
Discovery of a New Microbial Origin Cold-Active Neopullulanase Capable for Effective Conversion of Pullulan to Panose


Abstract
:1. Introduction
2. Results
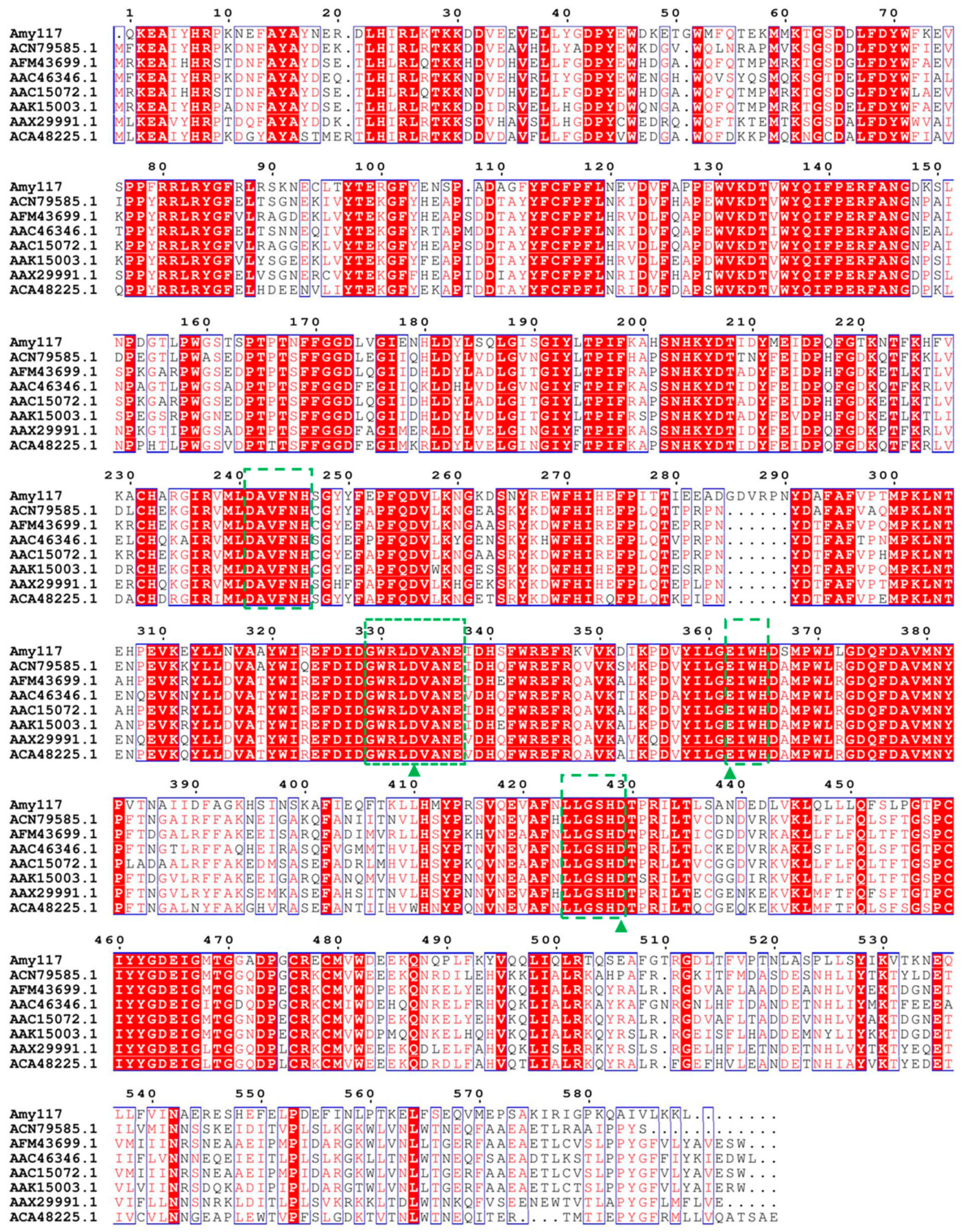
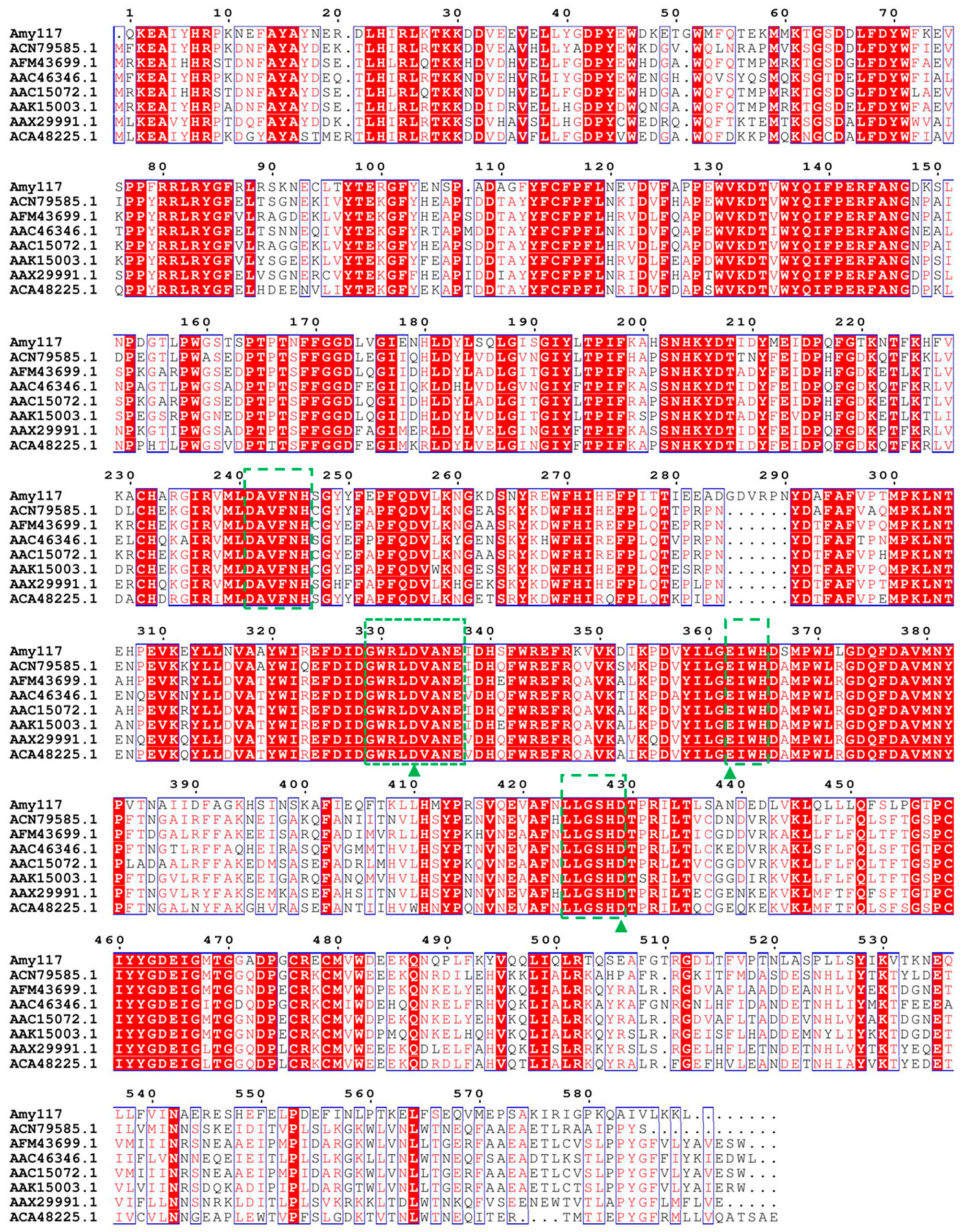
2.1. Sequence Analysis, Expression, and Purification of Amy117
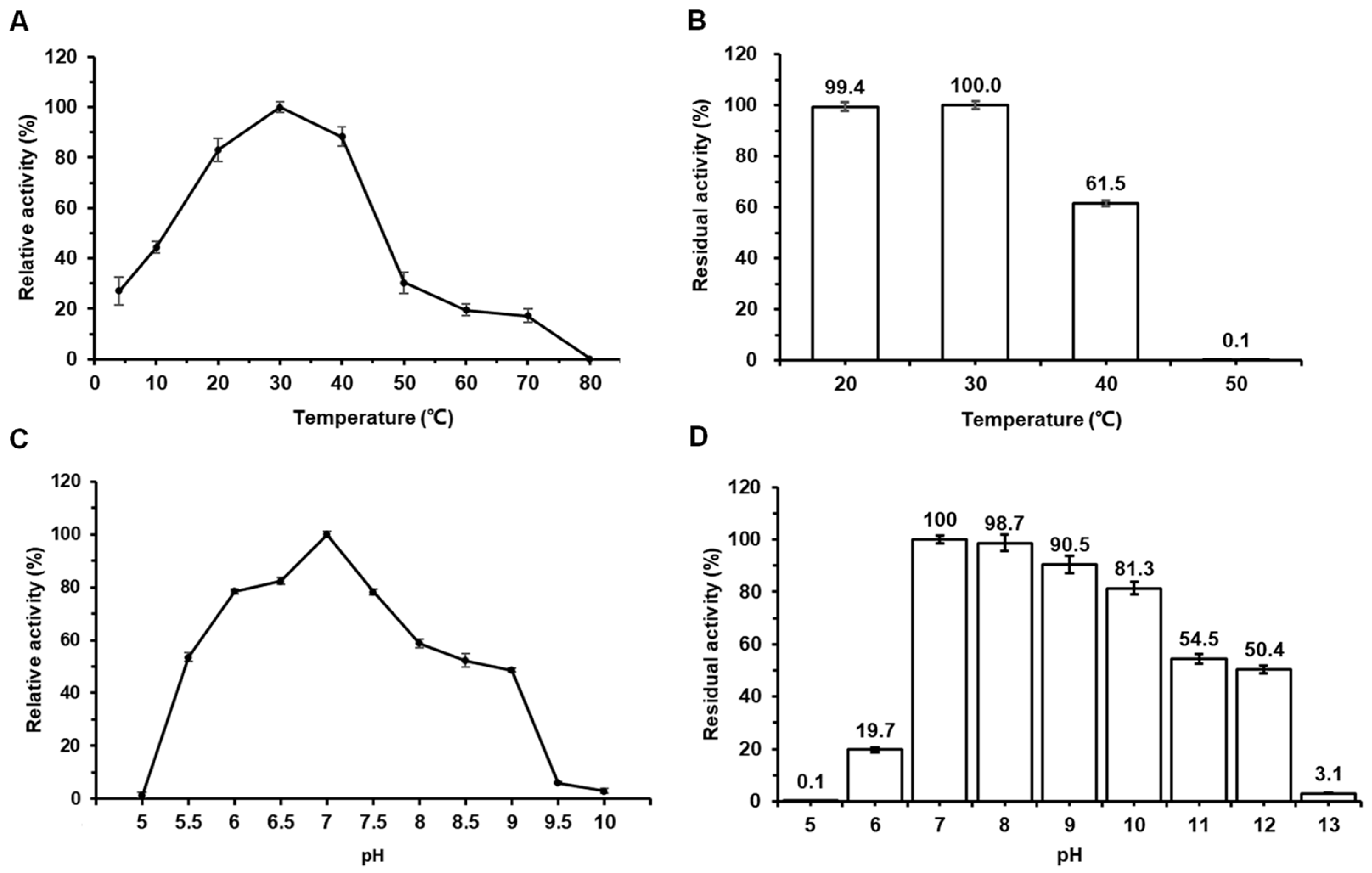
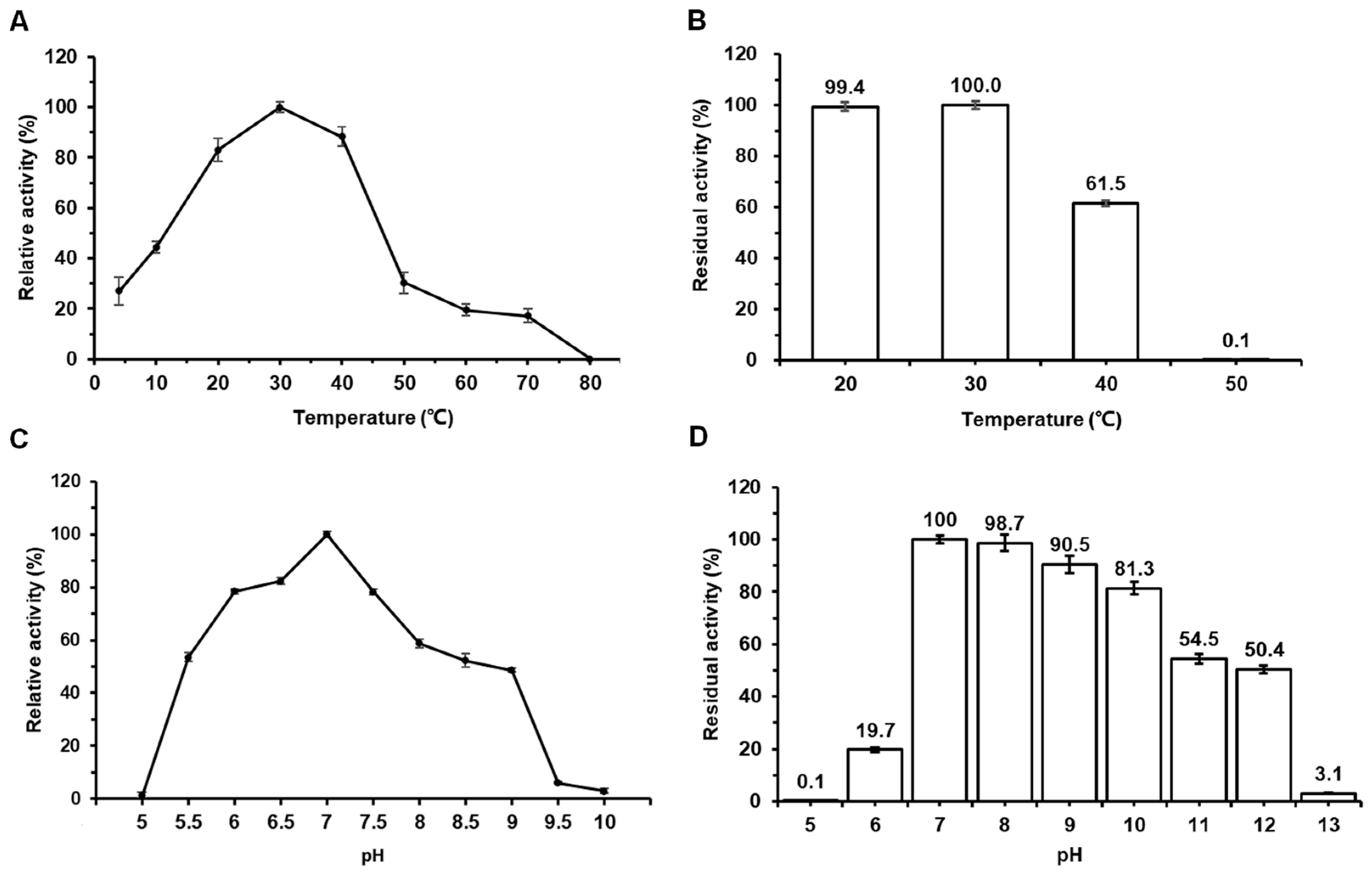
2.2. Effects of Temperature and pH on the Activity and Stability of Amy117
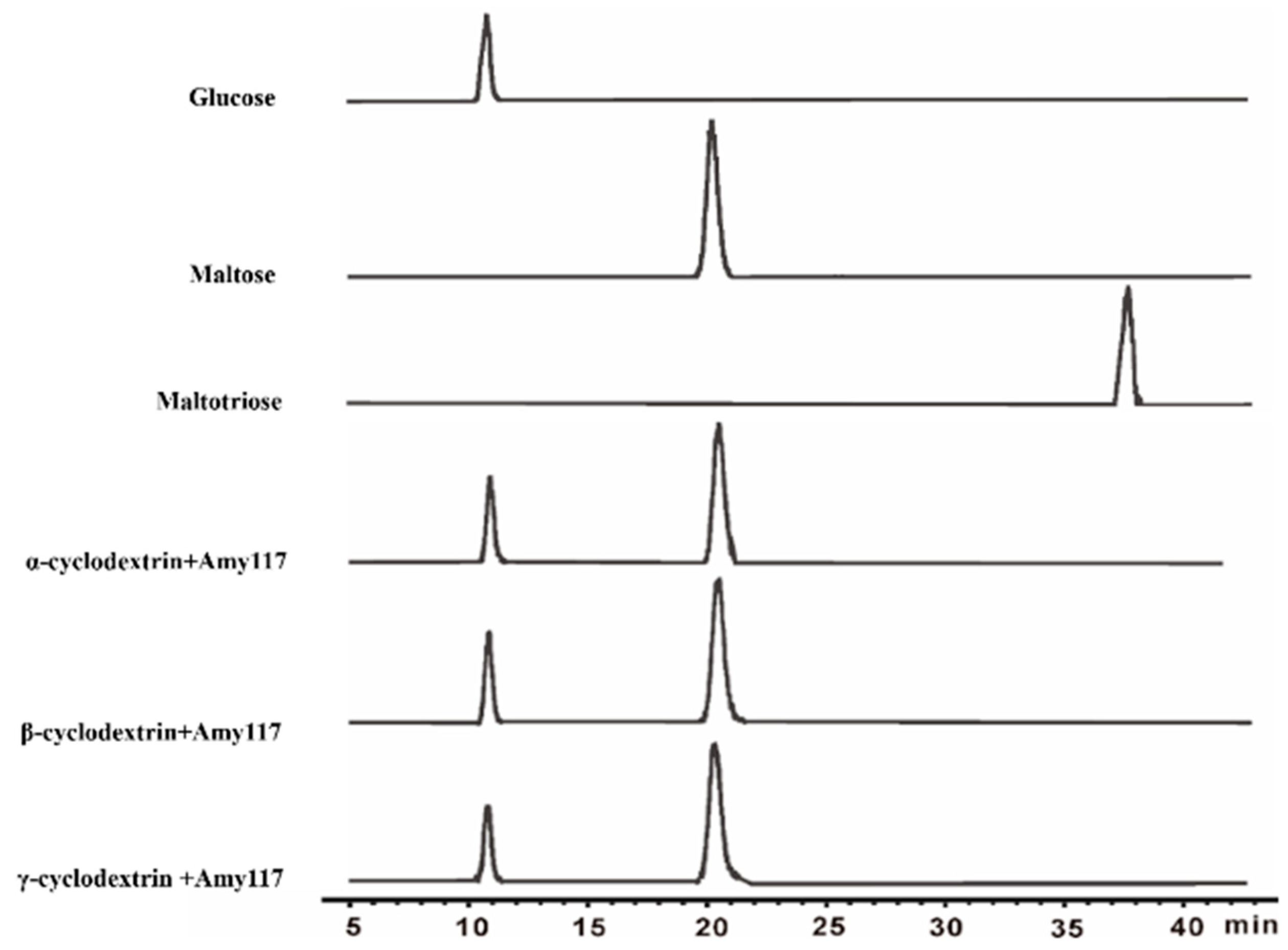
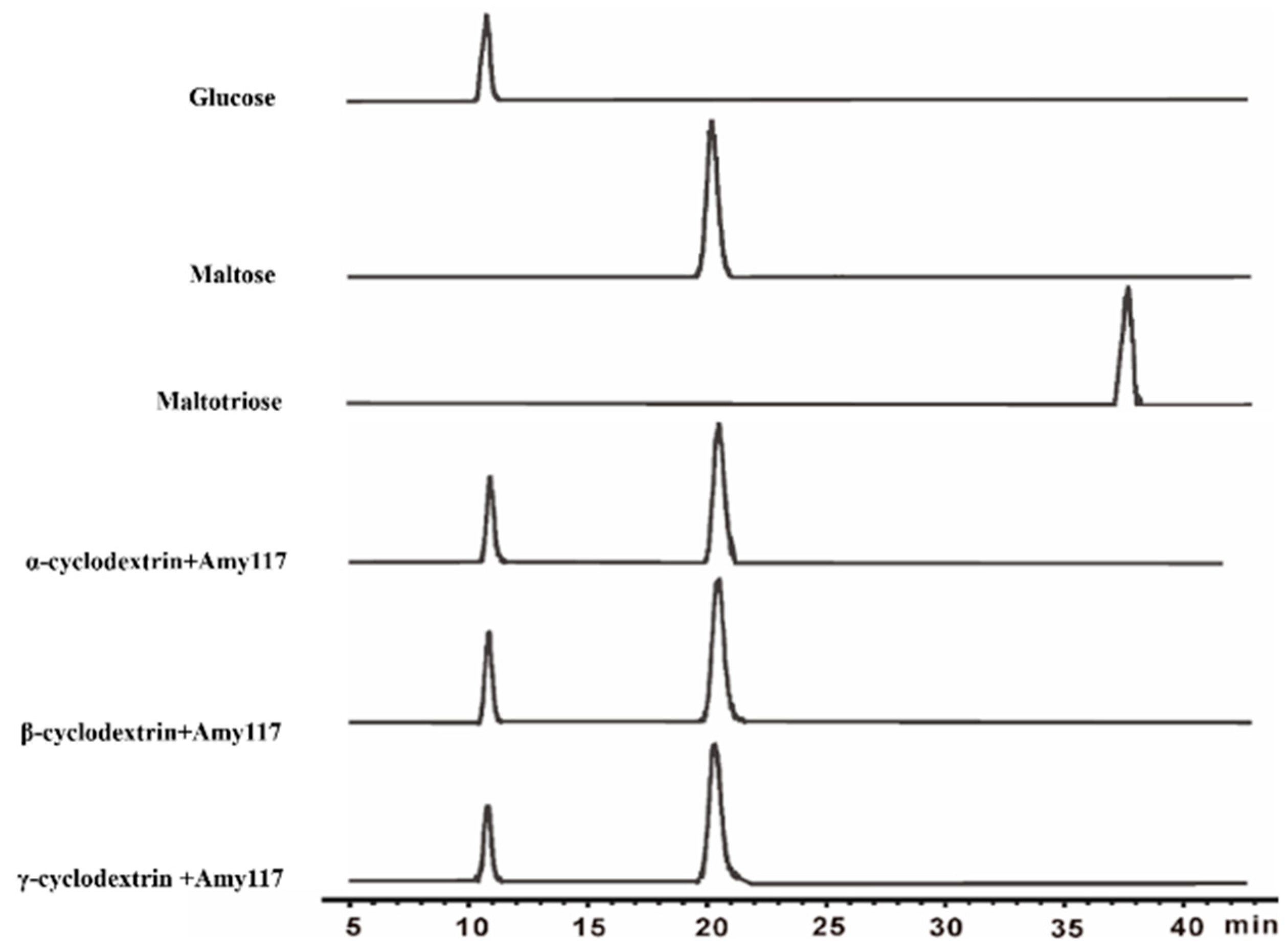
2.3. Substrate Specificity of Amy117
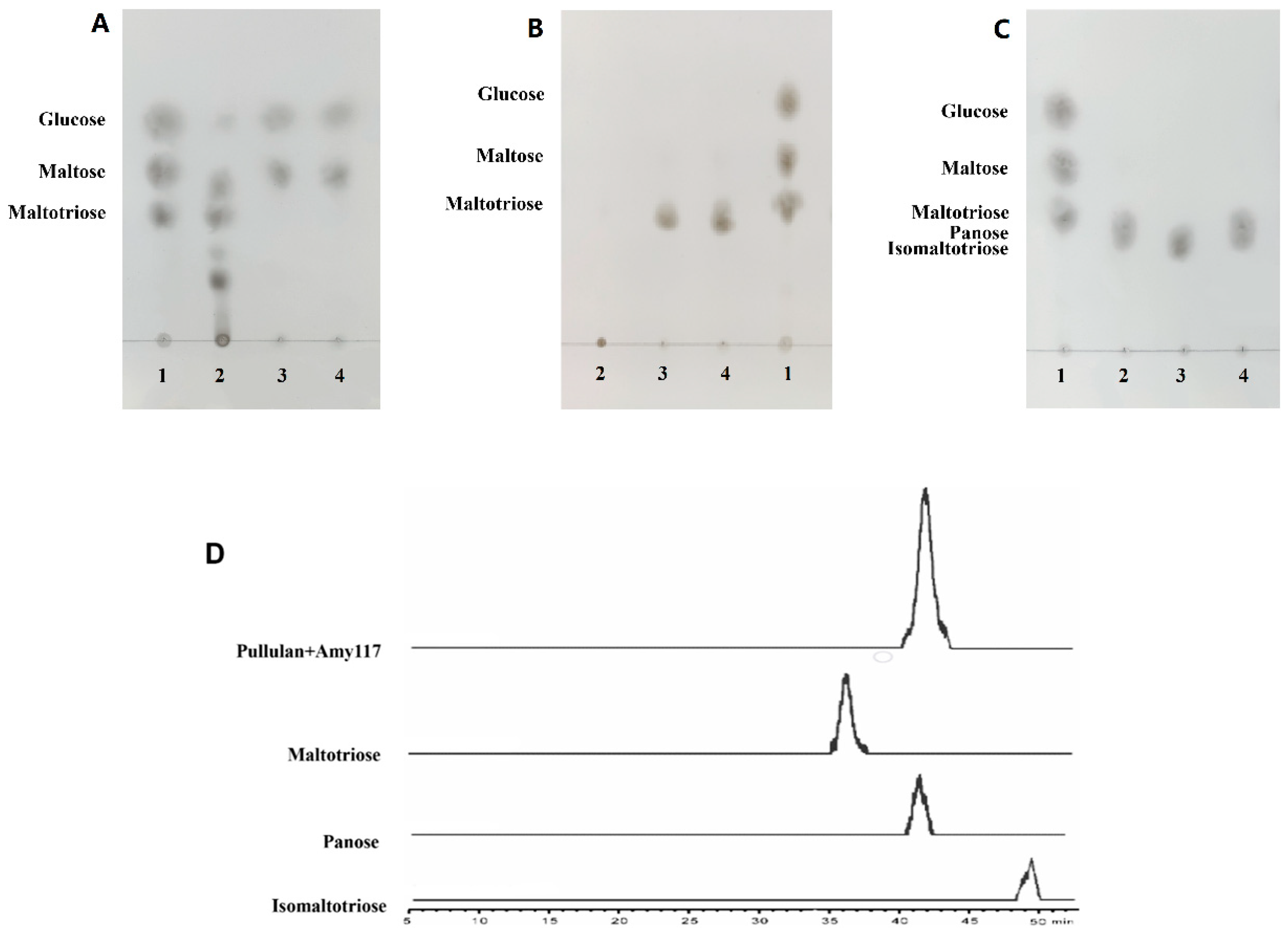
2.4. Action Modes of Amy117 on Various Substrates
2.5. Production of Panose by Amy117
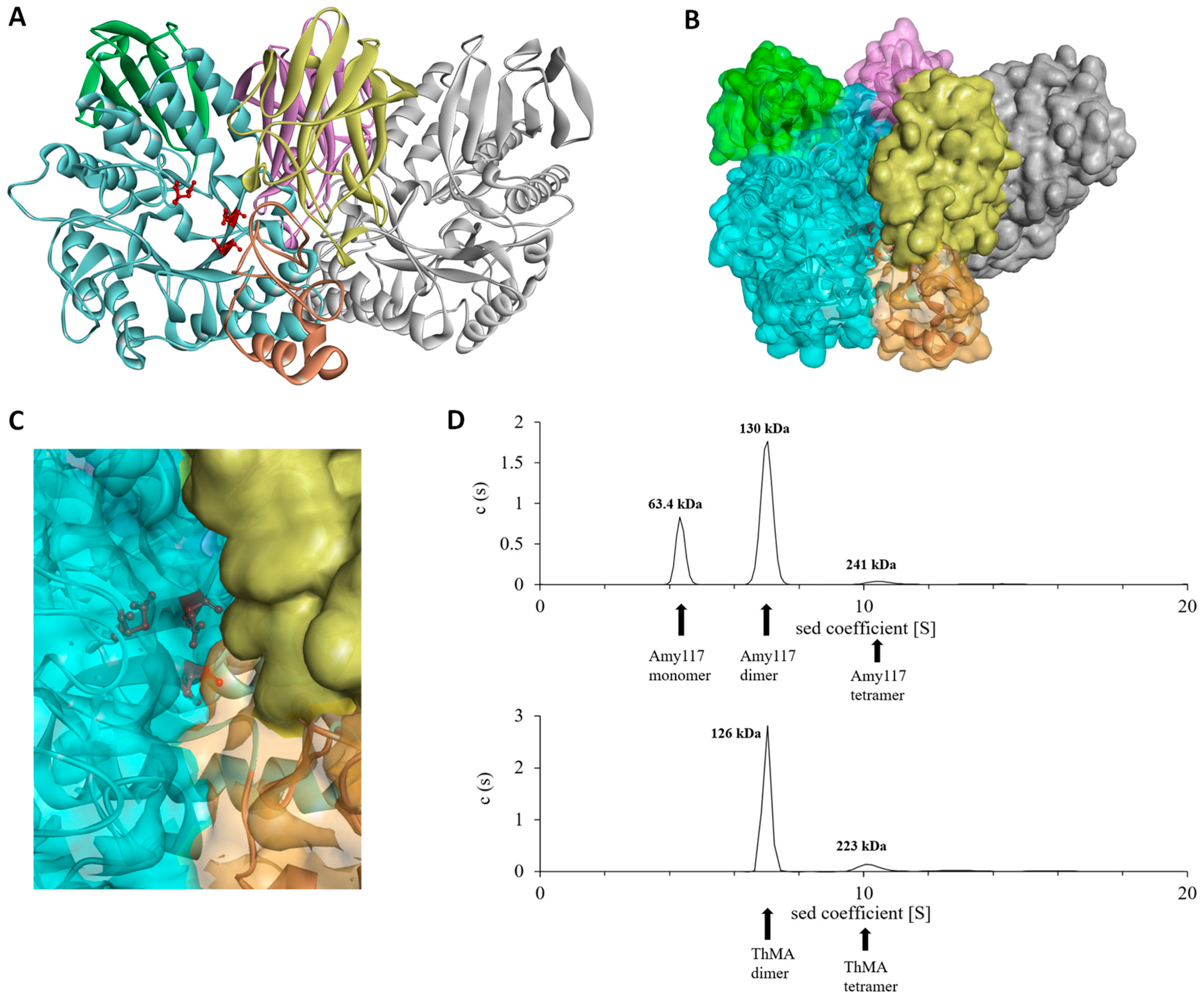
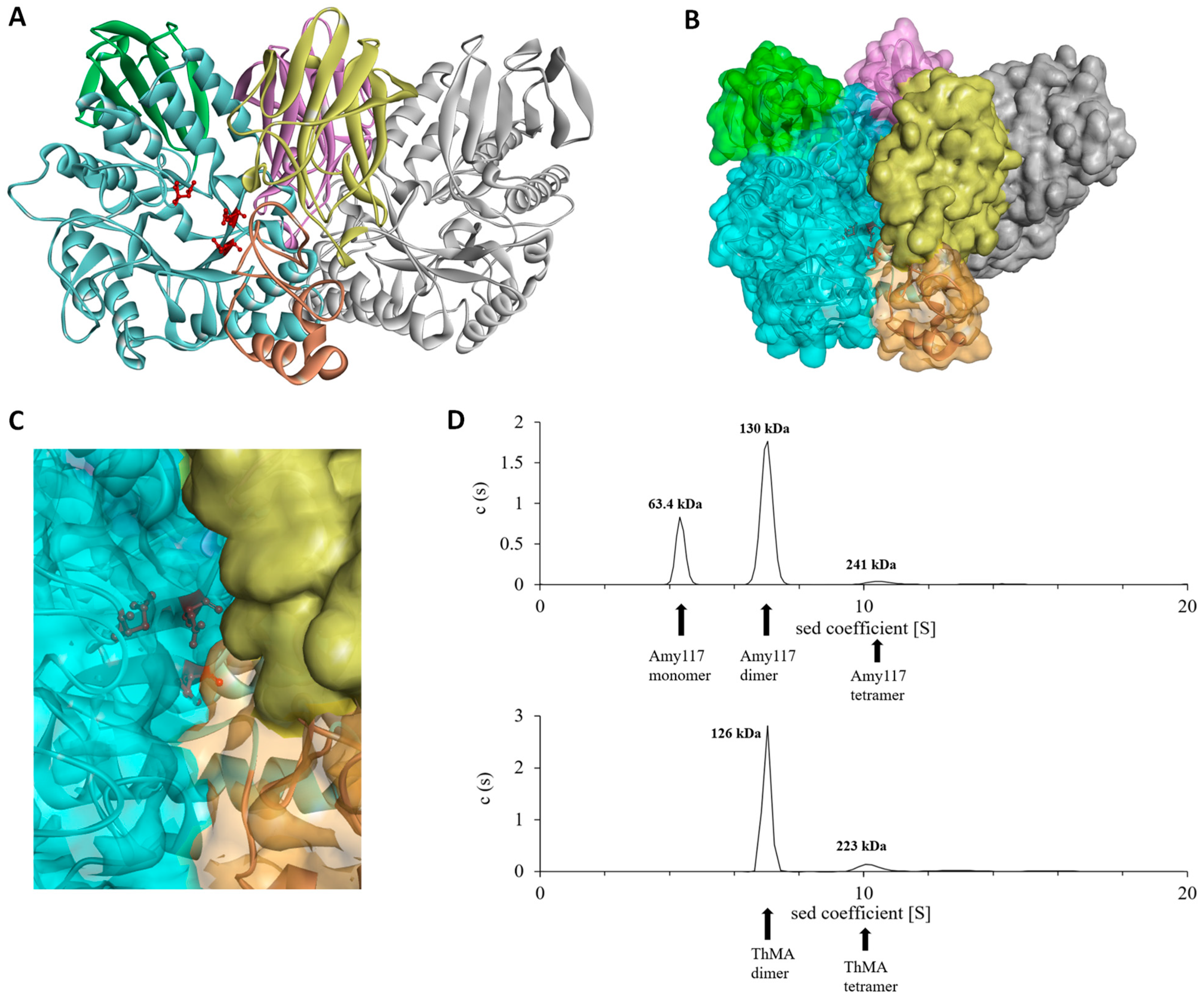
2.6. Structure Modeling of Amy117
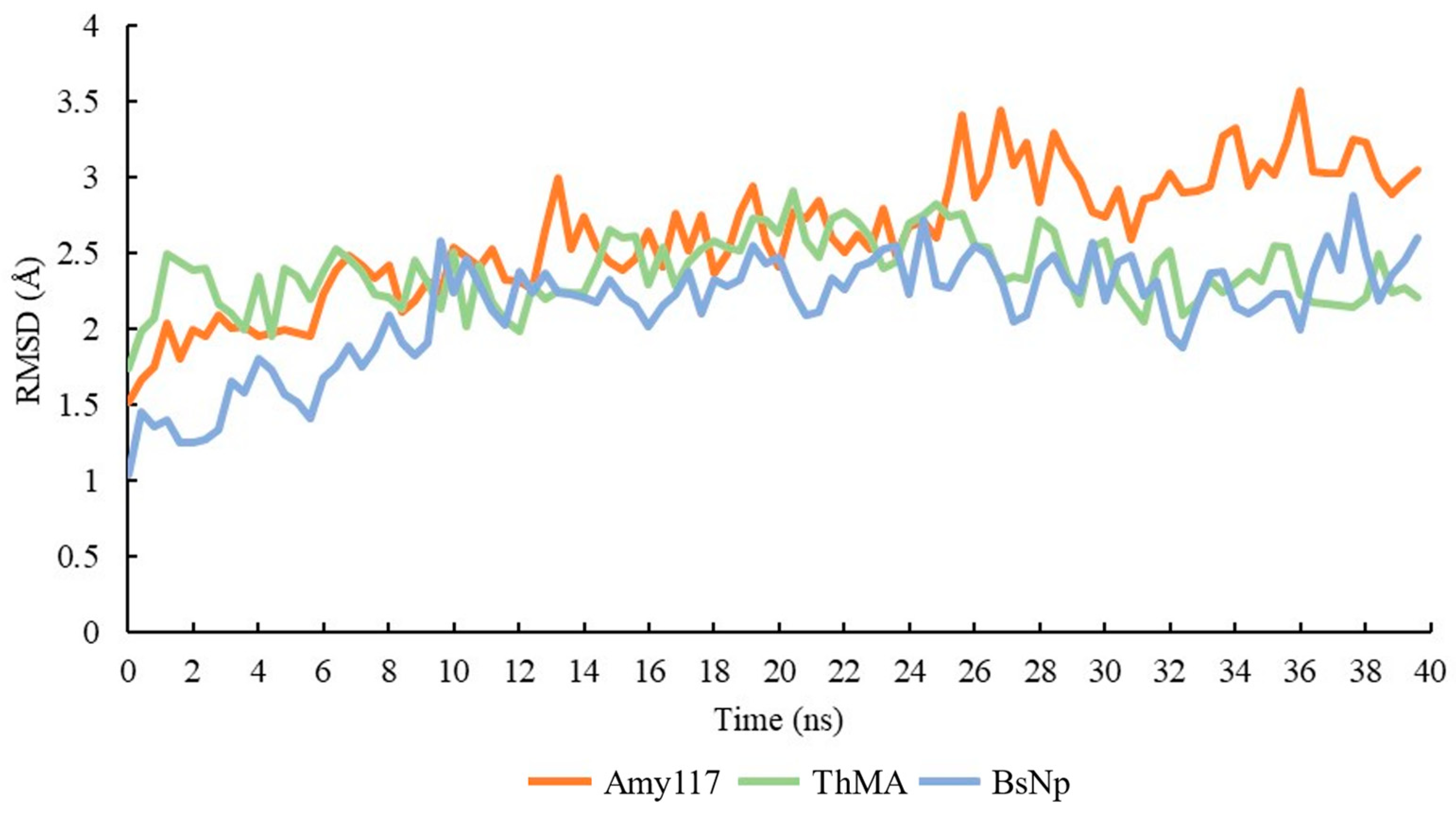
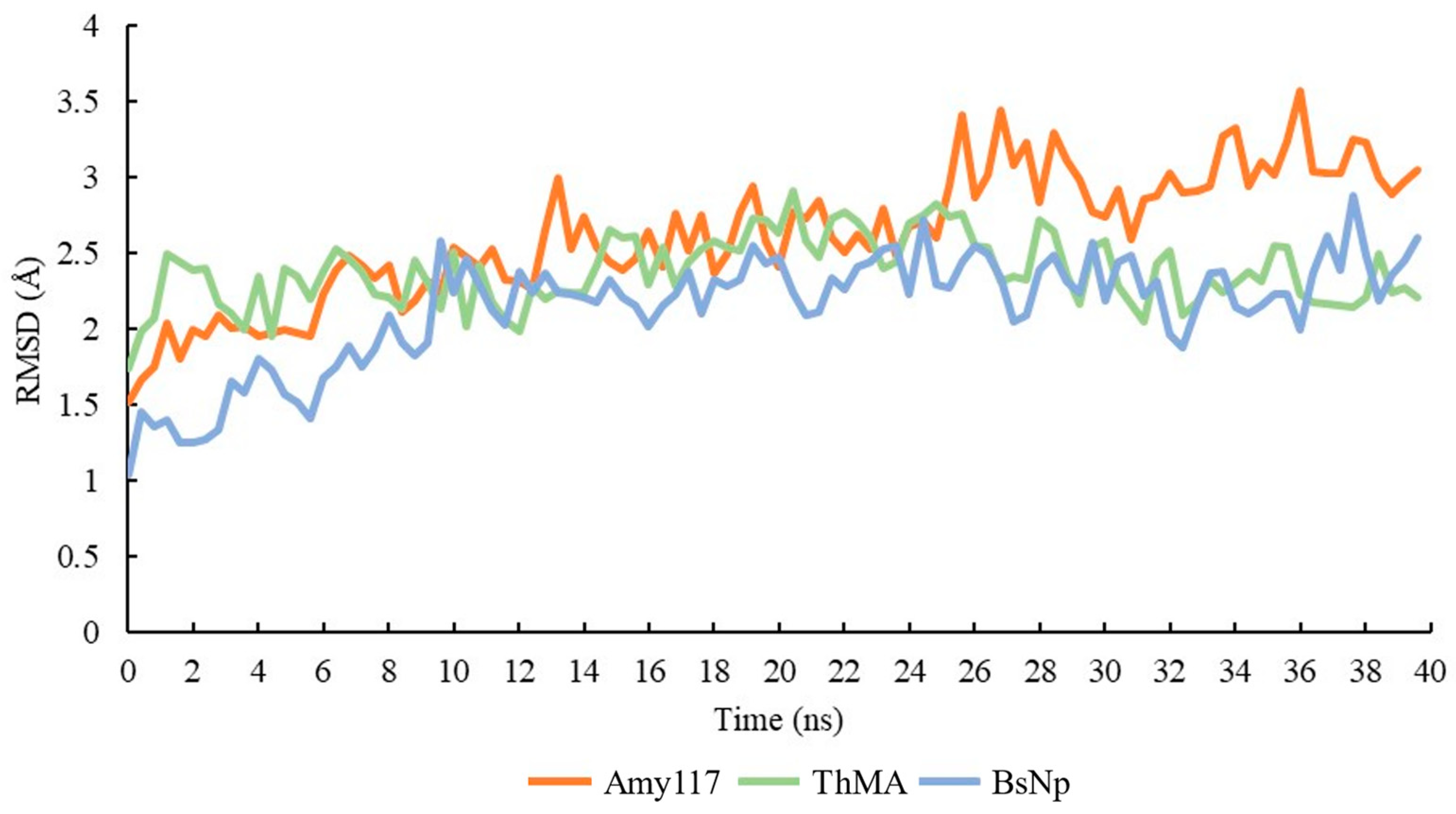
2.7. Molecular Dynamic (MD) of Amy117 and Its Homologues
3. Discussion
4. Materials and Methods
4.1. Plasmids, Strains, Chemicals, and Media
4.2. Cloning and Sequence Analysis of an α-Glycosidase or Putative NPase Gene
4.3. Expression and Purification of the Recombinant α-Glycosidase or Putative NPase in E. coli
4.4. Temperature and pH Preference, Substrate Specificity, and Catalytic Constants of the α-Glycosidase or Putative NPase Amy117
4.4.1. Temperature Preference of Amy117
4.4.2. Temperature Preference of Amy117
4.4.3. Substrate Specificity of Amy117
4.4.4. Catalytic Constants of Amy117
4.5. Analysis of Composition the Enzymatic Hydrolysis Products by HPLC and TLC
