
Polypeptides Isolated from Lactococcus lactis Alleviates Lipopolysaccharide (LPS)-Induced Inflammation in Ctenopharyngodon idella

Abstract
:1. Introduction
2. Results
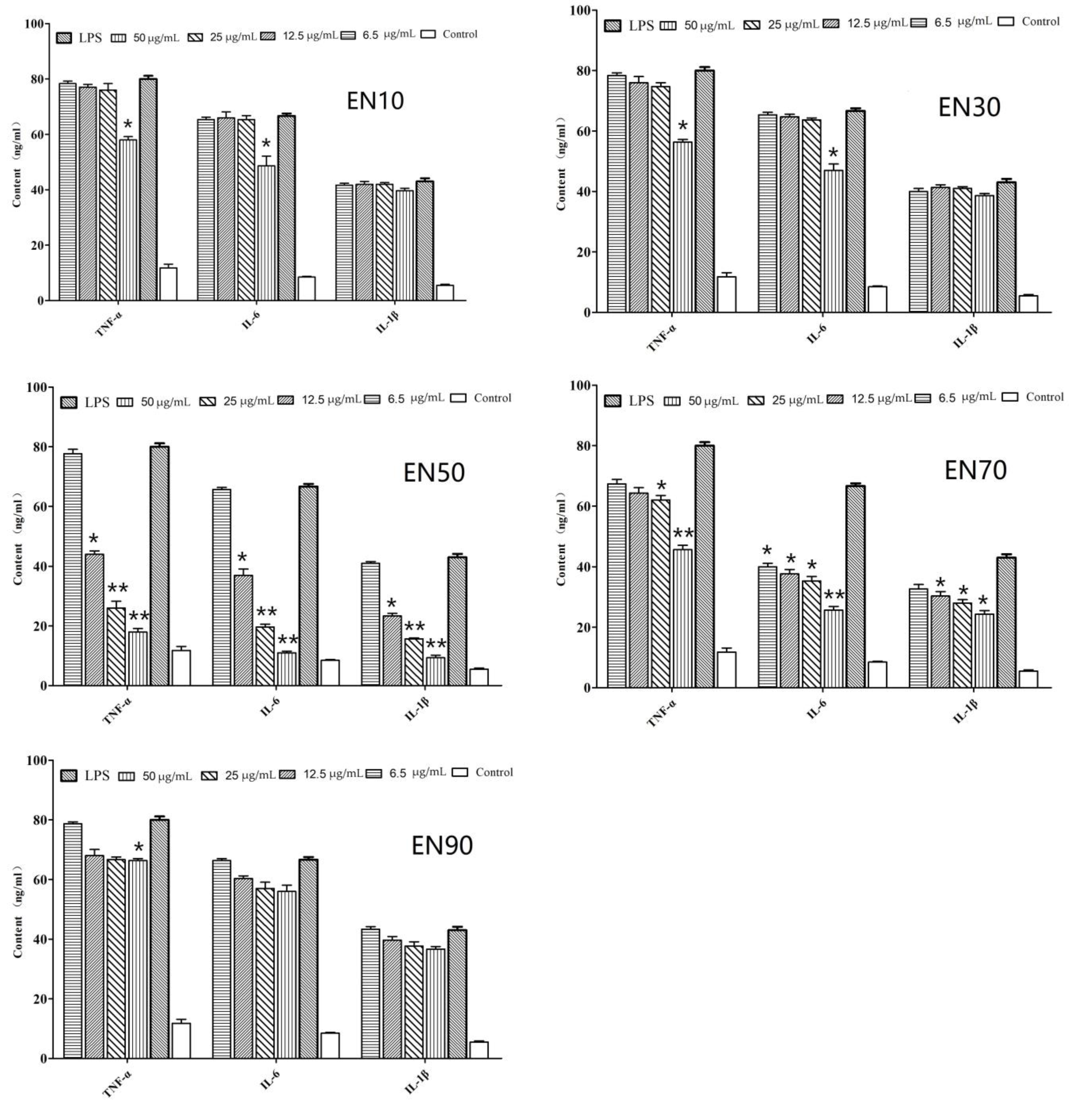
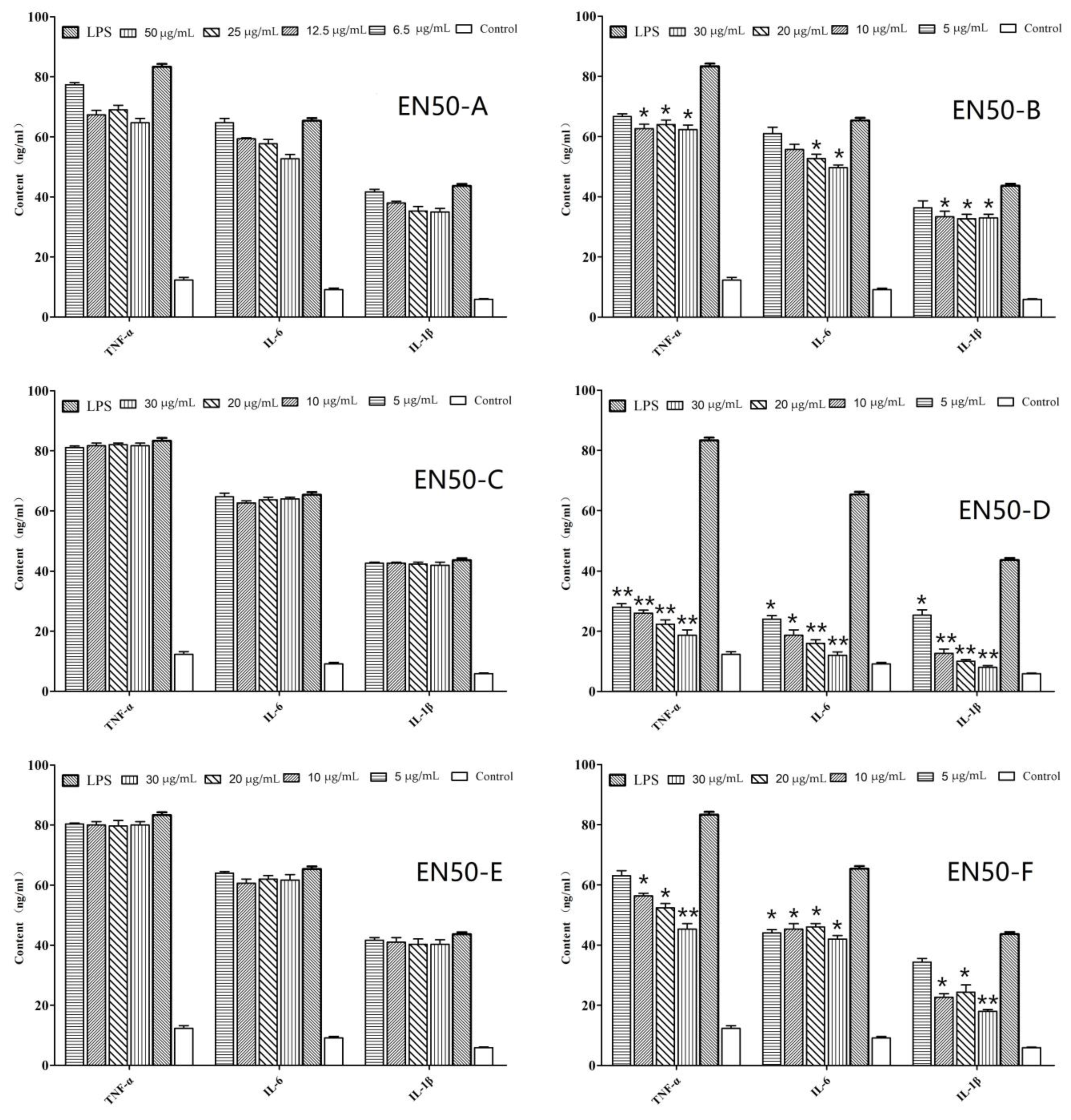
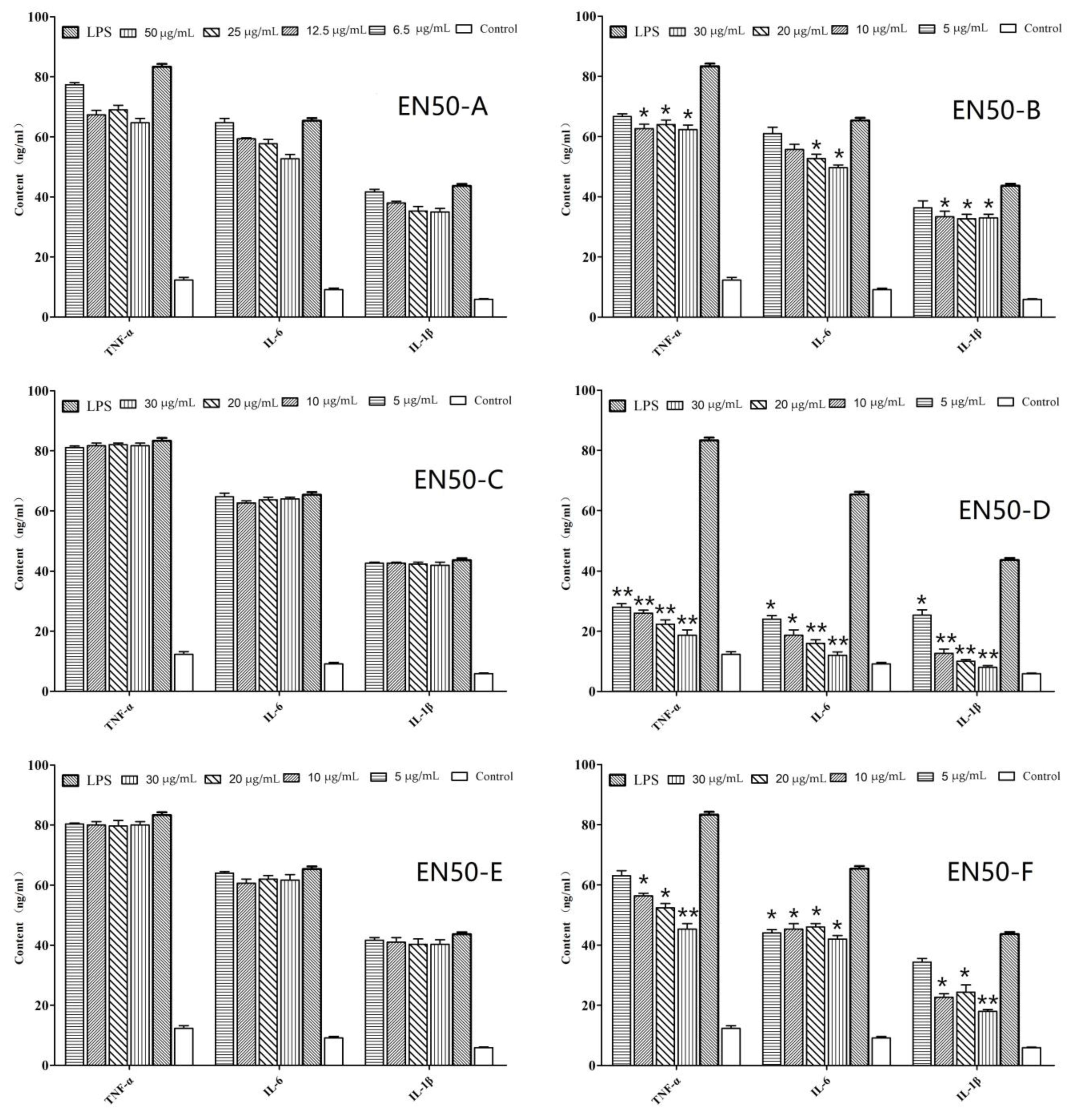
2.1. In Vitro Anti-Inflammatory Activity of Fraction and Active Peptides from L. lactis BL52
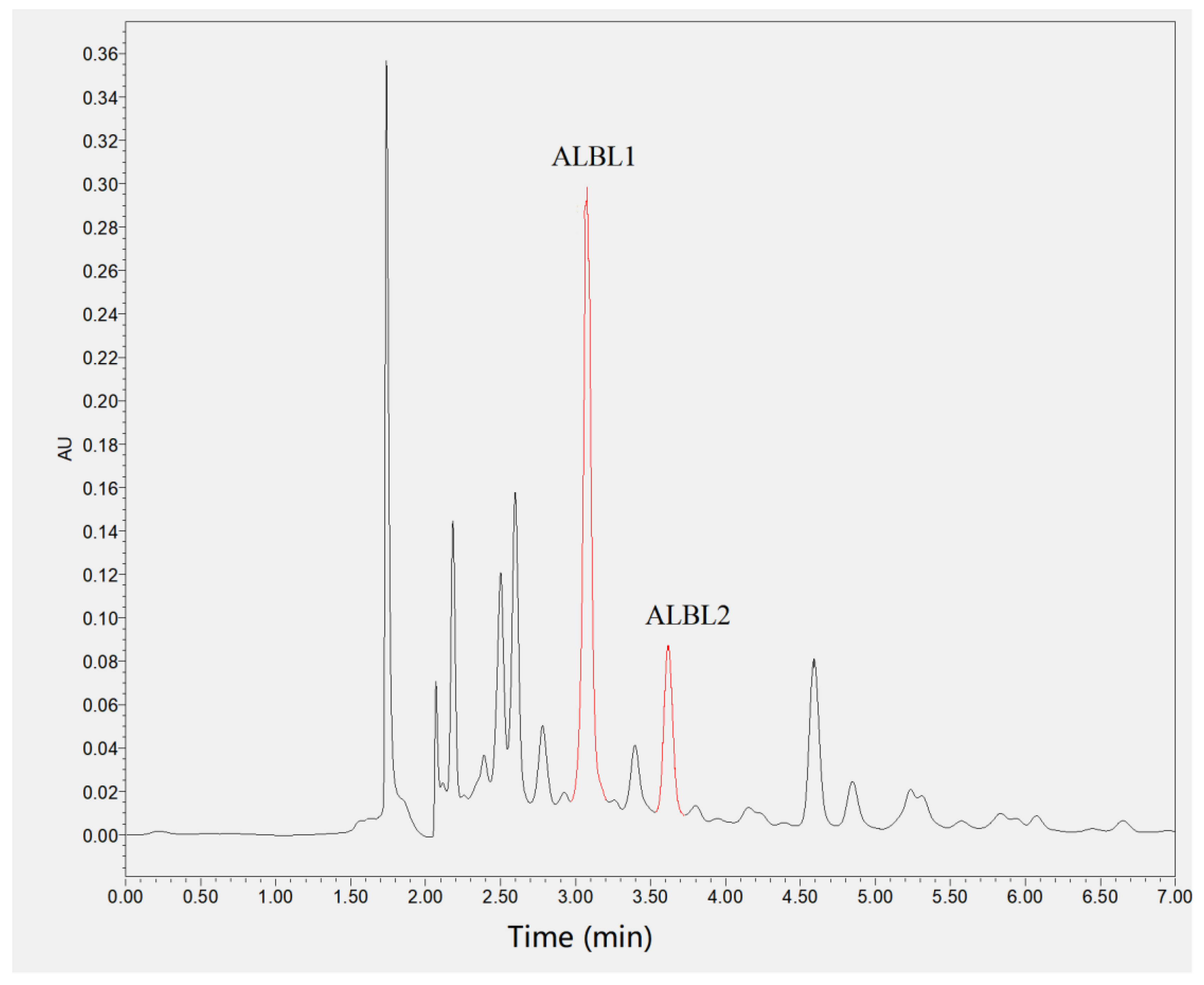
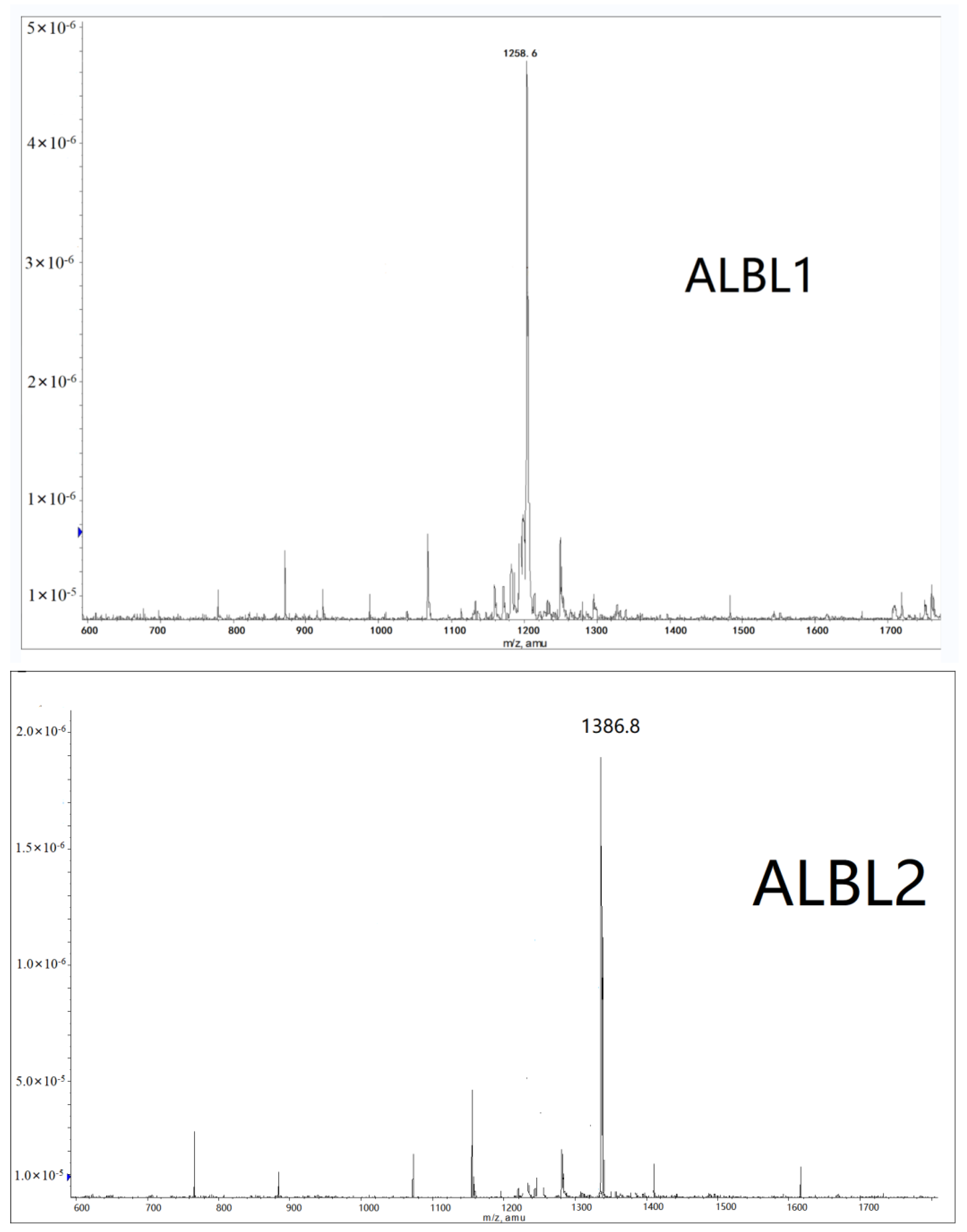
2.2. Mass Spectrometry Analysis
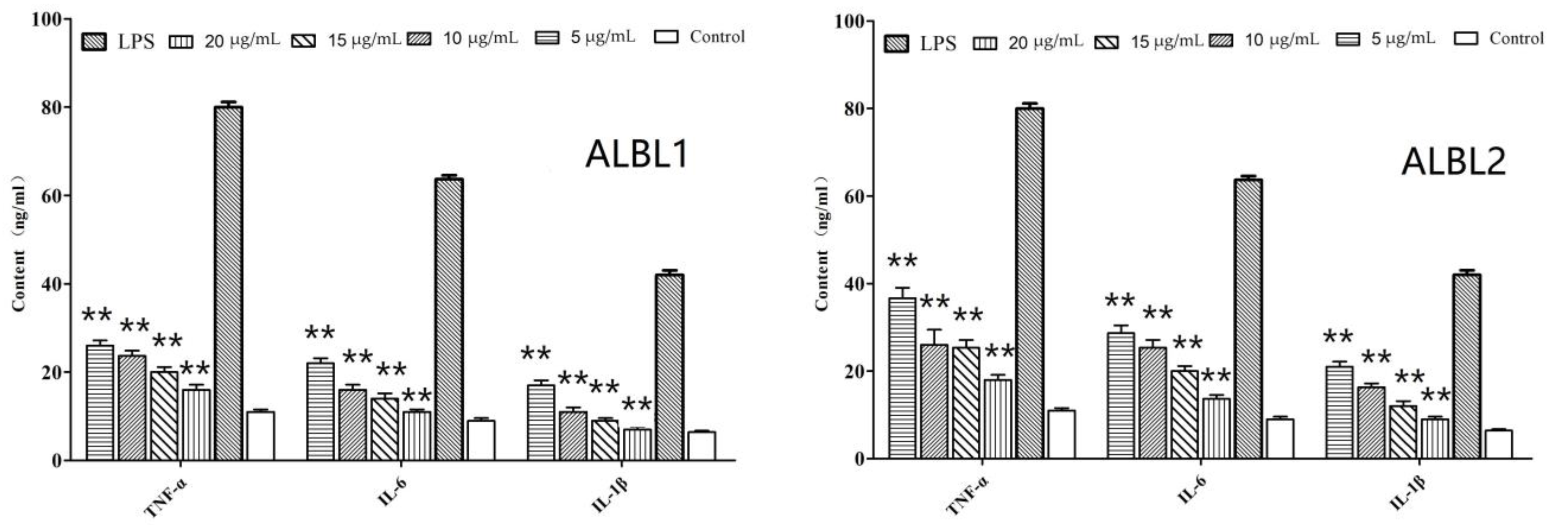
2.3. In Vivo Anti-Inflammatory Assay of ALBL1 and ALBL2
2.3.1. In Vivo Effects of ALBL1 and ALBL2 on Cytokine Production
2.3.2. In Vivo Effects of ALBL1 and ALBL2 on the Production of Inflammatory Mediators
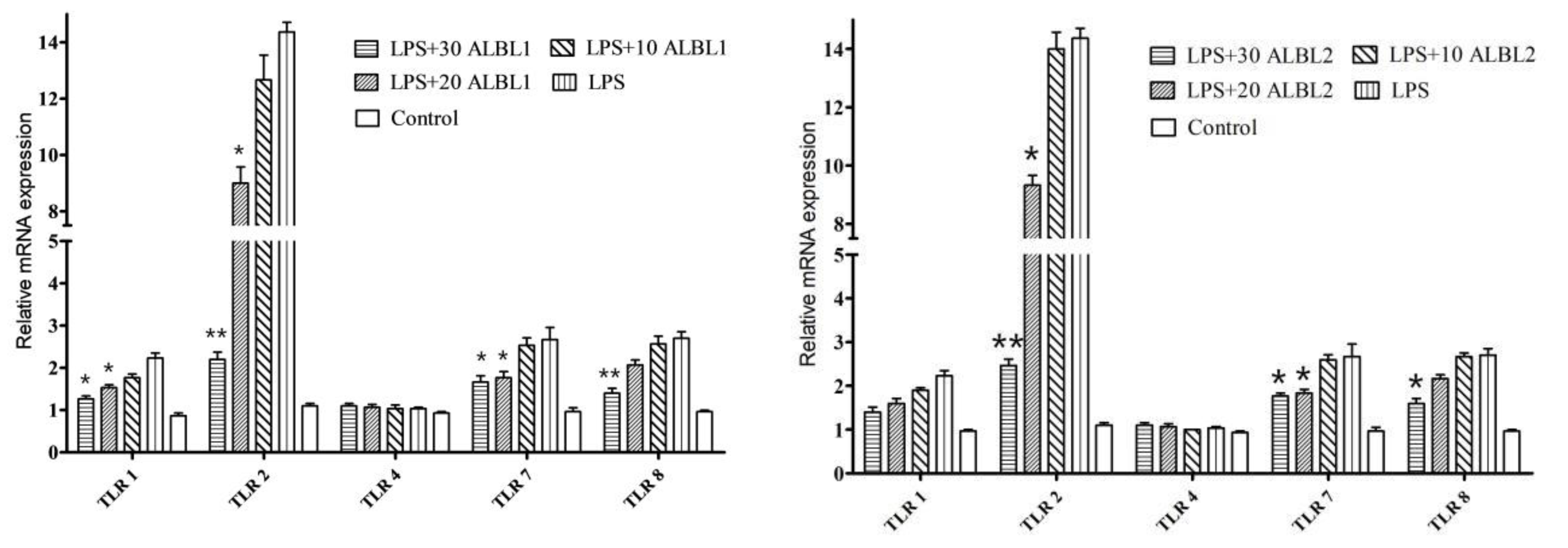
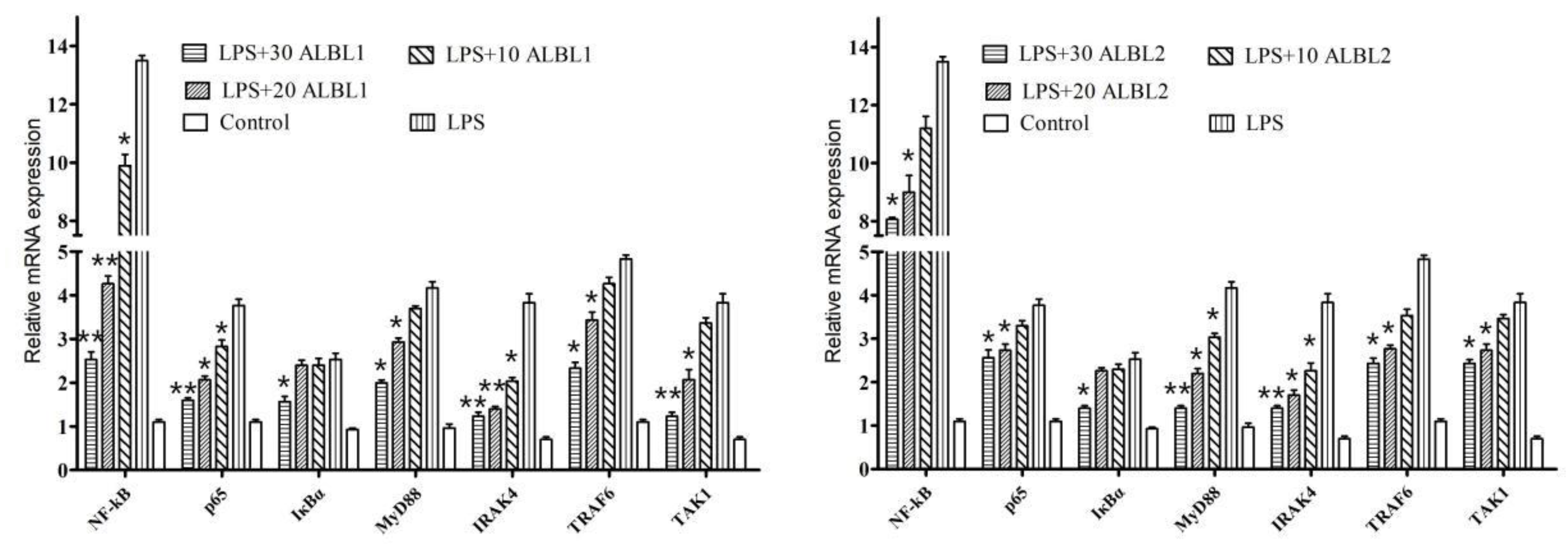
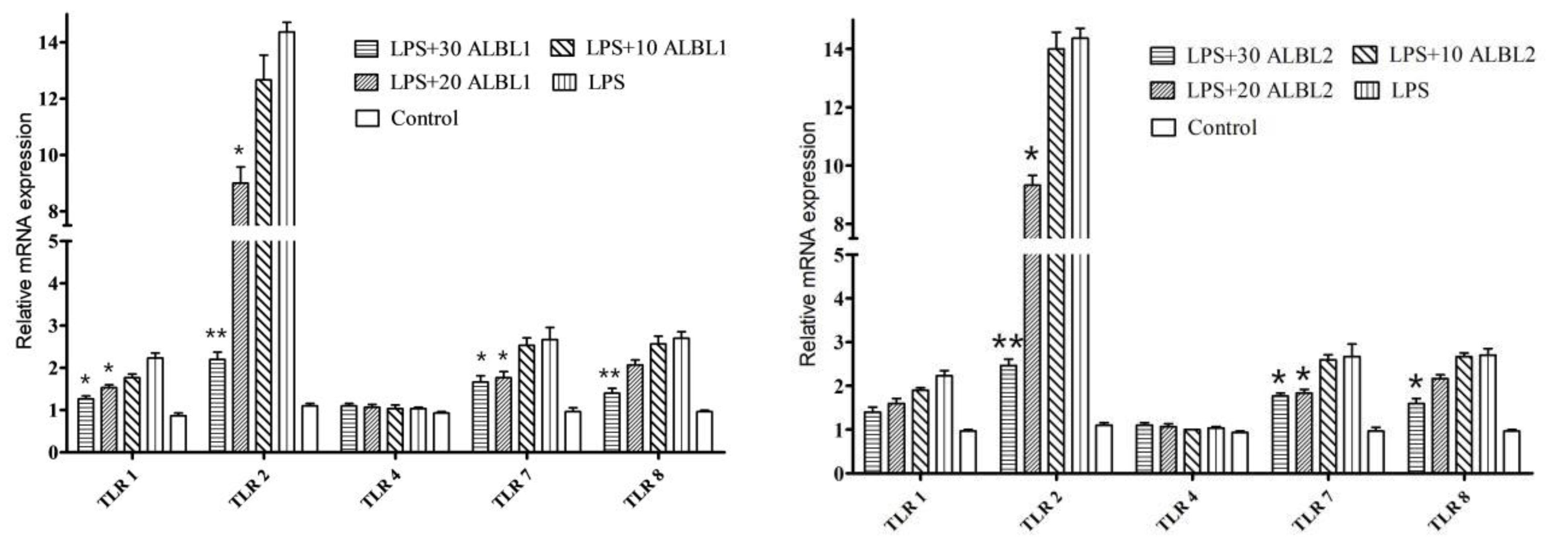
2.3.3. Expression of Toll/NF-KB Signal Pathways Related Genes
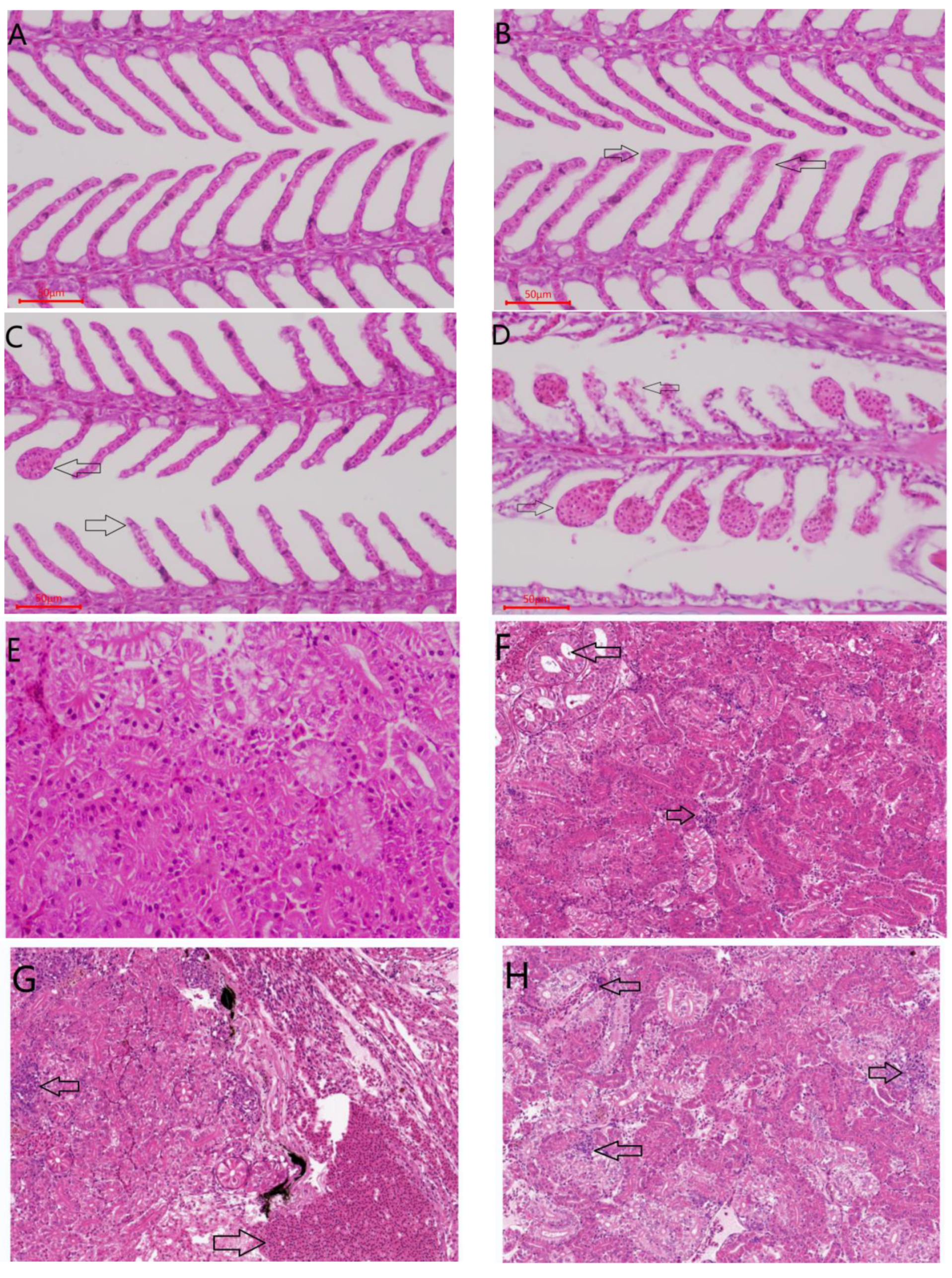
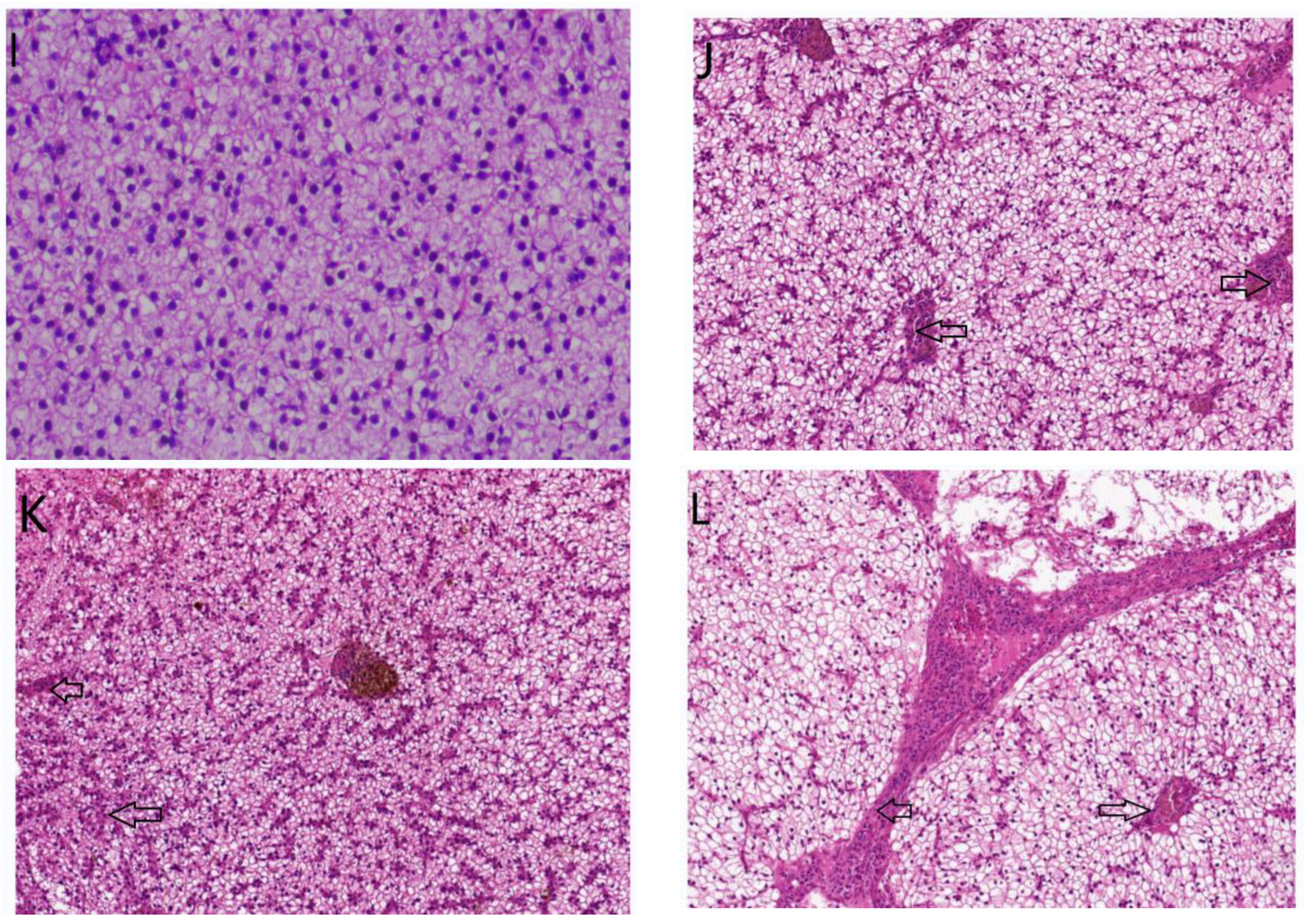
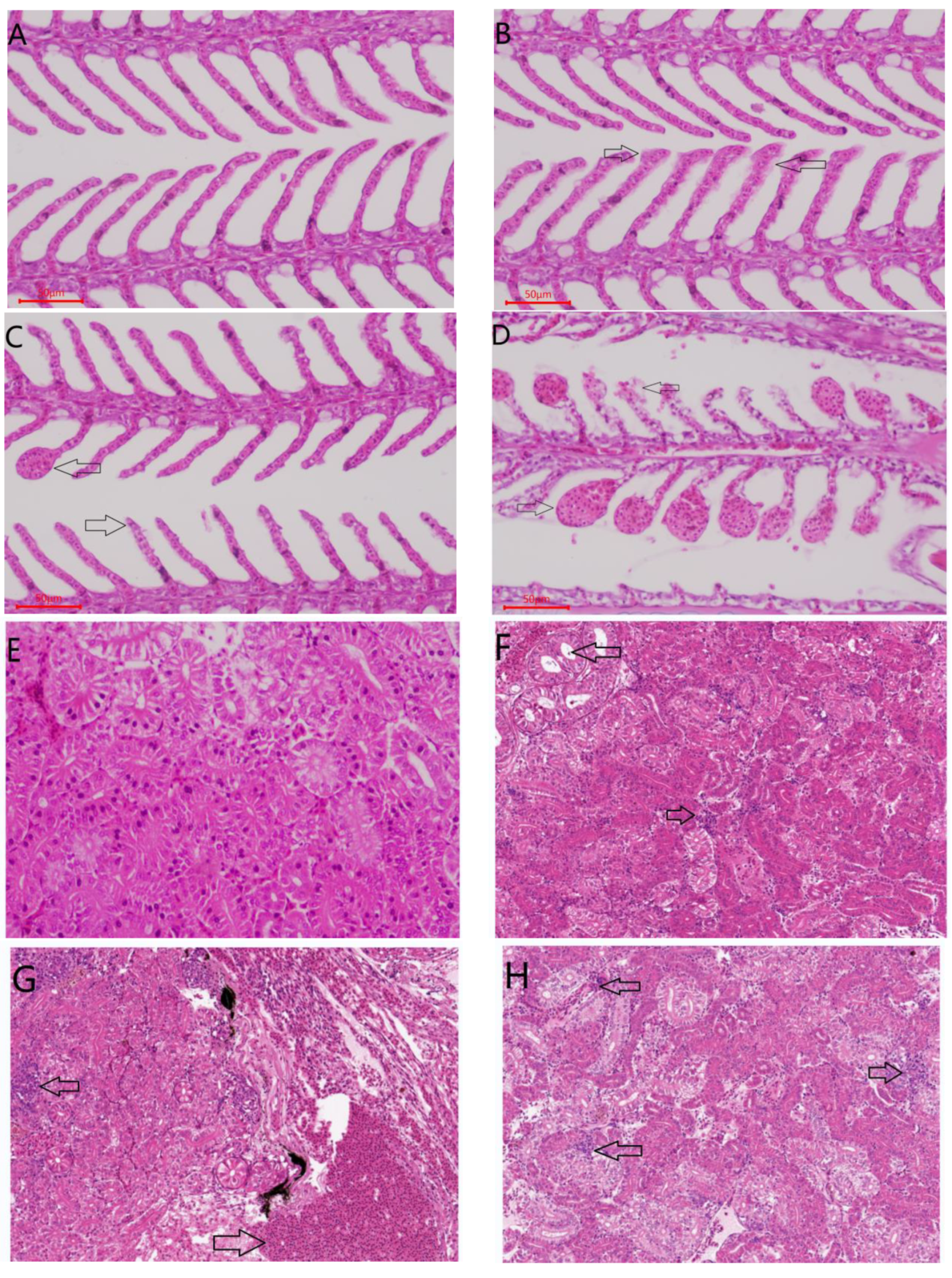
2.3.4. Histological Evaluation
3. Discussion
3.1. Peptide Inhibited LPS-Induced Inflammatory Cytokines
3.2. Peptide Inhibited Downregulated the Toll2/NF-κB Signaling Pathways
4. Material and Methods
4.1. Fish
4.2. In Vitro Study
4.3. Bioactivity-Guided Isolation of Active Compound from L. lactis BL52
4.4. Mass Spectrometry Analysis
