
Unraveling Nitrogen Fixing Potential of Endophytic Diazotrophs of Different Saccharum Species for Sustainable Sugarcane Growth

 ,
,  ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
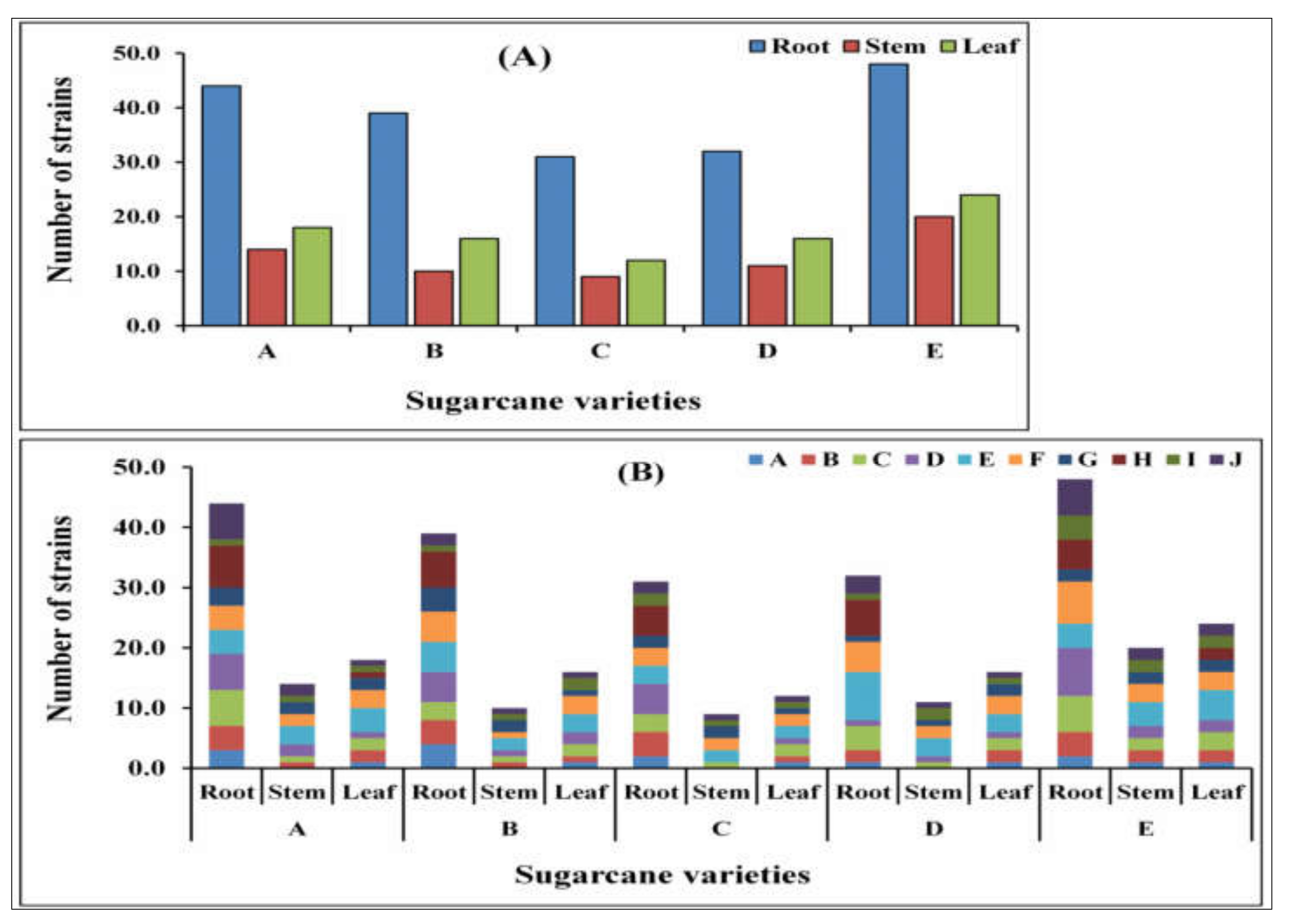
2.1. Bacterial Diversity Using the Culture-Dependent Method
2.2. Metagenomic Sequence Analysis of the nifH Gene
2.3. Richness and Evenness of Species
2.4. Operational Taxonomic Units (OTU) Analysis
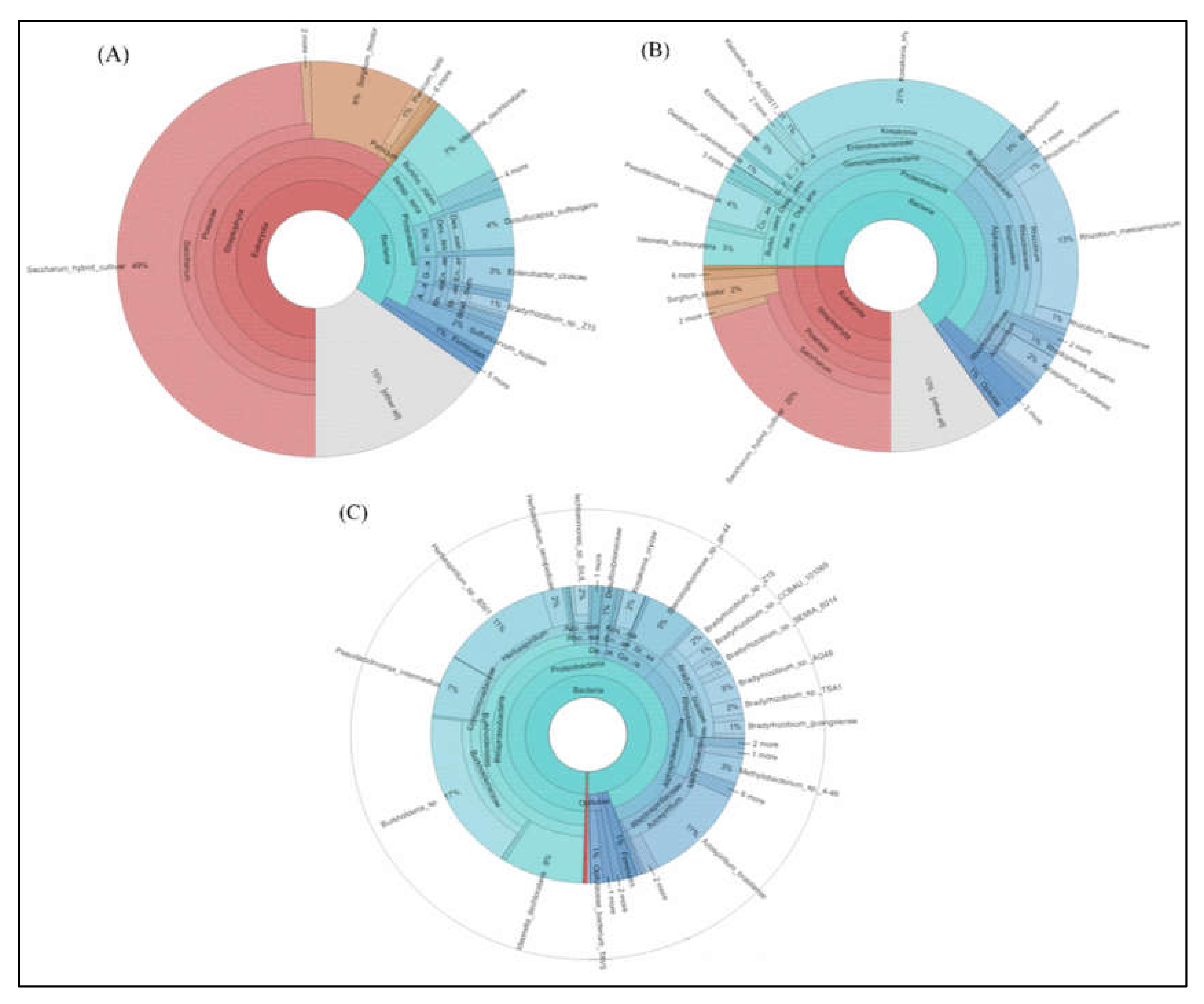
2.5. Metagenome Analysis Revealed Insight into the Major Microbial Taxonomic Classification from Sugarcane
2.6. Star Map Analysis of 10 Abundance Genus
2.7. Diversity Analysis of nifH Gene Sequences in Five Sugarcane Species
2.7.1. α-Diversity of Bacterial Community
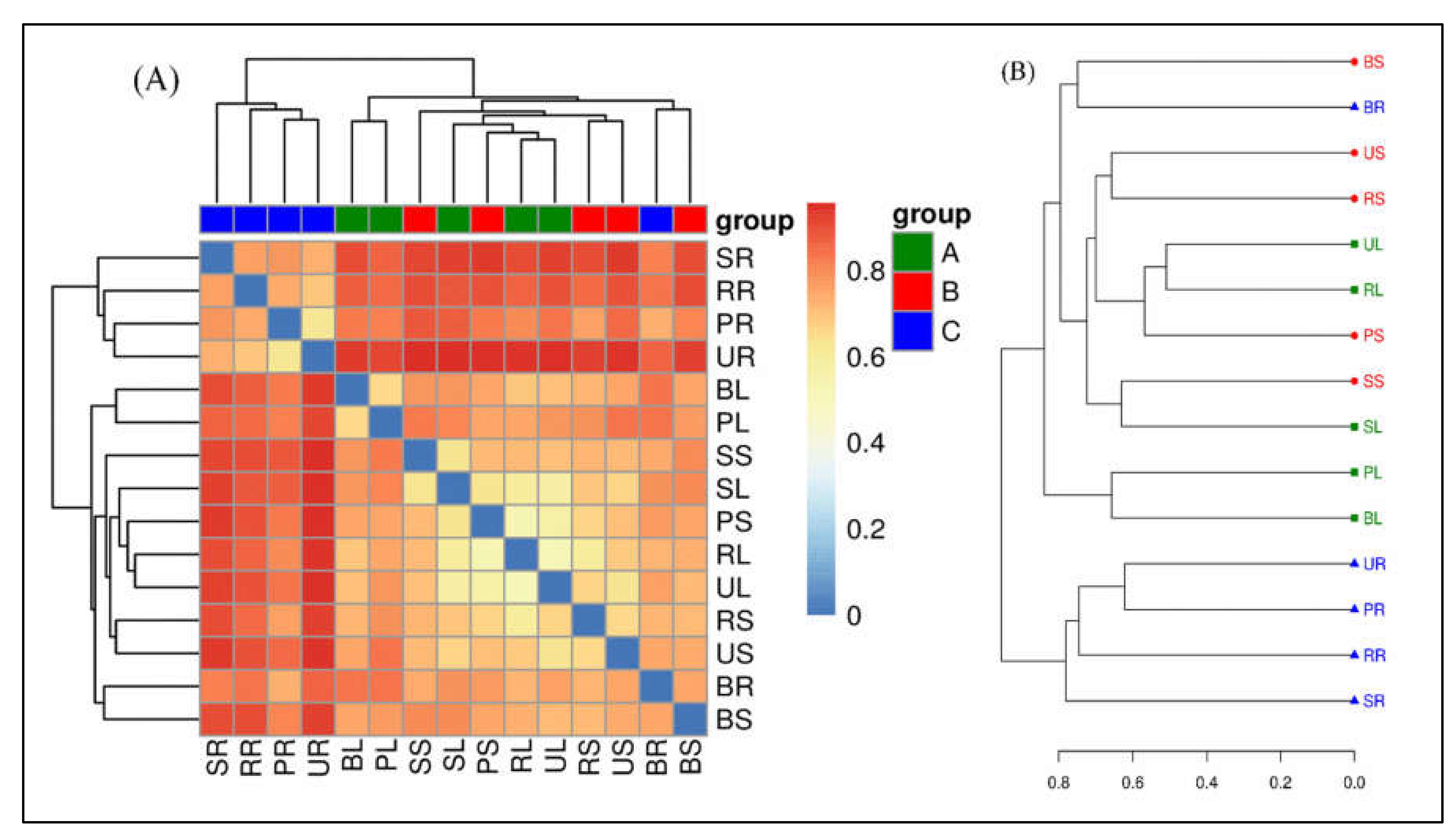
2.7.2. β-Diversity of Bacterial Community
2.8. Community Composition Histogram and Phylogeny Tree of Diazotrophs
2.9. Analysis of Species Differences in Each Classification Level
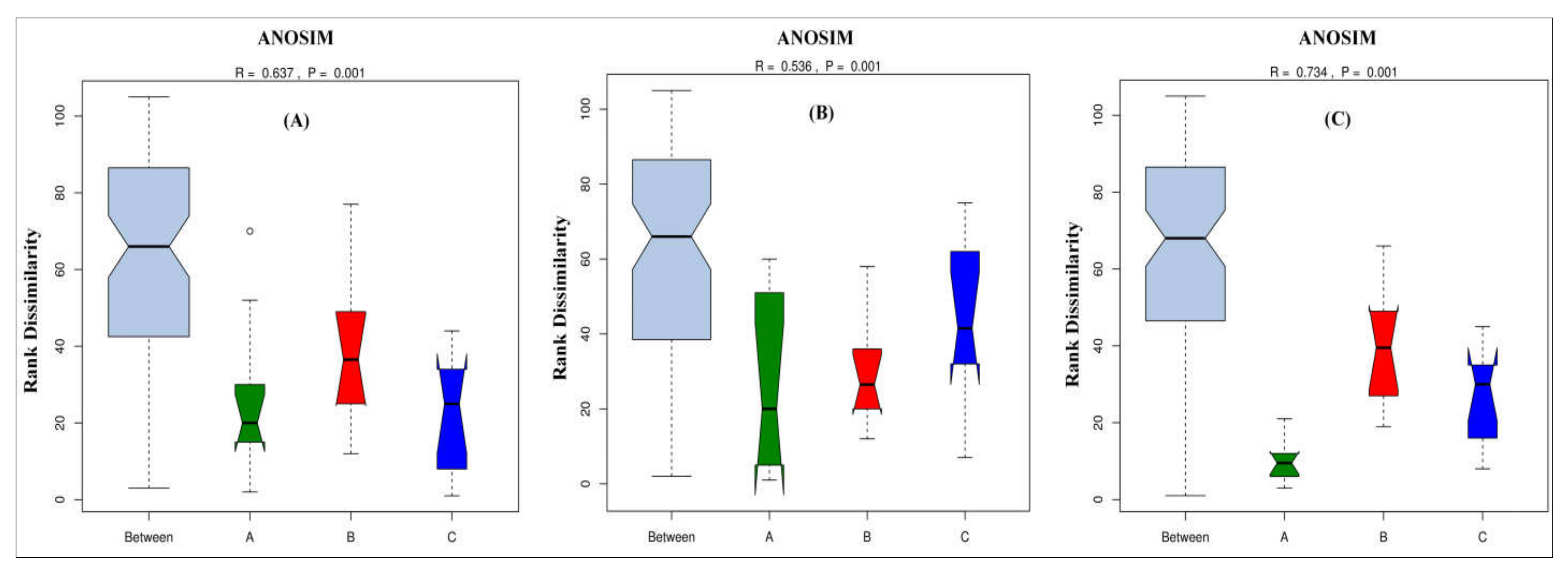
2.10. Analysis of Similarity (ANOSIM) Test
2.11. Linear Discriminant Analysis Effect Size (LEfSe)
3. Discussion
4. Material and Methods
4.1. Analysis of Endophytic Diazotrophic Bacterial Diversity through Culture-Dependent Techniques
4.1.1. Study Site and Sample Collection
4.1.2. Isolation of Endophytic Diazotrophs from Sugarcane Plants
4.1.3. Estimation for Nitrogenase Activity by Acetylene Reduction Assay
4.1.4. DNA Extraction, 16S and nifH Gene Amplification of Endophytic Microbes
4.2. Analysis of Endophytic Diazotrophic Bacterial Communities by Culture-Independent Technique
4.2.1. DNA Extraction
4.2.2. Construction of Sequencing Libraries and PCR Amplification
4.2.3. Diazotrophic Microbial Community Profile, Data Processing, and Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goldstein, D.; Mintz, S. The Oxford Companion to Sugar and Sweets; Oxford University Press: Oxford, UK, 2015. [Google Scholar] [CrossRef]
- Robinson, N.; Brackin, R.; Vinall, K.; Soper, F.; Holst, J.; Gamage, H. Nitrate paradigm does not hold up for sugarcane. PLoS ONE 2011, 6, e19045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Liu, X.; Zhang, Y.; Shen, J.; Han, W.; Zhang, W. Significant acidification in major Chinese croplands. Science 2010, 327, 1008–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zhang, F. Nitrogen fertilizer induced greenhouse gas emissions in China. Curr. Opin. Environ. Sustain. 2011, 3, 407–413. [Google Scholar] [CrossRef]
- Chen, X.; Cui, Z.; Fan, M.; Vitousek, P.; Zhao, M.; Ma, W.; Wang, Z.; Zhang, W.; Yan, X.; Yang, J.; et al. Producing more grain with lower environmental costs. Nature 2014, 514, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, S.; Wang, Y.; Zhang, Y.; Hu, C.; Liu, B. Long-Term Nitrogen Fertilization Elevates the Activity and Abundance of Nitrifying and Denitrifying Microbial Communities in an Upland Soil: Implications for Nitrogen Loss from Intensive Agricultural Systems. Front. Microbiol. 2018, 9, 2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menéndez, E.; Paço, A. Is the application of plant probiotic bacterial consortia always beneficial for plants? Exploring synergies between rhizobial and non-rhizobial bacteria and their effects on agro-economically valuable crops. Life 2020, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Rubio, L.M.; Ludden., P.W. The gene products of the nif regulon. In Nitrogen Fixation at the Millennium, 1st ed.; Leigh, G.J., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2002; pp. 101–136. [Google Scholar]
