
Effect of Anti-Osteoporotic Treatments on Circulating and Bone MicroRNA Patterns in Osteopenic ZDF Rats

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Study Design
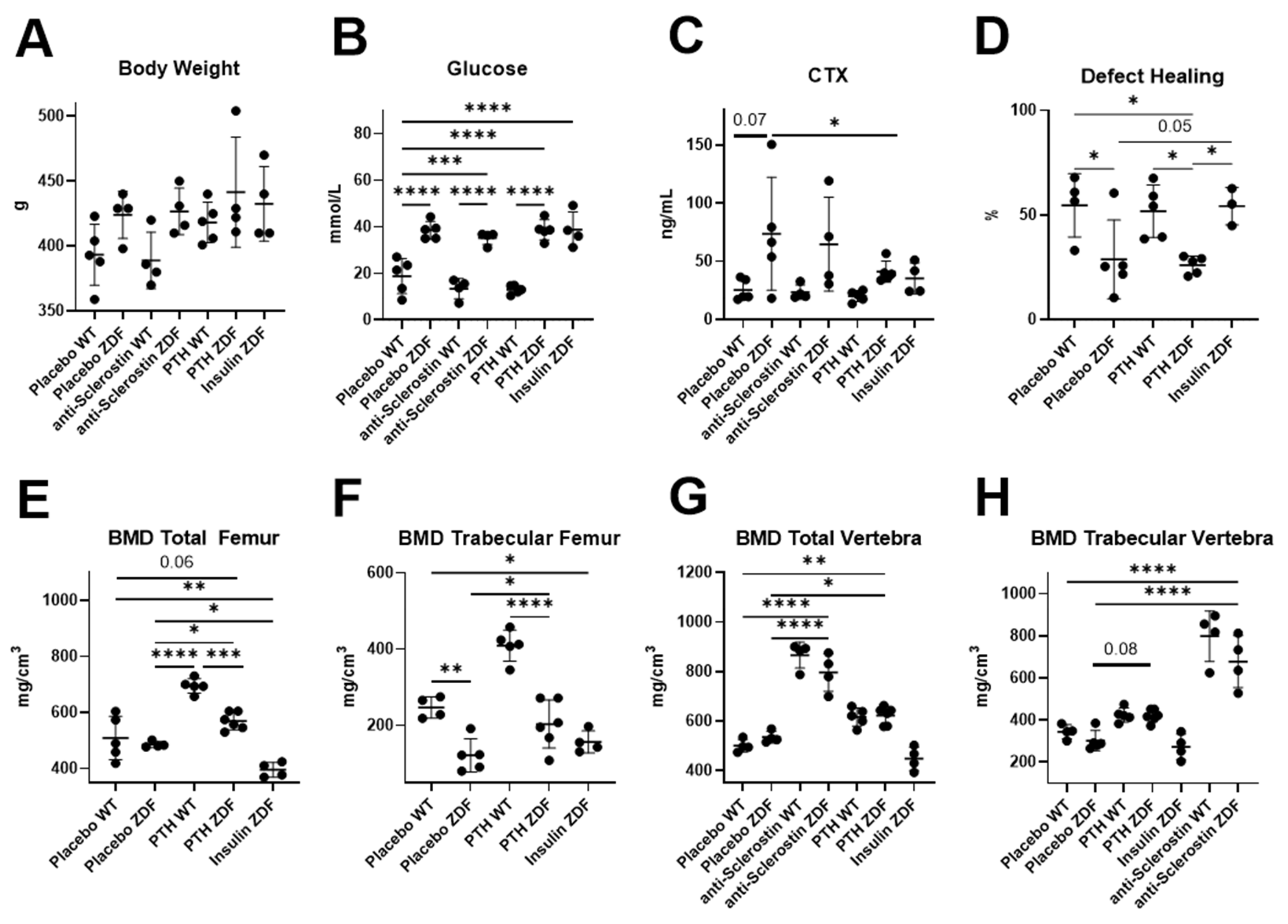
2.2. Animal Phenotype Analysis
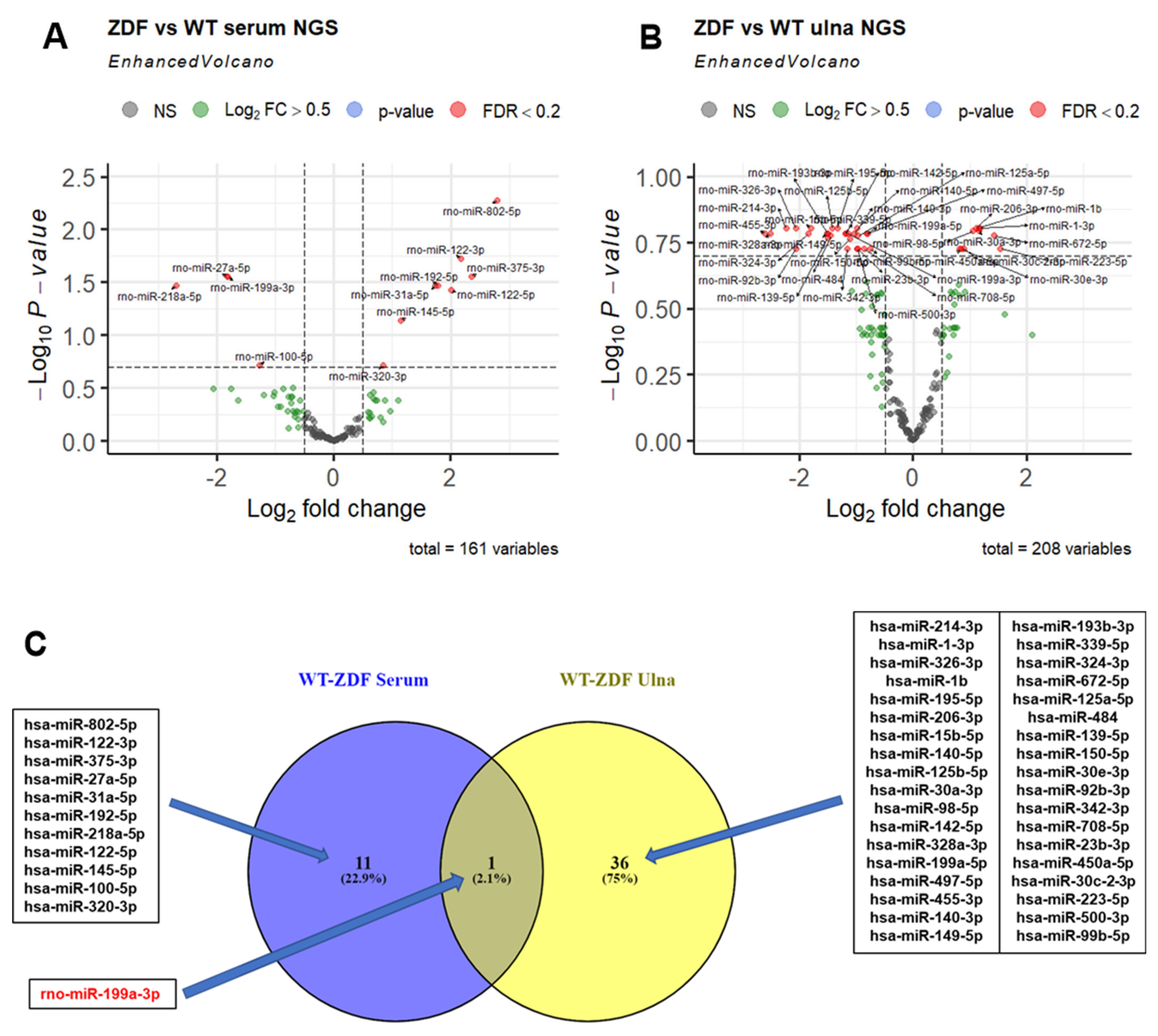
2.3. MiRNA Dysregulation in Serum and Bone Tissue of ZDF Rats
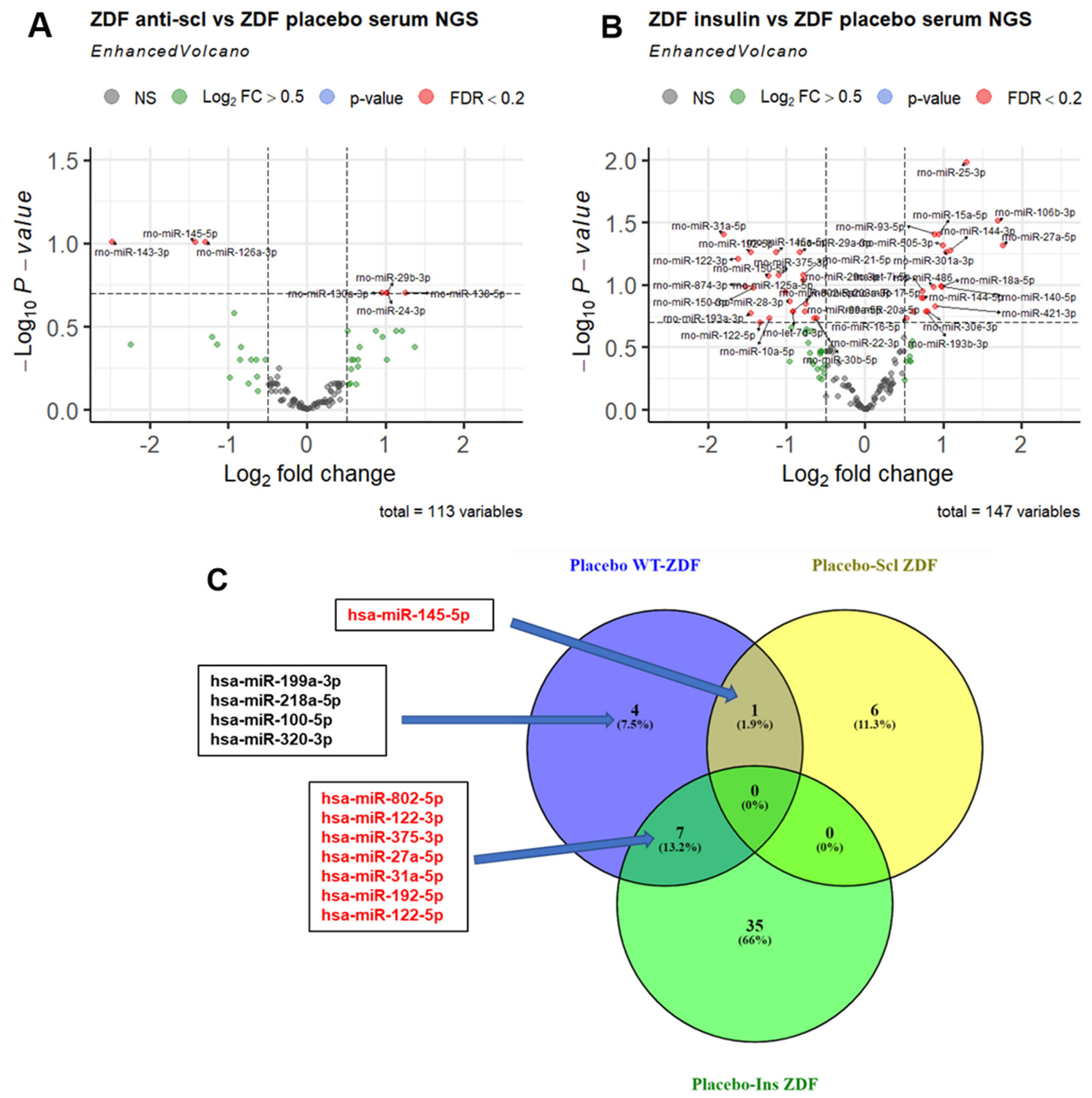
2.4. Effect of PTH, Anti-Sclerostin, and Insulin Treatment on MiRNA Serum Levels and Bone Tissue Expression
2.5. Anti-Scl and Insulin Treatment Rescue Serum Levels of Different MiRNAs in ZDF Rats