4.6. Panose Production by the Purified Amy117
4.7. Structural Simulation and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, M.; Jang, J.K.; Park, Y.S. Production optimization, structural analysis, and prebiotic- and anti-inflammatory effects of gluco-oligosaccharides produced by leuconostoc lactis sbc001. Microorganisms 2021, 9, 200. [Google Scholar] [CrossRef] [PubMed]
- Kuriki, T.; Takata, H.; Okada, S.; Imanaka, T. Analysis of active center of Bacillus stearothermophilus neopullulanase. J. Bacteriol. 1992, 173, 6147–6152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, F.A.; Rodrigues, S. Evaluation of enzymatic reactors for large-scale panose production. Appl. Biochem. Biotechnol. 2007, 142, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Makelainen, H.; Hasselwander, O.; Rautonen, N.; Ouwehand, A.C. Panose, a new prebiotic candidate. Appl. Biochem. Biotechnol. 2009, 49, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Nisha, M.; Satyanarayana, T. Characteristics, protein engineering and applications of microbial thermostable pullulanases and pullulan hydrolases. Appl. Microbiol. Biotechnol. 2016, 100, 5661–5679. [Google Scholar] [CrossRef] [PubMed]
- Hii, S.L.; Tan, J.S.; Ling, T.C.; Ariff, A.B. Pullulanase: Role in starch hydrolysis and potential industrial applications. Enzym. Res. 2012, 2012, 921362. [Google Scholar] [CrossRef] [Green Version]