4.5. In Vivo Experiment
4.5.1. Experimental Design
4.5.2. ELISA Assays of Inflammation
4.5.3. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.5.4. Histological Evaluation
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kalantar-Zadeh, K.; Ikizler, T.; Block, G.; Avram, M.M.; Kopple, J.D. Malnutrition-inflammation complex syndrome in dialysis patients: Causes and consequences. Am. J. Kidney Dis. 2003, 42, 864–881. [Google Scholar] [CrossRef] [Green Version]
- Nathan, C. Nitric oxide as a secretory product of mammalian cells. FASEB J. 1992, 6, 3051–3064. [Google Scholar] [CrossRef]
- Moncada, S. Nitric oxide: Discovery and impact on clinical medicine. J. R. Soc. Med. 1999, 92, 164–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jong, M.D.; Simmons, C.P.; Thanh, T.T.; Hien, V.M.; Smith, G.J.; Chau, T.N.; Hoang, D.M.; Chau, N.V.; Khanh, T.H.; Dong, V.C.; et al. Fatal outcome of human influenza A (H5N1) is associated with high viral load and hypercytokinemia. Nat. Med. 2006, 12, 1203–1207. [Google Scholar] [CrossRef] [PubMed]
- Laskin, D.L.; Pendino, K.J. Macrophages and inflammatory mediators in tissueinjury. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 655–677. [Google Scholar] [CrossRef] [PubMed]