2.6. Gene Target and Network Analysis Comprises Pathways Involved in Bone Biology and Osteoporosis Potentially Targeted by the Dysregulated MiRNAs in ZDF Rats in Serum and in Ulna Bone
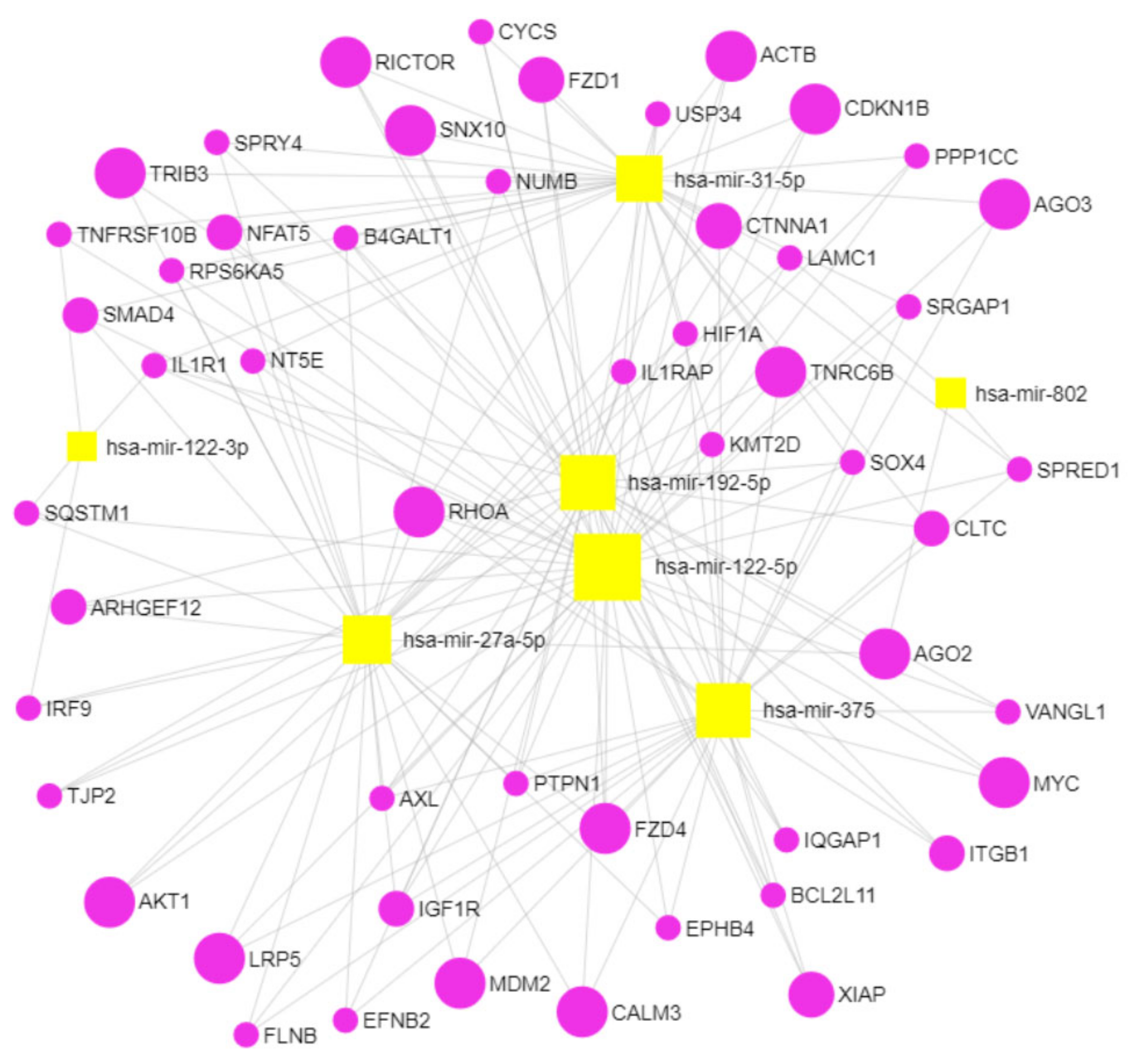
2.7. Gene Target and Network Analyses Reveal Several Pathways Involved in Bone Biology and Osteoporosis as Potential Targets for the Dysregulated MiRNAs in ZDF Rescued by Insulin
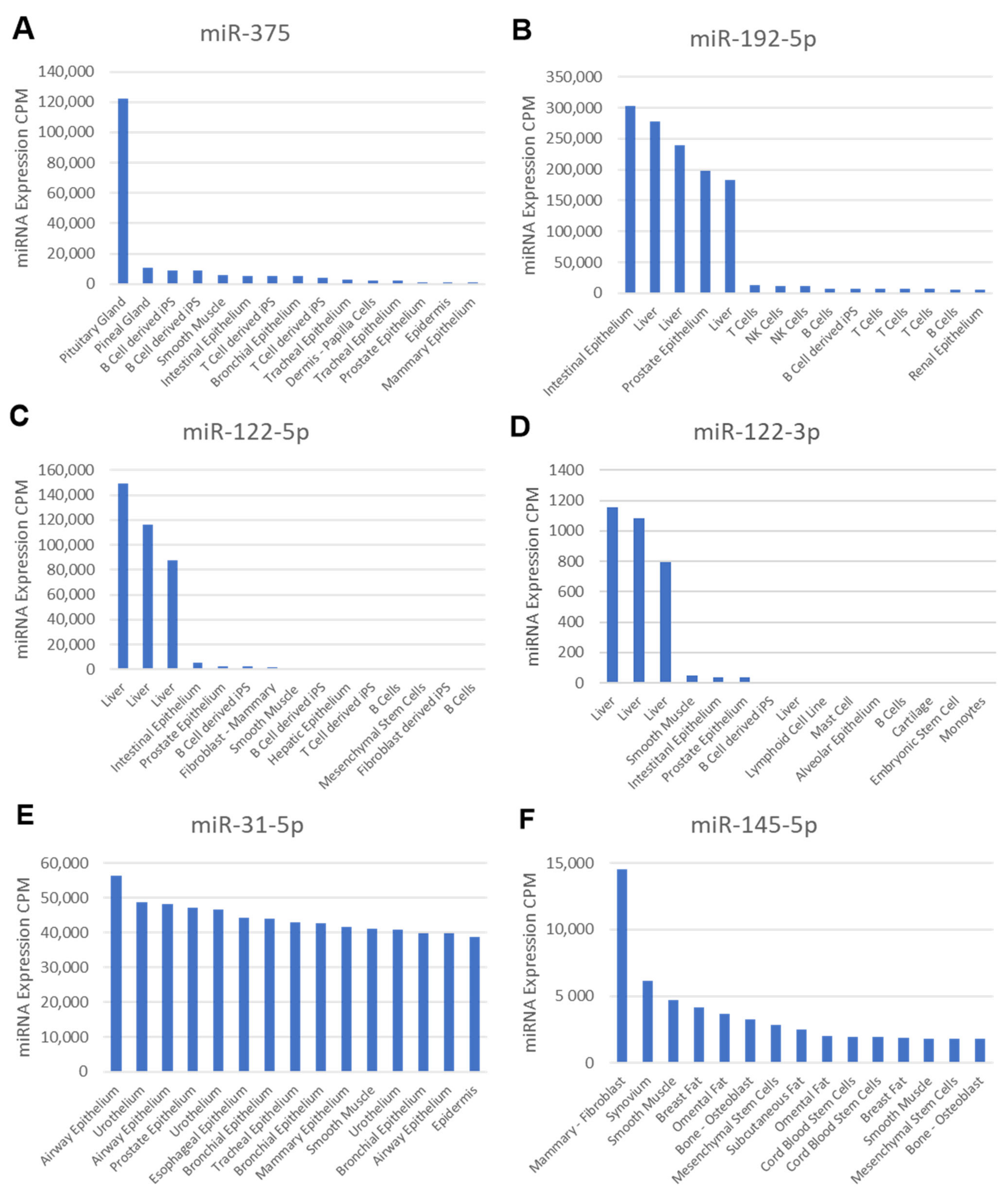
2.8. Cell-Type Enrichment Analysis Reveals Distinct Putative Donor Cell Types of the Dysregulated and Rescued MiRNAs in ZDF Rats
3. Discussion
3.1. MiR-199a-3p Is Jointly Downregulated in Serum and Bone Tissue of ZDF vs. WT Rats
3.2. Gene Network Analysis of Dysregulated MiRNAs in Serum or Bone of ZDF vs. WT Rats
3.3. Anti-Sclerostin Treatment
3.4. PTH Treatment
3.5. Insulin Treatment
3.6. MiRNAs Not Rescued by Any Treatment
3.7. Contribution to the Field and Limitations of This Study
4. Materials and Methods
4.1. Animals
4.2. RNA Extraction from Serum
4.3. RNA Extraction from Ulna Bone
4.4. Library Preparation for Small RNA-Seq
4.5. MiRNA Target Network Construction
4.6. Cell-Type Enrichment Analysis
4.7. Statistical Analysis
5. Conclusions
- -
- The previously described PTH resistance induced by diabetes in the ZDF rat model, with a blunting effect of T2DM over PTH.