- Park, K.H.; Kim, T.J.; Cheong, T.K.; Kim, J.W.; Oh, B.H.; Svensson, B. Structure, specificity and function of cyclomaltodextrinase, a multispecific enzyme of the α-amylase family. Biochim. Biophys. Acta 2000, 1478, 165–185. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, M.S.; Cho, H.S.; Kim, J.I.; Kim, T.J.; Choi, J.H.; Park, C.; Lee, H.S.; Oh, B.H.; Park, K.H. Cyclomaltodextrinase, neopullulanase, and maltogenic amylase are nearly indistinguishable from each other. J. Biol. Chem. 2002, 277, 21891–21897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domań-Pytka, M.; Bardowski, J. Pullulan degrading enzymes of bacterial origin. Crit. Rev. Microbiol. 2004, 30, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Roozbehi, S.; Dadashzadeh, S.; Sajedi, R.H. An enzyme-mediated controlled release system for curcumin based on cyclodextrin/cyclodextrin degrading enzyme. Enzym. Microb. Technol. 2020, 144, 109727. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.; Nilsson, C.; Svensson, D.; Holst, O.; Gorton, L.; Karlsson, E.N. Monomeric and dimeric cyclomaltodextrinases reveal different modes of substrate degradation. Biologia 2005, 60, 79–87. [Google Scholar]
- Cheong, K.A.; Kim, T.J.; Yoon, J.W.; Park, C.S.; Lee, T.S.; Kim, J.W. Catalytic activities of intracellular dimeric neopullulanase on cyclodextrin, acarbose and maltose. Biotechnol. Appl. Biochem. 2011, 35, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.N.; Labes, A.; Turner, P.; Fridjonsson, O.H.; Wennerberg, C.; Pozzo, T.; Hreggvidson, G.O.; Kristjansson, J.K.; Schönheit, P. Differences and similarities in enzymes from the neopullulanase subfamily isolated from thermophilic species. Biologia 2008, 63, 1006–1014. [Google Scholar] [CrossRef]
