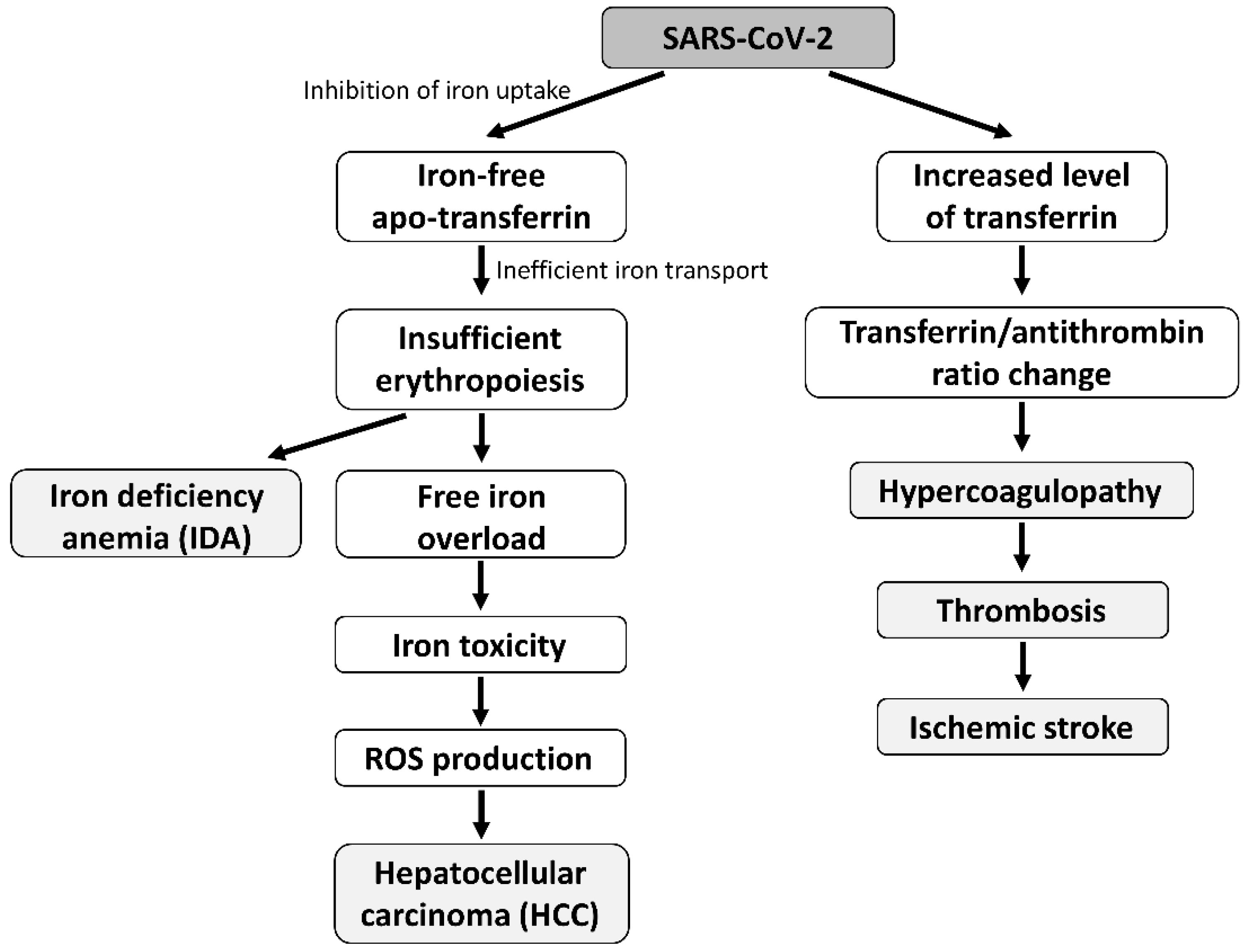
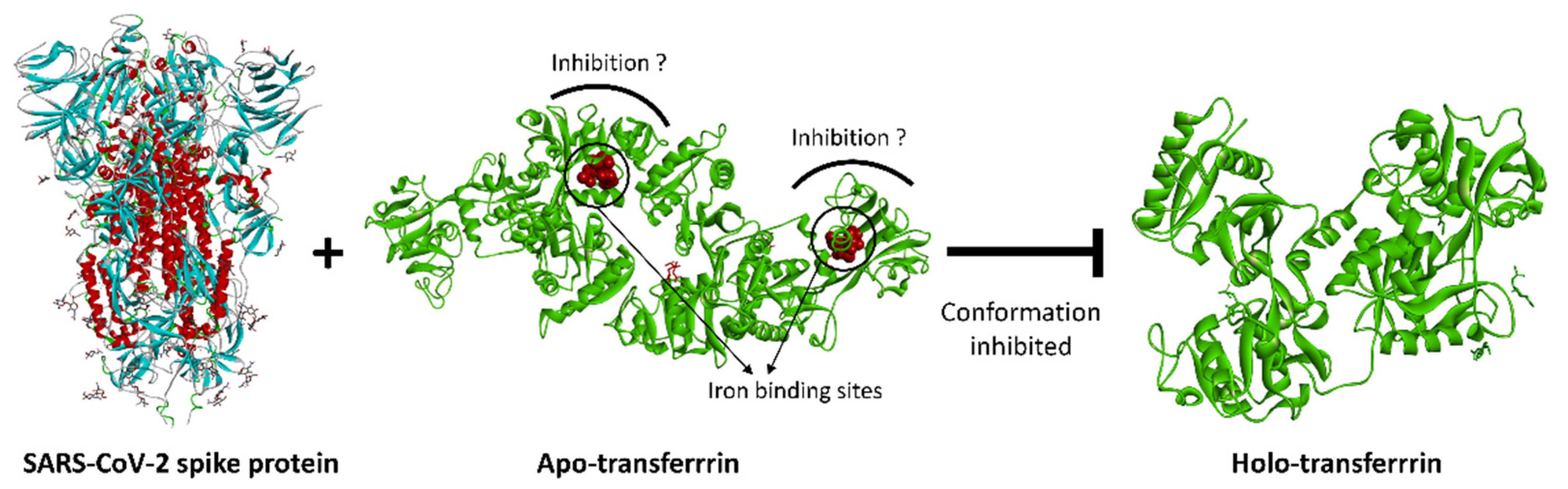
3. Discussion
4. Materials and Methods
4.1. Sample Preparation
4.2. Differential Scanning Calorimetry
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar]
- Kim, M.K.; Lee, B.; Choi, Y.Y.; Um, J.; Lee, K.S.; Sung, H.K.; Kim, Y.; Park, J.S.; Lee, M.; Jang, H.C.; et al. Clinical Characteristics of 40 Patients Infected With the SARS-CoV-2 Omicron Variant in Korea. J. Korean Med. Sci. 2022, 37, e31. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Tzou, P.L.; Nouhin, J.; Gupta, R.K.; de Oliveira, T.; Kosakovsky Pond, S.L.; Fera, D.; Shafer, R.W. The biological and clinical significance of emerging SARS-CoV-2 variants. Nat. Rev. Genet. 2021, 22, 757–773. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.; Ni, Z.; Hu, Y.; Liang, W.; Ou, C.; He, J.; Liu, L.; Shan, H.; Lei, C.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Marchandot, B.; Sattler, L.; Jesel, L.; Matsushita, K.; Schini-Kerth, V.; Grunebaum, L.; Morel, O. COVID-19 Related Coagulopathy: A Distinct Entity? J. Clin. Med. 2020, 9, 1651. [Google Scholar] [CrossRef]
- Iba, T.; Levy, J.H.; Levi, M.; Thachil, J. Coagulopathy in COVID-19. J. Thromb. Haemost. 2020, 18, 2103–2109. [Google Scholar] [CrossRef]
- Tang, N.; Li, D.; Wang, X.; Sun, Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 844–847. [Google Scholar] [CrossRef] [Green Version]
- Fan, B.E.; Chong, V.C.L.; Chan, S.S.W.; Lim, G.H.; Lim, K.G.E.; Tan, G.B.; Mucheli, S.S.; Kuperan, P.; Ong, K.H. Hematologic parameters in patients with COVID-19 infection. Am. J. Hematol. 2020, 95, E131–E134. [Google Scholar]
- Taneri, P.E.; Gómez-Ochoa, S.A.; Llanaj, E.; Raguindin, P.F.; Rojas, L.Z.; Roa-Díaz, Z.M.; Salvador, D.; Groothof, D.; Minder, B.; Kopp-Heim, D.; et al. Anemia and iron metabolism in COVID-19: A systematic review and meta-analysis. Eur. J. Epidemiol. 2020, 35, 763–773. [Google Scholar] [CrossRef]