- -
- The PTH dose probably being too high or too extended in time. This is also shown when compared to other studies that used ovariectomized (OVX) rats, resulting in a shorter time effect in miRNA patterns in ZDF rats compared to OVX rats.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Janghorbani, M.; van Dam, R.; Willett, W.C.; Hu, F.B. Systematic Review of Type 1 and Type 2 Diabetes Mellitus and Risk of Fracture. Am. J. Epidemiol. 2007, 166, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Grillari, J.; Mäkitie, R.E.; Kocijan, R.; Haschka, J.; Vázquez, D.C.; Semmelrock, E.; Hackl, M. Circulating miRNAs in bone health and disease. Bone 2021, 145, 115787. [Google Scholar] [CrossRef] [PubMed]
- Hackl, M.; Heilmeier, U.; Weilner, S.; Grillari, J. Circulating microRNAs as novel biomarkers for bone diseases—Complex signatures for multifactorial diseases? Mol. Cell. Endocrinol. 2016, 432, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Weilner, S.; Skalicky, S.; Salzer, B.; Keider, V.; Wagner, M.; Hildner, F.; Gabriel, C.; Dovjak, P.; Pietschmann, P.; Grillari-Voglauer, R.; et al. Differentially circulating miRNAs after recent osteoporotic fractures can influence osteogenic differentiation. Bone 2015, 79, 43–51. [Google Scholar] [CrossRef]
- Takahara, S.; Lee, S.Y.; Iwakura, T.; Oe, K.; Fukui, T.; Okumachi, E.; Waki, T.; Arakura, M.; Sakai, Y.; Nishida, K.; et al. Altered expression of microRNA during fracture healing in diabetic rats. Bone Jt. Res. 2018, 7, 139–147. [Google Scholar] [CrossRef]
- Tang, Y.; Zheng, L.; Zhou, J.; Chen, Y.; Yang, L.; Deng, F.; Hu, Y. miR-203-3p participates in the suppression of diabetes-associated osteogenesis in the jaw bone through targeting Smad. Int. J. Mol. Med. 2018, 41, 1595–1607. [Google Scholar] [CrossRef]
- Grieco, G.E.; Cataldo, D.; Ceccarelli, E.; Nigi, L.; Catalano, G.; Brusco, N.; Mancarella, F.; Ventriglia, G.; Fondelli, C.; Guarino, E.; et al. Serum Levels of miR-148a and miR-21-5p Are Increased in Type 1 Diabetic Patients and Correlated with Markers of Bone Strength and Metabolism. Non-Coding RNA 2018, 4, 37. [Google Scholar] [CrossRef]
- Heilmeier, U.; Hackl, M.; Skalicky, S.; Weilner, S.; Schroeder, F.; Vierlinger, K.; Patsch, J.M.; Baum, T.; Oberbauer, E.; Lobach, I.; et al. Serum miRNA Signatures Are Indicative of Skeletal Fractures in Postmenopausal Women with and without Type 2 Diabetes and Influence Osteogenic and Adipogenic Differentiation of Adipose Tissue-Derived Mesenchymal Stem Cells In Vitro. J. Bone Miner. Res. 2016, 31, 2173–2192. [Google Scholar] [CrossRef]
- Heilmeier, U.; Hackl, M.; Schroeder, F.; Torabi, S.; Kapoor, P.; Vierlinger, K.; Eiriksdottir, G.; Gudmundsson, E.F.; Harris, T.B.; Gudnason, V.; et al. Circulating serum microRNAs including senescent miR-31-5p are associated with incident fragility fractures in older postmenopausal women with type 2 diabetes mellitus. Bone 2022, 158, 116308. [Google Scholar] [CrossRef]
- Hamann, C.; Goettsch, C.; Mettelsiefen, J.; Henkenjohann, V.; Rauner, M.; Hempel, U.; Bernhardt, R.; Fratzl-Zelman, N.; Roschger, P.; Rammelt, S.; et al. Delayed bone regeneration and low bone mass in a rat model of insulin-resistant type 2 diabetes mellitus is due to impaired osteoblast function. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E1220–E1228. [Google Scholar] [CrossRef]
- Hamann, C.; Rauner, M.; Höhna, Y.; Bernhardt, R.; Mettelsiefen, J.; Goettsch, C.; Günther, K.-P.; Stolina, M.; Han, C.-Y.; Asuncion, F.J.; et al. Sclerostin antibody treatment improves bone mass, bone strength, and bone defect regeneration in rats with type 2 diabetes mellitus. J. Bone Miner. Res. 2013, 28, 627–638. [Google Scholar] [CrossRef]
- Hamann, C.; Picke, A.-K.; Campbell, G.M.; Balyura, M.; Rauner, M.; Bernhardt, R.; Huber, G.; Morlock, M.; Günther, K.-P.; Bornstein, S.R.; et al. Effects of Parathyroid Hormone on Bone Mass, Bone Strength, and Bone Regeneration in Male Rats with Type 2 Diabetes Mellitus. Endocrinology 2014, 155, 1197–1206. [Google Scholar] [CrossRef]
- Picke, A.-K.; Alaguero, I.G.; Campbell, G.; Glüer, C.-C.; Salbach-Hirsch, J.; Rauner, M.; Hofbauer, L. Bone defect regeneration and cortical bone parameters of type 2 diabetic rats are improved by insulin therapy. Bone 2016, 82, 108–115. [Google Scholar] [CrossRef]
- Zhu, G.; Pei, L.; Lin, F.; Yin, H.; Li, X.; He, W.; Liu, N.; Gou, X. Exosomes from human-bone-marrow-derived mesenchymal stem cells protect against renal ischemia/reperfusion injury via transferring miR-199a-3p. J. Cell. Physiol. 2019, 234, 23736–23749. [Google Scholar] [CrossRef]