- Korns, D.R.; Frasch, S.C.; Fernandez-Boyanapalli, R.; Henson, P.M.; Bratton, D.L. Modulation of Macrophage Efferocytosis in Inflammation. Front. Immunol. 2011, 2, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Kroncke, K.D.; Fehsel, K.; Kolb-Bachofen, V. Inducible nitric oxide synthase inhuman diseases. Clin. Exp. Immunol. 1998, 113, 147–156. [Google Scholar] [CrossRef]
- Turini, M.E.; DuBois, R.N. Cyclooxygenase-2: A Therapeutic Target. Annu. Rev. Med. 2002, 53, 35–57. [Google Scholar] [CrossRef]
- Wang, K.; Holtz, K.M.; Anderson, K.; Chubet, R.; Mahmoud, W.; Cox, M.M. Expression and purification of an influenza hemagglutinin—one step closer to a recombinant protein-based influenza vaccine. Vaccine 2006, 24, 2176–2185. [Google Scholar] [CrossRef]
- Marzia Sichetti, M.S.; Stefania, D.; Marco, M.S.; Pagiotti, B.S.; Giovanna, T.; Donatella, P. Anti-inflflammatory effect of multistrain probiotic formulation (L. rhamnosus, B. lactis, and B. longum). Nutrition 2018, 53, 95–102. [Google Scholar] [CrossRef]
- Kawahara, M.; Nemoto, M.; Nakata, T.; Kondo, S.; Takahashi, H.; Kimura, B.; Takashi, K. Anti-inflflammatory properties of fermented soy milk with Lactococcus lactis subsp. lactis S-SU2 in murine macrophage RAW264.7 cells and DSS-induced IBD model mice. Int. Immunopharmacol. 2015, 26, 295–303. [Google Scholar] [CrossRef]
- Jang, S.E.; Hyam, S.R.; Han, M.J.; Kim, S.Y.; Lee, B.G.; Kim, D.H. Lactobacillus brevis G-101 ameliorates colitis in mice by inhibiting NF-κB, MAPK, and AKT pathways and by polarizing M1 macrophages to M2-like macrophages. J. Appl. Microbiol. 2013, 115, 888–896. [Google Scholar] [CrossRef]