- Waite, A.A.C.; Hamilton, D.O.; Pizzi, R.; Ageno, W.; Welters, I.D. Hypercoagulopathy in Severe COVID-19: Implications for Acute Care. Thromb. Haemost. 2020, 120, 1654–1667. [Google Scholar] [CrossRef]
- Tang, X.; Fang, M.; Cheng, R.; Zhang, Z.; Wang, Y.; Shen, C.; Han, Y.; Lu, Q.; Du, Y.; Liu, Y.; et al. Iron-deficiency and estrogen are associated with ischemic stroke by up-regulating transferrin to induce hypercoagulability. Circ. Res. 2020, 127, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Torti, S.V.; Torti, F.M. Iron and cancer: More ore to be mined. Nat. Rev. Cancer 2013, 13, 342–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beguin, Y.; Aapro, M.; Ludwig, H.; Mizzen, L.; Österborg, A. Epidemiological and nonclinical studies investigating effects of iron in carcinogenesis-A critical review. Crit. Rev. Oncol. Hematol. 2014, 89, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Moukhadder, H.M.; Halawi, R.; Cappellini, M.D.; Taher, A.T. Hepatocellular carcinoma as an emerging morbidity in the thalassemia syndromes: A comprehensive review. Cancer 2017, 123, 751–758. [Google Scholar] [CrossRef]
- Miyanishi, K.; Tanaka, S.; Sakamoto, H.; Kato, J. The role of iron in hepatic inflammation and hepatocellular carcinoma. Free Radic Biol. Med. 2019, 133, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Banchini, F.; Cattaneo, G.M.; Capelli, P. Serum ferritin levels in inflammation: A retrospective comparative analysis between COVID-19 and emergency surgical non-COVID-19 patients. World J. Emerg. Surg. 2021, 16, 9. [Google Scholar] [CrossRef]
- Worwood, M. Ferritin. Blood Rev. 1990, 4, 259–269. [Google Scholar] [CrossRef]
- Richard, C.; Verdier, F. Transferrin receptors in erythropoiesis. Int. J. Mol. Sci. 2020, 21, 9713. [Google Scholar] [CrossRef]
- Chen, T.; Wu, D.; Chen, H.; Yan, W.; Yang, D.; Chen, G.; Ma, K.; Xu, D.; Yu, H.; Wang, H.; et al. Clinical characteristics of 113 deceased patients with coronavirus disease 2019: Retrospective study. BMJ 2020, 368, m1091. [Google Scholar] [CrossRef] [Green Version]