- Shuai, Y.; Yang, R.; Mu, R.; Yu, Y.; Rong, L.; Jin, L. MiR-199a-3p mediates the adipogenic differentiation of bone marrow-derived mesenchymal stem cells by regulating KDM6A/WNT signaling. Life Sci. 2019, 220, 84–91. [Google Scholar] [CrossRef]
- Wu, J.-C.; Sun, J.; Xu, J.-C.; Zhou, Z.-Y.; Zhang, Y.-F. Down-regulated microRNA-199a-3p enhances osteogenic differentiation of bone marrow mesenchymal stem cells by targeting Kdm3a in ovariectomized rats. Biochem. J. 2021, 478, 721–734. [Google Scholar] [CrossRef]
- Fu, J.; Hao, L.; Tian, Y.; Liu, Y.; Gu, Y.; Wu, J. miR-199a-3p is involved in estrogen-mediated autophagy through the IGF-1/mTOR pathway in osteocyte-like MLO-Y4 cells. J. Cell. Physiol. 2018, 233, 2292–2303. [Google Scholar] [CrossRef] [PubMed]
- Hallenborg, P.; Jensen, B.A.H.; Fjære, E.; Petersen, R.K.; Belmaâti, M.-S.; Rasmussen, S.S.; Gunnarsson, J.P.; Lauritzen, P.; Cheng, K.K.Y.; Hermansson, M.; et al. Adipose MDM2 regulates systemic insulin sensitivity. Sci. Rep. 2021, 11, 21839. [Google Scholar] [CrossRef] [PubMed]
- Olivos, D.J.; Perrien, D.S.; Hooker, A.; Cheng, Y.; Fuchs, R.K.; Hong, J.M.; Bruzzaniti, A.; Chun, K.; Eischen, C.M.; Kacena, M.A.; et al. The proto-oncogene function of Mdm2 in bone. J. Cell. Biochem. 2018, 119, 8830–8840. [Google Scholar] [CrossRef]
- Lalrohlui, F.; Zohmingthanga, J.; Hruaii, V.; Vanlallawma, A.; Kumar, N.S. Whole exome sequencing identifies the novel putative gene variants related with type 2 diabetes in Mizo population, northeast India. Gene 2021, 769, 145229. [Google Scholar] [CrossRef]
- Karasik, D.; Zillikens, M.C.; Hsu, Y.-H.; Aghdassi, A.; Akesson, K.; Amin, N.; Barroso, I.; A Bennett, D.; Bertram, L.; Bochud, M.; et al. Disentangling the genetics of lean mass. Am. J. Clin. Nutr. 2019, 109, 276–287. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X.; Yang, M.; Ruan, W.; Wei, W.; Gu, D.; Wang, J.; Guo, X.; Guo, L.; Yuan, Y. miR-145-5p Increases Osteoclast Numbers In Vitro and Aggravates Bone Erosion in Collagen-Induced Arthritis by Targeting Osteoprotegerin. Med. Sci. Monit. 2018, 24, 5292–5300. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Tian, Q.; Ling, S.; Liu, Y.; Yang, S.; Shao, Z. miR-145 suppresses osteogenic differentiation by targeting Sp7. FEBS Lett. 2013, 587, 3027–3031. [Google Scholar] [CrossRef]
- Jin, Y.; Hong, F.; Bao, Q.; Xu, Q.; Duan, R.; Zhu, Z.; Zhang, W.; Ma, C. MicroRNA-145 suppresses osteogenic differentiation of human jaw bone marrow mesenchymal stem cells partially via targeting semaphorin 3A. Connect. Tissue Res. 2020, 61, 577–585. [Google Scholar] [CrossRef]
- Kocijan, R.; Weigl, M.; Skalicky, S.; Geiger, E.; Ferguson, J.; Leinfellner, G.; Heimel, P.; Pietschmann, P.; Grillari, J.; Redl, H.; et al. MicroRNA levels in bone and blood change during bisphosphonate and teriparatide therapy in an animal model of postmenopausal osteoporosis. Bone 2020, 131, 115104. [Google Scholar] [CrossRef]
- Kim, B.; Cho, Y.J.; Lim, W. Osteoporosis therapies and their mechanisms of action (Review). Exp. Ther. Med. 2021, 22, 1379. [Google Scholar] [CrossRef]
- Chen, S.; Yang, L.; He, S.; Yang, J.; Liu, D.; Bao, Q.; Qin, H.; Du, W.; Zhong, X.; Chen, C.; et al. Preactivation of β-catenin in osteoblasts improves the osteoanabolic effect of PTH in type 1 diabetic mice. J. Cell. Physiol. 2020, 235, 1480–1493. [Google Scholar] [CrossRef]
- Leanza, G.; Fontana, F.; Lee, S.; Remedi, M.S.; Schott, C.; Ferron, M.; Hamilton-Hall, M.; Alippe, Y.; Strollo, R.; Napoli, N.; et al. Gain-of-Function Lrp5 Mutation Improves Bone Mass and Strength and Delays Hyperglycemia in a Mouse Model of Insulin-Deficient Diabetes. J. Bone Miner. Res. 2021, 36, 1403–1415. [Google Scholar] [CrossRef]
- Gu, C.; Xu, Y.; Zhang, S.; Guan, H.; Song, S.; Wang, X.; Wang, Y.; Li, Y.; Zhao, G. miR-27a attenuates adipogenesis and promotes osteogenesis in steroid-induced rat BMSCs by targeting PPARγ and GREM. Sci. Rep. 2016, 6, 38491. [Google Scholar] [CrossRef]
- Li, X.; Chen, R.; Li, Y.; Wang, P.; Cui, Y.; Yang, L.; Zhu, X.; Zhang, R. miR-27a-5p—Abundant Small Extracellular Vesicles Derived from Epimedium-Preconditioned Bone Mesenchymal Stem Cells Stimulate Osteogenesis by Targeting Atg4B-Mediated Autophagy. Front. Cell Dev. Biol. 2021, 9, 642646. [Google Scholar] [CrossRef]