- Gaby, J.C.; Buckley, D.H. A comprehensive aligned nifH gene database: A multipurpose tool for studies of nitrogen-fixing bacteria. Database 2014, 2014, bau001. [Google Scholar] [CrossRef] [Green Version]
- Roesch, L.F.W.; Camargo, F.A.O.; Bento, F.M.; Triplett, E.W. Biodiversity of diazotrophic bacteria within the soil, root and stem of field-grown maize. Plant Soil 2008, 302, 91–104. [Google Scholar] [CrossRef]
- Li, H.B.; Singh, R.K.; Singh, P.; Song, Q.Q.; Xing, Y.X.; Yang, L.T.; Li, Y.R. Genetic Diversity of Nitrogen-Fixing and Plant Growth Promoting Pseudomonas Species Isolated from Sugarcane Rhizosphere. Front. Microbiol. 2017, 8, 1268. [Google Scholar] [CrossRef]
- Singh, R.K.; Singh, P.; Li, H.B.; Song, Q.Q.; Guo, D.J.; Solanki, M.K.; Verma, K.K.; Malviya, M.K.; Song, X.P.; Lakshmanan, P.; et al. Diversity of nitrogen-fixing rhizobacteria associated with sugarcane: A comprehensive study of plant-microbe interactions for growth enhancement in Saccharum spp. BMC Plant Biol. 2020, 20, 220. [Google Scholar] [CrossRef]
- Singh, P.; Singh, R.K.; Li, H.B.; Guo, D.J.; Sharma, A.; Lakshmanan, P.; Malviya, M.K.; Song, X.P.; Solanki, M.K.; Verma, K.K.; et al. Diazotrophic Bacteria Pantoea dispersa and Enterobacter asburiae Promote Sugarcane Growth by Inducing Nitrogen Uptake and Defense-Related Gene Expression. Front. Microbiol. 2021, 11, 600417. [Google Scholar] [CrossRef]
- Singh, R.K.; Singh, P.; Guo, D.J.; Sharma, A.; Li, D.P.; Li, X.; Verma, K.K.; Malviya, M.K.; Song, X.P.; Lakshmanan, P.; et al. Root-Derived Endophytic Diazotrophic Bacteria Pantoea cypripedii AF1 and Kosakonia arachidis EF1 Promote Nitrogen Assimilation and Growth in Sugarcane. Front. Microbiol. 2021, 12, 774707. [Google Scholar] [CrossRef]
- Magnani, G.S.; Didonet, C.M.; Cruz, L.M.; Picheth, C.F.; Pedrosa, F.O.; Souza, E.M. Diversity of endophytic bacteria in Brazilian sugarcane. Genet. Mol. Res. 2010, 9, 250–258. [Google Scholar] [CrossRef]
- Olivares, F.L.; Baldani, V.L.D.; Reis, V.M.; Baldani, J.I.; Döbereiner, J. Occurrence of the endophytic diazotrophs Herbaspirillum spp. in roots, stems, and leaves, predominantly of Gramineae. Biol. Fertil. Soils 1996, 21, 197–200. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Martínez-Romero, E. Bacterial endophytes and their interactions with hosts. Mol. Plant Microbe Interact. 2006, 19, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Reinhold-Hurek, B.; Hurek, T. Life in grasses: Diazotrophic endophytes. Trends Microbiol. 1998, 6, 139–144. [Google Scholar] [CrossRef]
- Doty, S.L.; Sher, A.W.; Fleck, N.D.; Khorasani, M.; Bumgarner, R.E.; Khan, Z.; Ko, A.W.K.; Kim, S.-H.; DeLuca, T.H. Variable Nitrogen Fixation in Wild Populus. PLoS ONE 2016, 11, e0155979. [Google Scholar] [CrossRef] [Green Version]