- Richardson, S.; Hirsch, J.S.; Narasimhan, M.; Crawford, J.M.; McGinn, T.; Davidson, K.W.; Barnaby, D.P.; Becker, L.B.; Chelico, J.D.; Cohen, S.L.; et al. Presenting Characteristics, Comorbidities, and Outcomes among 5700 Patients Hospitalized with COVID-19 in the New York City Area. JAMA—J. Am. Med. Assoc. 2020, 323, 2052–2059. [Google Scholar] [CrossRef]
- Bojkova, D.; Klann, K.; Koch, B.; Widera, M.; Krause, D.; Ciesek, S.; Cinatl, J.; Münch, C. Proteomics of SARS-CoV-2-infected host cells reveals therapy targets. Nature 2020, 583, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Zakin, M.M.; Baron, B.; Guillou, F. Regulation of the tissue-specific expression of transferrin gene. Dev. Neurosci. 2002, 24, 222–226. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, K.M.; Bechtel, M.; Bojkova, D.; Münch, C.; Ciesek, S.; Wass, M.N.; Michaelis, M.; Cinatl, J. COVID-19-related coagulopathy-is transferrin a missing link? Diagnostics 2020, 10, 539. [Google Scholar] [CrossRef]
- Luck, A.N.; Mason, A.B. Transferrin-Mediated Cellular Iron Delivery. Curr. Top. Membr. 2012, 69, 3–35. [Google Scholar] [PubMed] [Green Version]
- Kawabata, H. Transferrin and transferrin receptors update. Free Radic. Biol. Med. 2019, 133, 46–54. [Google Scholar] [CrossRef]
- Benjamín-Rivera, J.A.; Cardona-Rivera, A.E.; Vázquez-Maldonado, Á.L.; Dones-Lassalle, C.Y.; Pabón-Colon, H.L.; Rodríguez-Rivera, H.M.; Rodríguez, I.; González-Espiet, J.C.; Pazol, J.; Pérez-Ríos, J.D.; et al. Exploring serum transferrin regulation of nonferric metal therapeutic function and toxicity. Inorganics 2020, 8, 48. [Google Scholar] [CrossRef]
- Byrne, S.L.; Mason, A.B. Human serum transferrin: A tale of two lobes. Urea gel and steady state fluorescence analysis of recombinant transferrins as a function of pH, time, and the soluble portion of the transferrin receptor. J. Biol. Inorg. Chem. 2009, 14, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Lin, N.; Mason, A.B.; Woodworth, R.C.; Brandts, J.F. Calorimetric Studies of Serum Transferrin and Ovotransferrin. Estimates of Domain Interactions, and Study of the Kinetic Complexities of Ferric Ion Binding. Biochemistry 1994, 33, 1881–1888. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, Z.; Fang, M.; Han, Y.; Wang, G.; Wang, S.; Xue, M.; Li, Y.; Zhang, L.; Wu, J.; et al. Transferrin plays a central role in coagulation balance by interacting with clotting factors. Cell Res. 2020, 30, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Borges do Nascimento, I.J.; Cacic, N.; Abdulazeem, H.M.; von Groote, T.C.; Jayarajah, U.; Weerasekara, I.; Esfahani, M.A.; Civile, V.T.; Marusic, A.; Jeroncic, A.; et al. Novel Coronavirus Infection (COVID-19) in Humans: A Scoping Review and Meta-Analysis. J. Clin. Med. 2020, 9, 941. [Google Scholar] [CrossRef] [Green Version]
- Lepock, J.R. Measurement of protein stability and protein denaturation in cells using differential scanning calorimetry. Methods 2005, 35, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Todinova, S.; Krumova, S.; Gartcheva, L.; Robeerst, C.; Taneva, S.G. Microcalorimetry of blood serum proteome: A modified interaction network in the multiple myeloma case. Anal. Chem. 2011, 83, 7992–7998. [Google Scholar] [CrossRef] [PubMed]
- DeGregorio-Rocasolano, N.; Martí-Sistac, O.; Ponce, J.; Castelló-Ruiz, M.; Millán, M.; Guirao, V.; García-Yébenes, I.; Salom, J.B.; Ramos-Cabrer, P.; Alborch, E.; et al. Iron-loaded transferrin (Tf) is detrimental whereas iron-free Tf confers protection against brain ischemia by modifying blood Tf saturation and subsequent neuronal damage. Redox Biol. 2018, 15, 143–158. [Google Scholar] [CrossRef]
- Misiti, F. SARS-CoV-2 infection and red blood cells: Implications for long term symptoms during exercise. Sports Med. Health Sci. 2021, 269, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.; Stefanoni, D.; Dzieciatkowska, M.; Issaian, A.; Nemkov, T.; Hill, R.C.; Francis, R.O.; Hudson, K.E.; Buehler, P.W.; Zimring, J.C.; et al. Evidence of Structural Protein Damage and Membrane Lipid Remodeling in Red Blood Cells from COVID-19 Patients. J. Proteome Res. 2020, 19, 4455–4469. [Google Scholar] [CrossRef]