- Labes, A.; Karlsson, E.N.; Fridjonsson, O.H.; Turner, P.; Hreggvidson, G.O.; Kristjansson, J.K.; Holst, O.; Schönheit, P. Novel members of glycoside hydrolase family 13 derived from environmental DNA. Appl. Environ. Microbiol. 2008, 74, 1914–1921. [Google Scholar] [CrossRef] [Green Version]
- Balolong, M.P.; Chae, J.P.; Kang, D.K. Expression and characterisation of neopullulanase from Lactobacillus mucosae. Biotechnol. Lett. 2016, 38, 1753–1760. [Google Scholar] [CrossRef]
- Zhou, J.; Li, Z.; Zhang, H.; Wu, J.; Ye, X.; Dong, W.; Jiang, M.; Huang, Y.; Cui, Z. Novel maltogenic amylase CoMA from Corallococcus sp. EGB catalyzes the conversion of maltooligosaccharides and soluble starch to maltose. Appl. Environ. Microbiol. 2018, 84, e00152-18. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.H.; An, Y.K.; Son, S.Y.; Jeong, S.Y.; Seo, D.H.; Kim, M.K.; Park, C.S. Characterization of a novel extracellular α-amylase from Ruminococcus bromii ATCC 27255 with neopullulanase-like activity. Int. J. Biol. Macromol. 2019, 130, 605–614. [Google Scholar] [CrossRef]
- Janeček, Š. How many conserved sequence regions are there in the α-amylase family. Biologia 2002, 57, 29–41. [Google Scholar]
- Kolcuoğlu, Y.; Colak, A.; Faiz, O.; Belduz, A.O. Cloning, expression and characterization of highly thermo- and pH-stable maltogenic amylase from a thermophilic bacterium Geobacillus caldoxylosilyticus TK4. Process Biochem. 2010, 45, 821–828. [Google Scholar] [CrossRef]