- Li, P.; Ye, J.; Zeng, S.; Yang, C. Florfenicol alleviated lipopolysaccharide (LPS)-induced inflammatory responses in Ctenopharyngodon idella through inhibiting Toll/NF-κB signaling pathways. Fish Shellfish Immunol. 2019, 45, 346–356. [Google Scholar] [CrossRef]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Kanwal, Z.; Wiegertjes, G.; Veneman, W.J.; Meijer, A.H.; Spaink, H.P. Comparative studies of Toll-like receptor signalling using zebrafish. Dev. Comp. Immunol. 2014, 46, 35–52. [Google Scholar] [CrossRef]
- Hua, L.; Jeong-Hyun, Y.; Hyo-Jun, W.; Hyeon-Seon, J.; HeongJoo, Y.; Hun, P.K.; Ho-Yong, P.; Tae-Sook, J. Isotrifoliol inhibits pro-inflammatory mediators by suppression of TLR/NF-κB and TLR/MAPK signaling in LPS-induced RAW264.7 cells. Int. Immunopharmacol. 2017, 45, 110–119. [Google Scholar]
- Li, M.-Y.; Sun, L.; Niu, X.-T.; Chen, X.-M.; Tian, J.-X.; Kong, Y.-D.; Wang, G. Astaxanthin protects lipopolysaccharide-induced inflammatory response in Channa argus through inhibiting NF-κB and MAPKs signaling pathways. Fish Shellfish Immunol. 2019, 86, 280–286. [Google Scholar] [CrossRef]
- Kagan, J.C.; Medzhitov, R. Phosphoinositide-Mediated Adaptor Recruitment Controls Toll-like Receptor Signaling. Cell 2006, 125, 943–955. [Google Scholar] [CrossRef] [Green Version]