- Liu, W.; Li, H. COVID-19: Attacks the 1-Beta Chain of Hemoglobin and Captures the Porphyrin to Inhibit Heme Metabolism. ChemRxiv 2020. This content is a preprint and has not been peer-r. [Google Scholar]
- Campione, E.; Cosio, T.; Rosa, L.; Lanna, C.; Di Girolamo, S.; Gaziano, R.; Valenti, P.; Bianchi, L. Lactoferrin as protective natural barrier of respiratory and intestinal mucosa against coronavirus infection and inflammation. Int. J. Mol. Sci. 2020, 21, 4903. [Google Scholar] [CrossRef]
- Coates, T.D. Physiology and pathophysiology of iron in hemoglobin-associated diseases. Free Radic. Biol. Med. 2014, 72, 23–40. [Google Scholar] [CrossRef] [Green Version]
- Mainous, A.G.; Gill, J.M.; Carek, P.J. Elevated serum transferrin saturation and mortality. Ann. Fam. Med. 2004, 2, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.D.; Sumeh, A.S.; Sheraz, M.; Kavitha, M.S.; Venmathi Maran, B.A.; Rodrigues, K.F. A mini-review on the impact of COVID 19 on vital organs. Biomed. Pharmacother. 2021, 143, 112158. [Google Scholar] [CrossRef]
- John, A.E.; Joseph, C.; Jenkins, G.; Tatler, A.L. COVID-19 and pulmonary fibrosis: A potential role for lung epithelial cells and fibroblasts. Immunol. Rev. 2021, 302, 228–240. [Google Scholar] [CrossRef]
- Bonaccorsi di Patti, M.C.; Cutone, A.; Polticelli, F.; Rosa, L.; Lepanto, M.S.; Valenti, P.; Musci, G. The ferroportin-ceruloplasmin system and the mammalian iron homeostasis machine: Regulatory pathways and the role of lactoferrin. BioMetals 2018, 31, 399–414. [Google Scholar] [CrossRef]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Lactoferrin Against SARS-CoV-2: In Vitro and In Silico Evidences. Front. Pharmacol. 2021, 12, 666600. [Google Scholar] [CrossRef] [PubMed]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Lactoferrin as antiviral treatment in COVID-19 management: Preliminary evidence. Int. J. Environ. Res. Public Health 2021, 18, 10985. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.; Lepanto, M.S.; Cutone, A.; Siciliano, R.A.; Paesano, R.; Costi, R.; Musci, G.; Valenti, P. Influence of oral administration mode on the efficacy of commercial bovine Lactoferrin against iron and inflammatory homeostasis disorders. BioMetals 2020, 33, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Özdemir, Z.C.; Düzenli Kar, Y.; Gündüz, E.; Turhan, A.B.; Bör, Ö. Evaluation of hypercoagulability with rotational thromboelastometry in children with iron deficiency anemia. Hematology 2018, 23, 664–668. [Google Scholar] [PubMed] [Green Version]
- Michnik, A.; Drzazga, Z. Thermal denaturation of mixtures of human serum proteins: DSC study. J. Therm. Anal. Calorim. 2010, 101, 513–518. [Google Scholar] [CrossRef]
- Todinova, S.; Krumova, S.; Kurtev, P.; Dimitrov, V.; Djongov, L.; Dudunkov, Z.; Taneva, S.G. Calorimetry-based profiling of blood plasma from colorectal cancer patients. Biochim. Biophys. Acta 2012, 1820, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Suwalsky, M.; Fierro, P.; Villena, F.; Gallardo, M.J.; Jemiola-Rzeminska, M.; Strzalka, K.; Gul-Hinc, S.; Ronowska, A.; Zysk, M.; Szutowicz, A. Effects of sodium metavanadate on in vitro neuroblastoma and red blood cells. Arch. Biochem. Biophys. 2013, 535, 248–256. [Google Scholar]
- Kȩdra-Królik, K.; Chmielewska, I.; Michnik, A.; Zarzycki, P. Blood Serum Calorimetry Indicates the Chemotherapeutic Efficacy in Lung Cancer Treatment. Sci. Rep. 2017, 7, 16796. [Google Scholar] [CrossRef] [Green Version]
- Tsvetkov, P.; Devred, F.; Tsvetkov, P.O.; Ois Devred, F. Plasmatic Signature of Disease by Differential Scanning Calorimetry (DSC). In Microcalorimetry of Biological Molecules: Methods and Protocols; Eric Ennifar, Ed.; Springer: New York, NY, USA, 2019; pp. 45–57. ISBN 978-1-4939-9179-2. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thermal Parameters of Peaks 1 and 2 | ||||
|---|---|---|---|---|
| 25 h Incubation | 50 h Incubation | |||