- Deepika, M.; Tulasi, S.; Uversky, V.N. Dimerization mediates thermo-adaptation, substrate affinity and transglycosylation in a highly thermostable maltogenic amylase of Geobacillus thermoleovorans. PLoS ONE 2013, 8, e73612. [Google Scholar]
- Cha, H.J.; Yoon, H.G.; Kim, Y.W.; Lee, H.S.; Kim, J.W.; Kweon, K.S.; Oh, B.H.; Park, K.H. Molecular and enzymatic characterization of a maltogenic amylase that hydrolyzes and transglycosylates acarbose. Eur. J. Biochem. 1998, 253, 251–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.S.; Cha, S.S.; Kim, H.J.; Kim, T.J.; Ha, N.C.; Oh, S.T.; Cho, H.S.; Cho, M.J.; Kim, M.J.; Lee, H.S.; et al. Crystal structure of a maltogenic amylase provides insights into a catalytic versatility. J. Biol. Chem. 1999, 274, 26279–26286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anna, C.; Katarzyna, K.; Katarzyna, N.; Marek, K.; Małgorzata, G.; Danuta, K. Effect of pullulan on physicochemical, microbiological, and sensory quality of yogurts. Curr. Pharm. Biotechnol. 2019, 20, 489–496. [Google Scholar]
- Kuchtová, A.; Janeček, Š. Domain evolution in enzymes of the neopullulanase subfamily. Microbiology 2016, 162, 2099–2115. [Google Scholar] [CrossRef] [PubMed]
- Machovic, M.; Janecek, Š. Starch-binding domains in the postgenome era. Cell. Mol. Life Sci. 2006, 63, 2710–2724. [Google Scholar] [CrossRef] [PubMed]
- Hondoh, H.; Kuriki, T.; Matsuura, Y. Three-dimensional structure and substrate binding of Bacillus stearothermophilus neopullulanase. J. Mol. Biol. 2003, 326, 177–188. [Google Scholar] [CrossRef]