- Schütz, L.; Gattinger, A.; Meier, M.; Müller, A.; Boller, T.; Mäder, P.; Mathimaran, N. Improving crop yield and nutrient use efficiency via biofertilization—A global meta-analysis. Front. Plant Sci. 2018, 8, 2204. [Google Scholar] [CrossRef] [Green Version]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [Green Version]
- Meena, M.; Swapnil, P.; Divyanshu, K.; Kumar, S.; Harish; Tripathi, Y.N.; Zehra, A.; Marwal, A.; Upadhyay, R.S. PGPR-mediated induction of systemic resistance and physiochemical alterations in plants against the pathogens: Current perspectives. J. Basic Microbiol. 2020, 60, 828–861. [Google Scholar] [CrossRef]
- Raklami, A.; Bechtaoui, N.; Tahiri, A.; Anli, M.; Meddich, A.; Oufdou, K. Use of rhizobacteria and mycorrhizae consortium in the open field as a strategy for improving crop nutrition, productivity and soil fertility. Front. Microbiol. 2019, 10, 1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Bai, Z.; Bao, L.; Xue, L.; Zhang, S.; Wei, Y.; Zhang, Z.; Zhuang, G.; Zhuang, X. Bacillus subtilis biofertilizer mitigating agricultural ammonia emission and shifting soil nitrogen cycling microbiomes. Environ. Int. 2020, 144, 105989. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahanty, T.; Bhattacharjee, S.; Goswami, M.; Bhattacharyya, P.; Das, B.; Ghosh, A.; Tribedi, P. Biofertilizers: A potential approach for sustainable agriculture development. Environ. Sci. Pollut. Res. 2017, 24, 3315–3335. [Google Scholar] [CrossRef]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Frank, I.E.; Turk-Kubo, K.A.; Zehr, J.P. Rapid annotation of nifH gene sequences using classification and regression trees facilitates environmental functional gene analysis. Environ. Microbiol. Rep. 2016, 8, 905–916. [Google Scholar] [CrossRef]
- Dong, M.; Yang, Z.; Cheng, G.; Peng, L.; Xu, Q.; Xu, J. Diversity of the bacterial microbiome in the roots of four saccharum species: S. spontaneum, S. robustum, S. barberi and S. officinarum. Front. Microbiol. 2018, 9, 267. [Google Scholar] [CrossRef]
- Gaby, J.C.; Buckley, D.H. The use of degenerate primers in qPCR analysis of functional genes can cause dramatic quantification bias as revealed by investigation of nifH primer performance. Microb. Ecol. 2017, 74, 701–708. [Google Scholar] [CrossRef]
- Babalola, O.O.; Sanni, A.I.; Odhiambo, G.D.; Torto, B. Plant growth-promoting rhizobacteria do not pose any deleterious effect on cowpea and detectable amounts of ethylene are produced. World J. Microbiol. Biotechnol. 2007, 23, 747–752. [Google Scholar] [CrossRef]
- Adedeji, A.A.; Haggblom, M.M.; Babalola, O.O. Sustainable agriculture in Africa: Plant growth-promoting rhizobacteria (PGPR) to the rescue. Sci. Afr. 2020, 9, e00492. [Google Scholar] [CrossRef]
- Compant, S.; Reiter, B.; Sessitsch, A.; Nowak, J.; Clément, C.; Barka, E.A. Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef] [Green Version]