- Kawagoe, T.; Sato, S.; Matsushita, K.; Kato, H.; Matsui, K.; Kumagai, Y.; Saitoh, T.; Kawai, T.; Takeuchi, O.; Akira, S. Sequential control of Toll-like receptor–dependent responses by IRAK1 and IRAK2. Nat. Immunol. 2008, 9, 684–691. [Google Scholar] [CrossRef]
- Sanghera, J.S.; Weinstein, S.L.; Aluwalia, M.; Girn, J.; Pelech, S.L. Activation of multiple proline-directed kinases by bacterial lipopolysaccharide in murine macrophages. J. Immunol. 1996, 156, 4457–4465. [Google Scholar]
- Swantek, J.L.; Cobb, M.H.; Geppert, T.D. Jun N-terminal kinase/stress-activated protein kinase (JNK/SAPK) is required for lipopolysaccharide stimulation of tumor necrosis factor alpha (TNF-α) translation: Glucocorticoids inhibit TNF-α translation by blocking JNK/SAPK. Mol. Cell. Biol. 1997, 17, 6274–6282. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Adachi, O.; Ogawa, T.; Takeda, K.; Akira, S. Unresponsiveness of MyD88-Deficient Mice to Endotoxin. Immunity 1999, 11, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Wesche, H.; Henzel, W.; Shillinglaw, W.; Li, S.; Cao, Z. MyD88: An Adapter That Recruits IRAK to the IL-1 Receptor Complex. Immunity 1997, 7, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Takeda, K.; Akira, S. TLR signaling pathways. Semin. Immunol. 2004, 16, 3–9. [Google Scholar] [CrossRef]