- Krieger, E.; Vriend, G. New ways to boost molecular dynamics simulations. J. Comput. Chem. 2015, 36, 996–1007. [Google Scholar] [CrossRef]
- Pikkemaat, M.G.; Linssen, A.B.; Berendsen, H.J.; Janssen, D.B. Molecular dynamics simulations as a tool for improving protein stability. Protein Eng. 2002, 15, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, M.; Tonozuka, T.; Uechi, A.; Ohtaki, A.; Ichikawa, K.; Kamitori, S.; Nishikawa, A.; Sakano, Y. The crystal structure of Thermoactinomyces vulgaris R-47 α-amylase II (TVA II) complexed with transglycosylated product. Eur. J. Biochem. 2004, 271, 2530–2538. [Google Scholar] [CrossRef]
- Matzke, J.; Herrmann, A.; Schneider, E.; Bakker, E.P. Gene cloning, nucleotide sequence and biochemical properties of a cytoplasmic cyclomaltodextrinase (neopullulanase) from Alicyclobacillus acidocaldarius, reclassification of a group of enzymes. FEMS Microbiol. Lett. 2000, 183, 55–61. [Google Scholar] [CrossRef]
- Russell, N.J. Toward a molecular understanding of cold activity of enzymes from psychrophiles. Extremophiles 2000, 4, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Nathalia, V.; Rafael, A.D.; Rodrigo, V.A.; Cristiane, D.A.; Debor, F.; David, R. Structural mechanism for the temperature-dependent activation of the hyperthermophilic Pf2001 esterase. Structure 2018, 26, 199–208. [Google Scholar]
- Shukla, R.J.; Singh, S.P. Characteristics and thermodynamics of α-amylase from thermophilic actinobacterium Laceyella sacchari TSI-2. Process Biochem. 2015, 50, 2128–2136. [Google Scholar] [CrossRef]