- Kandel, S.; Joubert, P.; Doty, S. Bacterial endophyte colonization and distribution within plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Chen, L.; Xin, H.L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.P. A friendly relationship between endophytic fungi and medicinal plants: A systematic review. Front. Microbiol. 2016, 7, 906. [Google Scholar] [CrossRef] [Green Version]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–687. [Google Scholar] [CrossRef] [Green Version]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Mei, C.; Flinn, B.S. The use of beneficial microbial endophytes for plant biomass and stress tolerance improvement. Recent Pat. Biotechnol. 2010, 4, 81–95. [Google Scholar] [CrossRef]
- Sturz, A.; Nowak, J. Endophytic communities of rhizobacteria and the strategies required to create yield enhancing associations with crops. Appl. Soil Ecol. 2000, 15, 183–190. [Google Scholar] [CrossRef]
- Di, Y.; Kui, L.; Singh, P.; Liu, L.F.; Xie, L.Y.; He, L.L.; Li, F.S. Identification and Characterization of Bacillus subtilis B9: A Diazotrophic Plant Growth-Promoting Endophytic Bacterium Isolated from Sugarcane Root. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Prayogo, F.A.; Budiharjo, A.; Kusumaningrum, H.P.; Wijanarka, W.; Suprihadi, A.; Nurhayati, N. Metagenomic applications in exploration and development of novel enzymes from nature: A review. J. Genet. Eng. Biotechnol. 2020, 18, 39. [Google Scholar] [CrossRef]
- Lehnert, N.; Dong, H.T.; Harland, J.B.; Hunt, A.P.; White, C.J. Reversing nitrogen fixation. Nat. Rev. Chem. 2018, 2, 278–289. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Ormeño-Orrillo, E.; López-López, A.; Rogel, M.A.; Reyes-Hernández, B.J.; Martínez-Romero, J.C.; Reddy, P.M.; Martínez-Romero, E. Nitrogen fixation in cereals. Front. Microbiol. 2018, 9, 1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, R.W.F.; Holsten, R.D.; Jackson, E.K.; Burns, R.C. The acetylene ethylene assay for N fixation, laboratory and field evaluation. Plant Physiol. 1968, 43, 1185–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kifle, M.H.; Laing, M.D. Isolation and screening of bacteria for their diazotrophic potential and their influence on growth promotion of maize seedlings in greenhouses. Front. Plant Sci. 2015, 6, 1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brusamarello-Santos, L.C.; Gilard, F.; Brulé, L.; Quilleré, I.; Gourion, B.; Ratet, P.; Maltempi de Souza, E.; Lea, P.J.; Hirel, B. Metabolic profiling of two maize (Zea mays L.) inbred lines inoculated with the nitrogen fixing plant-interacting bacteria Herbaspirillum seropedicae and Azospirillum brasilense. PLoS ONE 2017, 12, e0174576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Y.X.; Yang, L.T.; Huang, S.L.; Li, Y.R. Identification of a new nitrogen fixing endo-bacterium strain isolated from sugarcane stalk. Sugar Tech. 2006, 8, 49–53. [Google Scholar] [CrossRef]
- Mehnaz, S.; Baig, D.N.; Lazarovits, G. Genetic and phenotypic diversity of plant growth promoting rhizobacteria isolated from sugarcane plants growing in Pakistan. J. Microbiol. Biotechnol. 2010, 20, 1614–1623. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Guo, W.; Xing, Y.; Zhang, X.; Li, Z.; Hu, C.; Li, S.; Li, Y.R.; An, Q. The actinobacterium Microbacterium sp. 16SH accepts pBBR1-based pPROBE vectors, forms biofilms, invades roots, and fixes N2 associate d with micropropagated sugarcane plants. Appl. Microbiol. Biotechnol. 2012, 93, 1185–1195. [Google Scholar] [CrossRef]