- Zhang, F.; Ma, D.; Zhao, W.; Wang, D.; Liu, T.; Liu, Y.; Yang, Y.; Liu, Y.; Mu, J.; Li, B.; et al. Obesity-induced overexpression of miR-802 impairs insulin transcription and secretion. Nat. Commun. 2020, 11, 1822. [Google Scholar] [CrossRef]
- Higuchi, C.; Nakatsuka, A.; Eguchi, J.; Teshigawara, S.; Kanzaki, M.; Katayama, A.; Yamaguchi, S.; Takahashi, N.; Murakami, K.; Ogawa, D.; et al. Identification of Circulating miR-101, miR-375 and miR-802 as Biomarkers for Type 2 Diabetes. Metabolism 2015, 64, 489–497. [Google Scholar] [CrossRef]
- Cao, Z.-Q.; Shen, Z.; Huang, W.-Y. MicroRNA-802 Promotes Osteosarcoma Cell Proliferation by Targeting p27. Asian Pac. J. Cancer Prev. 2013, 14, 7081–7084. [Google Scholar] [CrossRef]
- Li, X. miR-375, a microRNA related to diabetes. Gene 2014, 533, 1–4. [Google Scholar] [CrossRef]
- Jiménez-Lucena, R.; Camargo, A.; Alcala-Diaz, J.F.; Romero-Baldonado, C.; Luque, R.M.; Van Ommen, B.; Delgado-Lista, J.; Ordovás, J.M.; Pérez-Martínez, P.; Rangel-Zuñiga, O.A.; et al. A plasma circulating miRNAs profile predicts type 2 diabetes mellitus and prediabetes: From the CORDIOPREV study. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef]
- García-Jacobo, R.E.; Uresti-Rivera, E.E.; Portales-Pérez, D.P.; González-Amaro, R.; Lara-Ramírez, E.E.; Enciso-Moreno, J.A.; García-Hernández, M.H. Circulating miR-146a, miR-34a and miR-375 in type 2 diabetes patients, pre-diabetic and normal-glycaemic individuals in relation to β-cell function, insulin resistance and metabolic parameters. Clin. Exp. Pharmacol. Physiol. 2019, 46, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hong, N.; Kim, Y.; Park, S.; Kim, K.-J.; Jeong, J.; Jung, H.-I.; Rhee, Y. Circulating miR-122-5p and miR-375 as Potential Biomarkers for Bone Mass Recovery after Parathyroidectomy in Patients with Primary Hyperparathyroidism: A Proof-of-Concept Study. Diagnostics 2021, 11, 1704. [Google Scholar] [CrossRef]
- Du, F.; Wu, H.; Zhou, Z.; Liu, Y. microRNA-375 inhibits osteogenic differentiation by targeting runt-related transcription factor 2. Exp. Ther. Med. 2015, 10, 207–212. [Google Scholar] [CrossRef]
- Lei, N.B.; Liang, X.; Wang, P.; Liu, Q.; Wang, W.G. Teriparatide alleviates osteoporosis by promoting osteogenic differentiation of hMSCs via miR-375/RUNX2 axis. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 11043–11050. [Google Scholar]
- Chen, S.; Zheng, Y.; Zhang, S.; Jia, L.; Zhou, Y. Promotion Effects of miR-375 on the Osteogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells. Stem Cell Rep. 2017, 8, 773–786. [Google Scholar] [CrossRef]
- Chen, S.; Tang, Y.; Liu, Y.; Zhang, P.; Lv, L.; Zhang, X.; Jia, L.; Zhou, Y. Exosomes derived from miR-375-overexpressing human adipose mesenchymal stem cells promote bone regeneration. Cell Prolif. 2019, 52, e12669. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Hong, J.; Cao, Y.; Shi, J.; Gu, W.; Ning, G.; Zhang, Y.; Wang, W. Elevated circulating microRNA-122 is associated with obesity and insulin resistance in young adults. Eur. J. Endocrinol. 2015, 172, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Pastukh, N.; Meerson, A.; Kalish, D.; Jabaly, H.; Blum, A. Serum miR-122 levels correlate with diabetic retinopathy. Clin. Exp. Med. 2019, 19, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Willeit, P.; Skroblin, P.; Moschen, A.R.; Yin, X.; Kaudewitz, D.; Zampetaki, A.; Barwari, T.; Whitehead, M.; Ramírez, C.M.; Goedeke, L.; et al. Circulating MicroRNA-122 Is Associated With the Risk of New-Onset Metabolic Syndrome and Type 2 Diabetes. Diabetes 2017, 66, 347–357. [Google Scholar] [CrossRef]
- Hao, W.; Liu, H.-Z.; Zhou, L.-G.; Sun, Y.-J.; Su, H.; Ni, J.-Q.; He, T.; Shi, P.; Wang, X. MiR-122-3p regulates the osteogenic differentiation of mouse adipose-derived stem cells via Wnt/β catenin signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3892–3898. [Google Scholar]
- Yang, Y.; Shen, Z.; Sun, W.; Gao, S.; Li, Y.; Guo, Y. The role of miR-122-5p in negatively regulating T-box brain 1 expression on the differentiation of mouse bone mesenchymal stem cells. Neuroreport 2017, 28, 367–374. [Google Scholar] [CrossRef]
- Meng, Y.-C.; Lin, T.; Jiang, H.; Zhang, Z.; Shu, L.; Yin, J.; Ma, X.; Wang, C.; Gao, R.; Zhou, X.-H. miR-122 Exerts Inhibitory Effects on Osteoblast Proliferation/Differentiation in Osteoporosis by Activating the PCP4-Mediated JNK Pathway. Mol. Ther. Nucleic Acids 2020, 20, 345–358. [Google Scholar] [CrossRef]
- Mandourah, A.Y.; Ranganath, L.; Barraclough, R.; Vinjamuri, S.; Hof, R.V.; Hamill, S.; Czanner, G.; Dera, A.A.; Wang, D.; Barraclough, D. Circulating microRNAs as potential diagnostic biomarkers for osteoporosis. Sci. Rep. 2018, 8, 8421. [Google Scholar] [CrossRef]