| Untreated | SARS-CoV-2 | Untreated | SARS-CoV-2 | |
| Tm of Peak 1 (°C) | 69.58 ± 0.11 | 69.14 ± 0.14 | 69.14 ± 0.10 | 69.16 ± 0.09 |
| Tm of Peak 2 (°C) | 80.69 ± 1.36 * | 75.53 ± 0.24 * | 83.89 ± 0.58 | 82.12 ± 1.23 |
| Peak 2:1 ratio 1 | 0.63 | 0.54 | 1.25 | 0.74 |
| Calculated ΔH (J/g) | 3.51 ± 0.13 ** | 2.85 ± 0.05 ** | 4.12 ± 0.19 ** | 3.44 ± 0.10 ** |
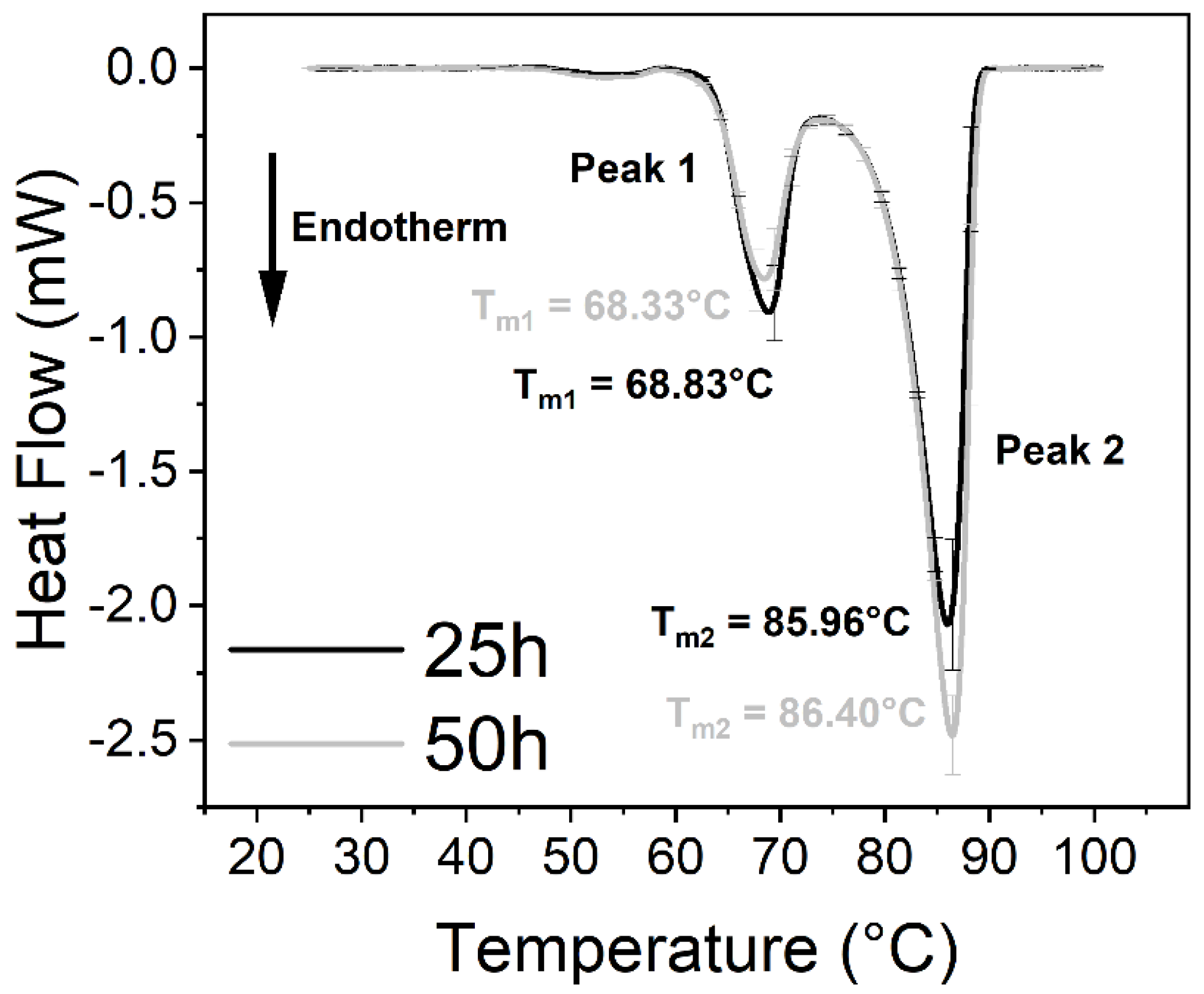
| Thermal Parameters of Peaks 1 and 2 of DMEM Control | ||
|---|---|---|
| 25 h Incubation | 50 h Incubation | |
| Tm of Peak 1 (°C) | 68.83 ± 0.17 | 68.33 ± 0.28 |
| Tm of Peak 2 (°C) | 85.96 ± 0.24 | 86.40 ± 0.18 |
| Peak 2:1 ratio 1 | 3.30 | 4.25 |
| Calculated ΔH (J/g) | 4.31 ± 0.06 | 4.64. ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Telek, E.; Ujfalusi, Z.; Kemenesi, G.; Zana, B.; Jakab, F.; Hild, G.; Lukács, A.; Hild, G. A Possible Way to Relate the Effects of SARS-CoV-2-Induced Changes in Transferrin to Severe COVID-19-Associated Diseases. Int. J. Mol. Sci. 2022, 23, 6189. https://doi.org/10.3390/ijms23116189
Telek E, Ujfalusi Z, Kemenesi G, Zana B, Jakab F, Hild G, Lukács A, Hild G. A Possible Way to Relate the Effects of SARS-CoV-2-Induced Changes in Transferrin to Severe COVID-19-Associated Diseases. International Journal of Molecular Sciences. 2022; 23(11):6189. https://doi.org/10.3390/ijms23116189
Chicago/Turabian StyleTelek, Elek, Zoltán Ujfalusi, Gábor Kemenesi, Brigitta Zana, Ferenc Jakab, Gabriella Hild, András Lukács, and Gábor Hild. 2022. "A Possible Way to Relate the Effects of SARS-CoV-2-Induced Changes in Transferrin to Severe COVID-19-Associated Diseases" International Journal of Molecular Sciences 23, no. 11: 6189. https://doi.org/10.3390/ijms23116189
APA StyleTelek, E., Ujfalusi, Z., Kemenesi, G., Zana, B., Jakab, F., Hild, G., Lukács, A., & Hild, G. (2022). A Possible Way to Relate the Effects of SARS-CoV-2-Induced Changes in Transferrin to Severe COVID-19-Associated Diseases. International Journal of Molecular Sciences, 23(11), 6189. https://doi.org/10.3390/ijms23116189