- Jung, J.H.; Seo, D.H.; Holden, J.F.; Park, C.S. Maltose-forming α-amylase from the hyperthermophilic archaeon pyrococcus sp. ST04. Appl. Microbiol. Biotechnol. 2014, 98, 2121–2131. [Google Scholar] [CrossRef] [PubMed]
- Jeon, E.J.; Jung, J.H.; Seo, D.H.; Jung, D.H.; Holden, J.F.; Park, C.S. Bioinformatic and biochemical analysis of a novel maltose-forming α-amylase of the GH57 family in the hyperthermophilic archaeon Thermococcus sp. CL1. Enzym. Microb. Technol. 2014, 60, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, S.; Varadarajan, R. Elucidation of determinants of protein stability through genome sequence analysis. FEBS Lett. 2000, 470, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Pack, S.P.; Yoo, Y.J. Protein thermostability: Structure-based difference of amino acid between thermophilic and mesophilic proteins. J. Biotechnol. 2004, 111, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Tian, C.; Li, A.; Zhang, G.; Ma, Y. Identification and characterization of a novel alkaline α-amylase Amy703 belonging to a new clade from Bacillus pseudofirmus. J. Ind. Microbiol. Biotechnol. 2014, 41, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yin, L.; Hu, H.; Selvaraj, J.N.; Zhou, Y.; Zhang, G. Expression, functional analysis and mutation of a novel neutral zearalenone-degrading enzyme. Int. J. Biol. Macromol. 2018, 118, 1284–1292. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, C.; Rodriguez-Jasso, R.M.; Gomes, N.; Teixeira, J.A.; Belo, I. Adaptation of dinitrosalicylic acid method to microtiter plates. Anal. Methods. 2010, 2, 2046–2048. [Google Scholar] [CrossRef] [Green Version]