- Ren, F.J.; Yao, Y.; Cai, X.Y.; Fang, G.Y. Emerging Role of MiR-192-5p in Human Diseases. Front. Pharmacol. 2021, 12, 614068. [Google Scholar] [CrossRef]
- Chen, X.; Zhong, L.; Li, X.; Liu, W.; Zhao, Y.; Li, J. Down-regulation of microRNA-31-5p inhibits proliferation and invasion of osteosarcoma cells through Wnt/β-catenin signaling pathway by enhancing AXIN. Exp. Mol. Pathol. 2019, 108, 32–41. [Google Scholar] [CrossRef]
- Xu, R.; Shen, X.; Si, Y.; Fu, Y.; Zhu, W.; Xiao, T.; Fu, Z.; Zhang, P.; Cheng, J.; Jiang, H. MicroRNA-31a-5p from aging BMSCs links bone formation and resorption in the aged bone marrow microenvironment. Aging Cell 2018, 17, e12794. [Google Scholar] [CrossRef]
- Weilner, S.; Schraml, E.; Wieser, M.; Messner, P.; Schneider, K.H.; Wassermann, K.; Micutkova, L.; Fortschegger, K.; Maier, A.; Westendorp, R.; et al. Secreted microvesicular miR-31 inhibits osteogenic differentiation of mesenchymal stem cells. Aging Cell 2016, 15, 744–754. [Google Scholar] [CrossRef]
- Sampilo, N.F.; Stepicheva, N.A.; Song, J.L. microRNA-31 regulates skeletogenesis by direct suppression of Eve and Wnt. Dev. Biol. 2021, 472, 98–114. [Google Scholar] [CrossRef]
- Materozzi, M.; Merlotti, D.; Gennari, L.; Bianciardi, S. The Potential Role of miRNAs as New Biomarkers for Osteoporosis. Int. J. Endocrinol. 2018, 2018, 2342860. [Google Scholar] [CrossRef]
- Mäkitie, E.R.; Hackl, M.; Niinimäki, R.; Kakko, S.; Grillari, J.; Mäkitie, O. Altered MicroRNA Profile in Osteoporosis Caused by Impaired WNT Signaling. J. Clin. Endocrinol. Metab. 2018, 103, 1985–1996. [Google Scholar] [CrossRef]
- Khamina, K.; Diendorfer, A.B.; Skalicky, S.; Weigl, M.; Pultar, M.; Krammer, T.L.; Fournier, C.A.; Schofield, A.L.; Otto, C.; Smith, A.T.; et al. A MicroRNA Next-Generation-Sequencing Discovery Assay (miND) for Genome-Scale Analysis and Absolute Quantitation of Circulating MicroRNA Biomarkers. Int. J. Mol. Sci. 2022, 23, 1226. [Google Scholar] [CrossRef] [PubMed]
- De Rie, D.; Abugessaisa, I.; Alam, T.; Arner, E.; Arner, P.; Ashoor, H.; Åström, G.; Babina, M.; Bertin, N.; Burroughs, A.M.; et al. An integrated expression atlas of miRNAs and their promoters in human and mouse. Nat. Biotechnol. 2017, 35, 872–878. [Google Scholar] [CrossRef]
- Diendorfer, A.; Khamina, K.; Pultar, M.; Hackl, M. miND (miRNA NGS Discovery pipeline): A small RNA-seq analysis pipeline and report generator for microRNA biomarker discovery studies. F1000Research 2022, 11, 233. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Type of Treatment | Age at Beginning of Study | Treatment Administration | Treatment Duration | No. of Animals | Bone Defect | Serum/Ulna Collection at 12 Weeks | Original Reference |
|---|---|---|---|---|---|---|---|---|
| WT | Placebo | 11 weeks | s.c. (daily) | 12 weeks | 5 | yes | yes/yes | [12,13] |
| ZDF | Placebo | 11 weeks | s.c. (daily) | 12 weeks | 5 | yes | yes/yes | [12,13] |
| WT | Anti-Sclerostin | 11 weeks | s.c. (twice a week) 25 mg/kg | 12 weeks | 4 | yes | yes/no | [11] |
| ZDF | Anti-Sclerostin | 11 weeks | s.c. (twice a week) 25 mg/kg | 12 weeks | 4 | yes | yes/no | [11] |
| WT | PTH | 11 weeks | s.c. (5 times a week) 75 µg/kg | 12 weeks | 5 | yes | yes/yes | [12] |
| ZDF | PTH | 11 weeks | s.c. (5 times a week) 75 µg/kg | 12 weeks | 5 | yes | yes/yes | [12] |
| ZDF | Insulin | 11 weeks | s.c. (daily) 0.5 IU at week 1–13 IU at week 12 | 12 weeks | 4 | yes | yes/no | [13] |
| Placebo ZDF vs. Placebo WT | Treatment ZDF vs. Placebo ZDF | ||||
|---|---|---|---|---|---|
| Treatment | MiRNA | Log2FC | FDR | Log2FC | FDR |
| Anti-Scl | rno-miR-145-5p | 1.144 | 0.073 | −1.424 | 0.098 |
| Insulin | rno-miR-802-5p | 2.789 | 0.005 | −0.918 | 0.162 |
| rno-miR-27a-5p | −1.823 | 0.028 | 1.75 | 0.048 | |
| rno-miR-375-3p | 2.349 | 0.028 | −1.1 | 0.083 | |
| rno-miR-192-5p | 1.780 | 0.034 | −1.45 | 0.054 | |
| rno-miR-122-5p | 2.012 | 0.038 | −1.34 | 0.199 | |
| rno-miR-122-3p | 2.2 | 0.019 | −1.6 | 0.062 | |
| rno-miR-31a-5p | 1.7 | 0.034 | −1.8 | 0.039 | |
| Database | Pathway | Hits | p-Value | FDR |
|---|---|---|---|---|
| KEGG | Focal adhesion | CRK; THBS1; CCND1; CCND2; CRKL; IGF1R; VEGFA; JUN; FLNA; DIAPH1; PAK2; ACTG1 | 6.1 × 10−6 | 1.9 × 10−4 |
| TGF-beta signaling | EP300; THBS1; SMAD3; SP1; SMAD2; BMPR2 | 6.4 × 10−4 | 0.0076 | |
| Neurotrophin signaling | CRK; YWHAG; CRKL; JUN; YWHAQ; IRS4; YWHAB | 8.9 × 10−4 | 0.0076 | |