- Guo, D.J.; Singh, R.K.; Singh, P.; Li, D.P.; Sharma, A.; Xing, Y.X.; Song, X.P.; Yang, L.T.; Li, Y.R. Complete Genome Sequence of Enterobacter roggenkampii ED5, a Nitrogen Fixing Plant Growth Promoting Endophytic Bacterium with Biocontrol and Stress Tolerance Properties, Isolated from Sugarcane Root. Front. Microbiol. 2020, 11, 580081. [Google Scholar] [CrossRef]
- Bowen, G.D.; Rovira, A.D. The rhizosphere and its management to improve plant growth. Adv. Agron. 1999, 66, 1–102. [Google Scholar]
- Donn, S.; Kirkegaard, J.A.; Perera, G.; Richardson, A.E.; Watt, M. Evolution of bacterial communities in the wheat crop rhizosphere. Environ. Microbiol. 2015, 17, 610–621. [Google Scholar] [CrossRef]
- Bouffaud, M.; Poirier, M.; Muller, D.; Moënne-Loccoz, Y. Root microbiome relates to plant host evolution in maize and other Poaceae. Environ. Microbiol. 2014, 16, 2804–2814. [Google Scholar] [CrossRef]
- Aasfar, A.; Bargaz, A.; Yaakoubi, K.; Hilali, A.; Bennis, I.; Zeroual, Y.; Meftah Kadmiri, I. Nitrogen Fixing Azotobacter Species as Potential Soil Biological Enhancers for Crop Nutrition and Yield Stability. Front. Microbiol. 2021, 12, 628379. [Google Scholar] [CrossRef]
- Fukami, J.; Cerezini, P.; Hungria, M. Azospirillum: Benefits that go far beyond biological nitrogen fixation. AMB Expr. 2018, 8, 73. [Google Scholar] [CrossRef]
- Cortés-Patiño, S.; Vargas, C.; Álvarez-Flórez, F.; Bonilla, R.; Estrada-Bonilla, G. Potential of Herbaspirillum and Azospirillum Consortium to Promote Growth of Perennial Ryegrass under Water Deficit. Microorganisms 2021, 9, 91. [Google Scholar] [CrossRef]
- Carvalho, T.L.; Pires, E.; Saraiva, R.; Vargas, L.; Bomfim, A.C.J.; Ballesteros, H.; Baldani, J.I.; Ferreira, P.C.; Hemerly, A.S. Nitrogen fixation in grasses—Gluconacetobacter activates genes in sugarcane. BMC Proc. 2014, 8, O20. [Google Scholar] [CrossRef] [Green Version]
- Paungfoo-Lonhienne, C.; Lonhienne, T.G.; Yeoh, Y.K.; Donose, B.C.; Webb, R.I.; Parsons, J.; Liao, W.; Sagulenko, E.; Lakshmanan, P.; Hugenholtz, P.; et al. Crosstalk between sugarcane and a plant-growth promoting Burkholderia species. Sci. Rep. 2016, 6, 37389. [Google Scholar] [CrossRef]
- Meng, H.; Zhou, Z.; Wu, R.; Wang, Y.; Gu, J.D. Diazotrophic microbial community and abundance in acidic subtropical natural and revegetated forest soils revealed by high-throughput sequencing of nifH gene. Appl. Microbiol. Biotech. 2019, 103, 995–1005. [Google Scholar] [CrossRef]
- Luo, J.; Lin, Z.L.; Li, S.Y.; Que, Y.X.; Zhang, C.F.; Yang, Z.Q.; Yao, K.C.; Feng, J.F.; Chen, J.F.; Zhang, H. Effects of different soil improvement measures on soil physicochemical properties and microbial community structures in mechanically compacted acidified sugarcane field. Acta Agron. 2020, 46, 596–613. [Google Scholar]
- Gu, Y.; Wang, J.; Cai, W.; Li, G.; Mei, Y.; Yang, S. Different Amounts of Nitrogen Fertilizer Applications Alter the Bacterial Diversity and Community Structure in the Rhizosphere Soil of Sugarcane. Front. Microbiol. 2021, 12, 721441. [Google Scholar] [CrossRef]