- Sepulcre, M.P.; Alcaraz-Perez, F.; Lopez-Munoz, A.; Cayuela, M.L.; Mulero, V. Evolution of lipopolysaccharide (LPS) recognition and signaling: Fish TLR4 does not recognize LPS and negatively regulates NF-kappaB activation. J. Immunol. 2009, 182, 1836–1845. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.W.; Cai, L.; Qi, Z.H.; Wang, C.; Liu, Y.; Wang, W.N. Effects of a recombinant complement component C3b functional fragment α 2 MR (α 2-macroglobulin receptor) additive on the immune response of juvenile orange-spotted grouper (Epinepheluscoioides) after the exposure to cold shock challenge. Fish Shellfish Immunol. 2015, 45, 346–356. [Google Scholar] [CrossRef]
- Jangiam, W.; Tungjai, M.; Rithidech, K.N. Induction of chronic oxidative stress, chronic inflammation and aberrant patterns of DNA methylation in the liver of titanium-exposed CBA/CaJ mice. Int. J. Radiat. Biol. 2015, 91, 389–398. [Google Scholar] [CrossRef]
- Yu, H.; Valerio, M.; Bielawski, J. Fenretinide inhibited de novo ceramide synthesis and proinflammatory cytokines induced by Aggregatibacteractinomycetemcomitans. J. Lipid Res. 2013, 54, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.Z.; Tang, X.F.; Zhao, X.H.; Zhang, M.H.; Zhang, W.J.; Hou, S.H.; Yuan, W.F.; Zhang, H.F.; Shi, L.J.; Jia, H.; et al. Tylvalosin exhibits anti-inflammatory property and attenuates acute lung injury in different models possibly through suppression of NF-kappa B activation. Biochem. Pharm. 