| Adherens junction | IGF1R; SMAD3; SMAD2; ACTG1 | 0.012 | 0.059 | |
| MAPK signaling | CRK; CRKL; JUN; FLNA; RAPGEF2; ELK4; HSPA1B; PAK2 | 0.019 | 0.080 | |
| Reactome | Cellular Senescence | CDK6; CDKN1B; E2F3; JUN; TNRC6A; SP1; AGO1; TNRC6B; CBX6; MDM2 | 9.6 × 10−6 | 1.1 × 10−4 |
| Signaling by NOTCH | EP300; CCND1; E2F3; TNRC6A; NOTCH2; AGO1; TNRC6B | 8.7 × 10−5 | 6.2 × 10−4 | |
| Signaling by TGF-beta Receptor Complex | CCNT2; SMAD3; SP1; SMAD2; XPO1; PARP1 | 2.5 × 10−4 | 0.0015 | |
| NGF signaling via TRKA | CDKN1B; CRK; CRKL; TNRC6A; AGO1; TNRC6B; MDM2; YWHAB | 0.0023 | 0.0074 | |
| VEGFA-VEGFR2 Pathway | CRK; VEGFA; PAK2; ACTG1; YWHAB | 0.0056 | 0.010 | |
| Signaling by SCF-KIT | CDKN1B; TNRC6A; AGO1; TNRC6B; MDM2; YWHAB | 0.0060 | 0.010 | |
| Signaling by NGF | CDKN1B; CRK; BCL2L11; CRKL; TNRC6A; AGO1; TNRC6B; MDM2; YWHAB | 0.0066 | 0.011 | |
| PI3K/AKT activation | CDKN1B; TNRC6A; AGO1; TNRC6B; MDM2 | 0.0070 | 0.011 | |
| DAP12 signaling | CDKN1B; TNRC6A; AGO1; TNRC6B; MDM2; YWHAB | 0.012 | 0.013 | |
| Signaling by EGFR | CDKN1B; TNRC6A; AGO1; TNRC6B; MDM2; YWHAB | 0.018 | 0.018 | |
| DisGeNET | Acro-Osteolysis | NOTCH2; ATL3 | 0.0029 | 0.010 |
| Pathological fracture | CCND1; NOTCH2 | 0.0066 | 0.010 |
| Database | Pathway | Hits | p-Value | FDR |
|---|---|---|---|---|
| KEGG | Adherens junction | RHOA, SMAD4, IGF1R, IQGAP1, PTPN1, CTNNA1, ACTB | 1.5 × 10−4 | 0.0055 |
| Apoptosis | AKT1, XIAP, IL1R1, IL1RAP, CYCS, TNFRSF10B | 0.0026 | 0.045 | |
| Wnt signaling | RHOA, SMAD4, FZD1, NFAT5, FZD4, VANGL1, LRP5, MYC | 0.0027 | 0.045 | |
| Axon guidance | RHOA, NFAT5, EFNB2, ITGB1, EPHB4, ARHGEF12, SRGAP1 | 0.0035 | 0.05 | |
| Focal adhesion | RHOA, AKT1, XIAP, IGF1R, ITGB1, ACTB, LAMC1, FLNB, PPP1CC | 0.0061 | 0.068 | |
| Jak-STAT signaling | SPRY4, SPRED1, AKT1, MYC, IRF9 | 0.025 | 0.14 | |
| Tight junction | RHOA, AKT1, TJP2, CTNNA1, ACTB | 0.048 | 0.21 | |
| Reactome | Signaling by Wnt | RHOA, SOX4, FZD1, TNRC6B, AKT1, XIAP, FZD4, USP34, AGO2, CALM3, AGO3, LRP5, MYC, CLTC, KMT2D | 7.2 × 10−5 | 0.0040 |
| PI3K/AKT activation | RHOA, TNRC6B, TRIB3, AKT1, CDKN1B, AGO2, AGO3, MDM2, RICTOR | 10−4 | 0.0040 | |
| Signaling by NGF | RHOA, TNRC6B, TRIB3, AKT1, CDKN1B, BCL2L11, AGO2, CALM3, RPS6KA5, AGO3, MDM2, SQSTM1, RICTOR, ARHGEF12 | 7.8 × 10−4 | 0.0049 | |
| Ca2+ pathway | TNRC6B, FZD4, AGO2, CALM3, AGO3 | 9.7 × 10−4 | 0.0049 | |
| Signaling by NOTCH | NUMB, TNRC6B, AGO2, B4GALT1 | 0.0013 | 0.053 | |
| Beta-catenin independent WNT signaling | RHOA, FZD1, TNRC6B, FZD4, AGO2, CALM3, AGO3, CLTC | 0.0015 | 0.0054 | |
| VEGFA-VEGFR2 Pathway | RHOA, AKT1, AXL, CALM3, CTNNA1, ACTB, RICTOR | 0.0012 | 0.0063 | |
| Signaling by FGFR | TNRC6B, TRIB3, AKT1, CDKN1B, AGO2, CALM3, AGO3, MDM2, RICTOR | 0.00257 | 0.0069 | |
| DAP12 signaling | TNRC6B, TRIB3, AKT1, CDKN1B, AGO2, CALM3, AGO3, MDM2, RICTOR | 0.0029 | 0.0074 | |
| Signaling by SCF-KIT | TNRC6B, TRIB3, AKT1, CDKN1B, AGO2, AGO3, MDM2, RICTOR | 0.0042 | 0.0095 | |
| DisGeNET | Abnormality of the vertebral column | LRP5, NT5E | 0.0010 | 0.010 |
| Osteopetrosis | SNX10, LRP5 | 0.0034 | 0.014 | |
| Varying degree of multiple fractures/increased fracture rate | FZD4, SNX10, LRP5 | 0.0123 | 0.018 | |
| Osteopetrosis, autosomal recessive 8 | SNX10 | 0.0188 | 0.018 | |
| Osteopetrosis, autosomal dominant 1/osteoporosis with pseudoglioma/bone mineral density quantitative trait locus 1 | LRP5 | 0.0188 | 0.018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carro Vázquez, D.; Emini, L.; Rauner, M.; Hofbauer, C.; Grillari, J.; Diendorfer, A.B.; Eastell, R.; Hofbauer, L.C.; Hackl, M. Effect of Anti-Osteoporotic Treatments on Circulating and Bone MicroRNA Patterns in Osteopenic ZDF Rats. Int. J. Mol. Sci. 2022, 23, 6534. https://doi.org/10.3390/ijms23126534
Carro Vázquez D, Emini L, Rauner M, Hofbauer C, Grillari J, Diendorfer AB, Eastell R, Hofbauer LC, Hackl M. Effect of Anti-Osteoporotic Treatments on Circulating and Bone MicroRNA Patterns in Osteopenic ZDF Rats. International Journal of Molecular Sciences. 2022; 23(12):6534. https://doi.org/10.3390/ijms23126534
Chicago/Turabian StyleCarro Vázquez, David, Lejla Emini, Martina Rauner, Christine Hofbauer, Johannes Grillari, Andreas B. Diendorfer, Richard Eastell, Lorenz C. Hofbauer, and Matthias Hackl. 2022. "Effect of Anti-Osteoporotic Treatments on Circulating and Bone MicroRNA Patterns in Osteopenic ZDF Rats" International Journal of Molecular Sciences 23, no. 12: 6534. https://doi.org/10.3390/ijms23126534