- Singh, P.; Singh, R.K.; Song, Q.Q.; Li, H.B.; Yang, L.T.; Li, Y.R. Methods for estimation of nitrogen components in plants and microorganisms. Methods Mol. Biol. 2020, 2057, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poly, F.; Monrozier, L.J.; Bally, R. Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Res. Microbiol. 2001, 152, 95–103. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 10 January 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and pre-processing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Huse, S.M.; Welch, D.M.; Morrison, H.G.; Sogin, M.L. Ironing out the wrinkles in the rare biosphere through improved OTU clustering. Environ. Microbiol. 2010, 12, 1889–1898. [Google Scholar] [CrossRef] [Green Version]
- Roeselers, G.; Mittge, E.K.; Stephens, W.Z.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a core gut microbiota in the zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Chai, B.; Farris, R.J.; Wang, Q.; Kulam, S.A.; McGarrell, D.M.; Garrity, G.M.; Tiedje, J.M. The Ribosomal Database Project (RDP-II): Sequences and tools for high throughput rRNA analysis. Nucleic Acids Res. 2005, 33, D294–D296. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved green genes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949; pp. 1–117. [Google Scholar]
- Rodrigues, V.D.; Torres, T.T.; Ottoboni, L.M. Bacterial diversity assessment in soil of an active Brazilian copper mine using high-throughput sequencing of 16S rDNA amplicons. Anton Leeuw. Int. J. Gen. 2014, 106, 879–890. [Google Scholar] [CrossRef]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shen, T.J. A new statistical approach for assessing compositional similarity based on incidence and abundance data. Ecol. Lett. 2005, 80, 148–159. [Google Scholar]
- Hughes, J.B.; Hellmann, J.J.; Ricketts, T.H.; Bohannan, J.M. Counting the uncountable: Statistical approaches to estimating microbial diversity. Appl. Environ. Microbiol. 2001, 67, 4399–4406. [Google Scholar] [CrossRef] [Green Version]
- Hamady, M.; Lozupone, C.; Knight, R. Fast UniFrac: Facilitating high-throughput phylogenetic analyses of microbial communities including analysis of pyrosequencing and PhyloChip data. ISME J. 2010, 4, 17–27. [Google Scholar] [CrossRef]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Li, S.; Yang, L.; Huang, P.; Li, W.; Wang, S.; Zhao, G.; Zhang, M.; Pang, X.; Yan, Z.; et al. Structural modulation of gut microbiota in life-long calorie-restricted mice. Nat. Commun. 2013, 4, 2163. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.Q.W.; Yan, W.M.; Shangguan, Z.P. Impact of long-term N additions upon coupling between soil microbial community structure and activity, and nutrient use efficiencies. Soil Biol. Biochem. 2015, 91, 151–159. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | nifH Gene Amplification | Accession Number | ARA (nmoL C2H4 mg Protein h−1) |
|---|---|---|---|
| Achromobacter sp. | + | - | 12.415 ± 0.225 r |
| Achromobacter xylosoxidans | + | - | 17.133 ± 0.310 l |
| Atlantibacter hermannii | + | - | 13.258 ± 0.240 q |
| Bacillus sp. | + | - | 9.274 ± 0.168 v |
| Bacillus amyloliquefaciens | + | - | 11.161 ± 0.202 t |
| Bacillus velezensis | + | - | 21.800 ± 0.395 g |
| Burkholderia sp. | + | - | 26.457 ± 0.479 b |