2014, 90, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.L.; Xu, Y.S.; Wu, W.T.; Li, W.Y.; Sun, Y.R.; Yang, J.; Zhu, Y.; Zhang, C. Tetrandrine Suppresses Lipopolysaccharide-Induced Microglial Activation by Inhibiting NF-kappa B and ERK Signaling Pathways in BV2 Cells. PLoS ONE 2014, 9, e102522. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.R.; Chen, X.; Chen, C.F. Distribution of Intraperitoneally Administered Cytophagacolum naris Lipopolysaccharidein Gibel Carp, Carassius auratus gibelio. J. Huazhong Agric. Univ. 2003, 22, 243–246. [Google Scholar]
- Chen, X.Y.; Hu, C.J. Tissue distribution of vibio alginolyticus lipopolysaccharide in malabar grouper, epinephelus malabaricus, abudefduf vaigiensis. J. Trop. Oceanogr. 2002, 21, 30–35. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| NF-kB | GAAGAAGGATGTGGGAGATG | TGTTGTCGTAGATGGGCTGAG |
| TLR1 | TGTGCCACCGTTTGGATA | TTCAGGGCGAACTTGTCG |
| TLR2 | CTATCAAGTGCTCCTCAAA | CCTCACCCATGTAGTATGT |
| TLR4 | CCACCTATTCATCTTTGCCT | GTCTTCCCCTCTTCCACATC |
| TLR7 | GAGCATACAGTTGAGTAAACGCAC | TCTCCAAGAATATCAGGACGATAA |
| TLR8 | TCACATCGCTTCCAGGTCTC | ACGGTGAAATAATGGGGGTT |
| NF-κB p65 | AACCAAGAACCAGCCATACAAG | AACCAAGAACCAGCCATACAAG |
| IκB | TCTTGCCATTATTCACGAGG | TGTTACCACAGTCATCCACCA |