| Burkholderia cenocepacia | + | - | 15.778 ± 0.286 mn |
| Burkholderia gladioli | + | - | 18.498 ± 0.335 jk |
| Chryseobacterium sp. | + | - | 14.513 ± 0.263 o |
| Enterobacter sp. | + | MT649072 | 11.173 ± 0.191 u |
| Enterobacter asburiae | + | MT649071 | 08.234 ± 0.222 rs |
| Enterobacter cloacae | + | MT649070 | 14.724 ± 0.267 o |
| Enterobacter roggenkampii | + | MT649078 | 29.709 ± 0.538 a |
| Enterobacter tabaci | + | - | 13.309 ± 0.241 q |
| Herbaspirillum aquaticum | + | - | 24.781 ± 0.449 cd |
| Kosakonia oryzae | + | MZ497007 | 10.285 ± 0.422 f |
| Lelliottia nimipressuralis | + | - | 24.158 ± 0.438 e |
| Lysinibacillus macroides | + | - | 13.891 ± 0.252 p |
| Metakosakonia sp. | + | - | 6.163 ± 0.112 w |
| Microbacterium sp. | + | - | 24.570 ± 0.445 de |
| Pantoea sp. | + | - | 15.356 ± 0.278 n |
| Pantoea agglomerans | + | MZ502262 | 10.082 ± 0.183 u |
| Pantoea ananatis | + | MZ502260 | 18.645 ± 0.339 j |
| Pantoea dispersa | + | MZ502257 | 15.305 ± 0.422 f |
| Pseudomonas sp. | + | - | 17.243 ± 0.312 l |
| Pseudomonas aeruginosa | + | MW027642 | 12.626 ± 0.229 r |
| Pseudomonas chlororaphis | + | - | 18.156 ± 0.329 k |
| Pseudomonas plecoglossicida | + | - | 18.919 ± 0.343 ij |
| Pseudomonas putida | + | - | 25.202 ± 0.456 c |
| Pseudomonas koreensis | + | - | 15.567 ± 0.282 n |
| Pseudomonas taiwanensis | + | - | 12.215 ± 0.221 rs |
| Rhizobium sp. | + | - | 13.258 ± 0.240 q |
| Serratia sp. | + | - | 18.919 ± 0.343 ij |
| Serratia marcescens | + | - | 14.513 ± 0.263 o |
| Sphingomonas azotifigens | + | - | 17.614 ± 0.319 l |
| Sphingomonas echinoides | + | - | 19.401 ± 0.351 hi |
| Sphingomonas trueperi | + | - | 16.189 ± 0.293 m |
| Staphylococcus arlettae | + | - | 11.793 ± 0.214 s |
| Stenotrophomonas sp. | + | - | 13.258 ± 0.240 q |
| Stenotrophomonas maltophilia | + | - | 12.415 ± 0.225 r |
| Xanthomonas sacchari | + | - | 19.541 ± 0.354 h |
| Sugarcane Variety | Plant Parts | Number of Valid Sequences | Optimized Number of Sequences | Optimized Number of Sequence Bases | Optimized Sequence GC Content (%) | Optimized Average Sequence Length | Optimized Sequence Length Range |
|---|---|---|---|---|---|---|---|
| S. officinarum L. cv. Badila | BL | 32,624 | 25,686 | 7,891,580 | 50.374 | 307 | 200→541 |
| BS | 18,573 | 15,532 | 5,212,342 | 58.127 | 336 | 200→502 | |
| BR | 36,189 | 27,995 | 10,001,455 | 61.731 | 357 | 201→469 | |
| S. barberi Jesw. cv Pansahi | PL | 19,097 | 12,937 | 3,714,556 | 42.949 | 297 | 200→451 |
| PS | 21,840 | 15,093 | 4,309,289 | 44.523 | 296 | 201→541 | |
| PR | 37,161 | 21,601 | 7,752,193 | 61.927 | 359 | 202→405 | |
| S. robustum | RL | 27,071 | 11,358 | 3,669,350 | 56.315 | 323 | 200→470 |
| RS | 37,944 | 24,191 | 8,616,963 | 55.621 | 356 | 201→470 | |
| RR | 37,700 | 30,476 | 10,925,709 | 62.800 | 359 | 202→393 | |
| S. spontaneum | SL | 35,409 | 21,038 | 5,897,235 | 48.693 | 290 | 200→471 |
| SS | 37,892 | 19,058 | 5,851,645 | 56.441 | 307 | 202→476 | |
| SR | 36,135 | 31,283 | 11,253,996 | 62.336 | 360 | 261→470 | |
| S. sinense Roxb. cv Uba | UL | 31,562 | 16,004 | 4,724,294 | 48.035 | 295 | 200→478 |
| US | 36,943 | 25,611 | 8,610,553 | 57.739 | 336 | 200→478 | |
| UR | 36,005 | 21,903 | 7,870,912 | 63.059 | 359 | 226→369 |
| Sugarcane Variety | Sample Code | Reads | OTU | Ace | Chao | Coverage | Shannon | Simpson | Sobs | PD Whole Tree |
|---|---|---|---|---|---|---|---|---|---|---|
| S. officinarum L. cv. Badila | BL | 25,686 | 150 | 231.49 | 193.50 | 0.998832 | 3.78 | 0.0445 | 150.00 | 53.66 |
| BR | 27,995 | 170 | 186.34 | 198.11 | 0.999178 | 2.52 | 0.2148 | 170.00 | 45.02 | |
| BS | 15,532 | 136 | 160.16 | 155.12 | 0.998326 | 3.49 | 0.0503 | 136.00 | 55.06 | |
| S. barberi Jesw. cv Pansahi | PL | 12,937 | 106 | 125.54 | 121.11 | 0.998686 | 3.33 | 0.0808 | 106.00 | 33.68 |
| PR | 21,601 | 261 | 279.13 | 286.37 | 0.998657 | 4.08 | 0.0426 | 261.00 | 26.27 | |
| PS | 15,093 | 295 | 446.16 | 395.04 | 0.995097 | 4.52 | 0.0172 | 295.00 | 79.38 | |
| S. robustum | RL | 11,358 | 281 | 344.88 | 346.21 | 0.994541 | 3.79 | 0.0998 | 281.00 | 73.35 |
| RR | 30,476 | 152 | 160.49 | 174.00 | 0.999606 | 3.71 | 0.0587 | 152.00 | 19.59 | |
| RS | 24,191 | 184 | 222.06 | 234.17 | 0.998222 | 1.62 | 0.363 | 184.00 | 57.39 | |
| S. spontaneum | SL | 21,038 | 293 | 411.03 | 371.00 | 0.997528 | 4.34 | 0.0354 | 293.00 | 88.57 |
| SR | 31,283 | 93 | 96.90 | 95.50 | 0.999840 | 2.53 | 0.1437 | 93.00 | 11.37 | |
| SS | 19,058 | 206 | 216.61 | 219.13 | 0.999213 | 4.5 | 0.0191 | 206.00 | 88.53 | |
| S. sinense Roxb. cv Uba | UL | 16,004 | 394 | 419.22 | 429.65 | 0.997438 | 4.92 | 0.0228 | 394.00 | 94.76 |
| UR | 21,903 | 220 | 223.81 | 224.09 | 0.999543 | 4.07 | 0.0389 | 220.00 | 13.87 | |
| US | 25,611 | 256 | 277.85 | 283.00 | 0.998907 | 3.07 | 0.2503 | 256.00 | 87.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.K.; Singh, P.; Sharma, A.; Guo, D.-J.; Upadhyay, S.K.; Song, Q.-Q.; Verma, K.K.; Li, D.-P.; Malviya, M.K.; Song, X.-P.; et al. Unraveling Nitrogen Fixing Potential of Endophytic Diazotrophs of Different Saccharum Species for Sustainable Sugarcane Growth. Int. J. Mol. Sci. 2022, 23, 6242. https://doi.org/10.3390/ijms23116242
Singh RK, Singh P, Sharma A, Guo D-J, Upadhyay SK, Song Q-Q, Verma KK, Li D-P, Malviya MK, Song X-P, et al. Unraveling Nitrogen Fixing Potential of Endophytic Diazotrophs of Different Saccharum Species for Sustainable Sugarcane Growth. International Journal of Molecular Sciences. 2022; 23(11):6242. https://doi.org/10.3390/ijms23116242
Chicago/Turabian StyleSingh, Rajesh Kumar, Pratiksha Singh, Anjney Sharma, Dao-Jun Guo, Sudhir K. Upadhyay, Qi-Qi Song, Krishan K. Verma, Dong-Ping Li, Mukesh Kumar Malviya, Xiu-Peng Song, and et al. 2022. "Unraveling Nitrogen Fixing Potential of Endophytic Diazotrophs of Different Saccharum Species for Sustainable Sugarcane Growth" International Journal of Molecular Sciences 23, no. 11: 6242. https://doi.org/10.3390/ijms23116242
APA StyleSingh, R. K., Singh, P., Sharma, A., Guo, D.-J., Upadhyay, S. K., Song, Q.-Q., Verma, K. K., Li, D.-P., Malviya, M. K., Song, X.-P., Yang, L.-T., & Li, Y.-R. (2022). Unraveling Nitrogen Fixing Potential of Endophytic Diazotrophs of Different Saccharum Species for Sustainable Sugarcane Growth. International Journal of Molecular Sciences, 23(11), 6242. https://doi.org/10.3390/ijms23116242