| MyD88 | GACTGTCGCCGAAATGA | TGCCTTCTCGCTCCTGT |
| IRAK4 | CTCCACACTGAGAGCTTTATC | ATGTGCAGCTGTGTGTATCT |
| TRAF6 | TCACTCACTGTCAGATGTC | TGTTGGCTCTTGTGTTCA |
| TAK1 | AGACAGGACAGACACCAAT | CATCTTACAGTGCTGCTCAA |
| GAPDH | AGGGGCTCAGTATGTTGTGG | CTCTCTTGGCACCACCCTTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Xu, Y.; Cao, Y.; Ding, Z. Polypeptides Isolated from Lactococcus lactis Alleviates Lipopolysaccharide (LPS)-Induced Inflammation in Ctenopharyngodon idella. Int. J. Mol. Sci. 2022, 23, 6733. https://doi.org/10.3390/ijms23126733
Li P, Xu Y, Cao Y, Ding Z. Polypeptides Isolated from Lactococcus lactis Alleviates Lipopolysaccharide (LPS)-Induced Inflammation in Ctenopharyngodon idella. International Journal of Molecular Sciences. 2022; 23(12):6733. https://doi.org/10.3390/ijms23126733
Chicago/Turabian StyleLi, Pei, Youqing Xu, Yupo Cao, and Zhaokun Ding. 2022. "Polypeptides Isolated from Lactococcus lactis Alleviates Lipopolysaccharide (LPS)-Induced Inflammation in Ctenopharyngodon idella" International Journal of Molecular Sciences 23, no. 12: 6733. https://doi.org/10.3390/ijms23126733
APA StyleLi, P., Xu, Y., Cao, Y., & Ding, Z. (2022). Polypeptides Isolated from Lactococcus lactis Alleviates Lipopolysaccharide (LPS)-Induced Inflammation in Ctenopharyngodon idella. International Journal of Molecular Sciences, 23(12), 6733. https://doi.org/10.3390/ijms23126733