- Noble, J.E.; Bailey, M.J. Quantitation of protein. Methods Enzymol. 2009, 463, 73–95. [Google Scholar] [PubMed]
- Nathalie, D.; Mischa, M.; Philippe, J.; Georg, W.; Robert, H.; Claude, G. Hyperthermostabilization of bacillus licheniformis α-amylase and modulation of its stability over a 50 °C temperature range. Protein Eng. 2003, 16, 287–293. [Google Scholar]
- Himmelsbach, M.; Waser, M.; Klampfl, C.W. Thin layer chromatography–spray mass spectrometry: A method for easy identification of synthesis products and UV filters from TLC aluminum foils. Anal. Bioanal. Chem. 2014, 406, 3647–3656. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Land, H.; Humble, M.S. YASARA: A Tool to Obtain Structural Guidance in Biocatalytic investiations. Methods Mol. Biol. 2018, 1685, 43–67. [Google Scholar]
- Pawar, S.S.; Rohane, S.H. Review on Discovery Studio: An important Tool for Molecular Docking. Asian J. Res. Chem. 2021, 14, 86–88. [Google Scholar] [CrossRef]
- Schuck, P.; Gillis, R.B.; Besong, T.M.D.; Almutairi, F.; Adams, G.G.; Rowe, A.J.; Harding, S.E. SEDFIT-MSTAR: Molecular weight and molecular weight distribution analysis of polymers by sedimentation equilibrium in the ultracentrifuge. Analyst 2014, 139, 79–92. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrates 1 | Specific Activity (U/mg) |
|---|---|
| α-cyclodextrin | 851.92 ± 10.60 |
| β-cyclodextrin | 388.49 ± 1.33 |
| γ-cyclodextrin | 94.88 ± 0.34 |
| Pullulan | 62.50 ± 0.11 |
| Soluble starch | 29.31 ± 0.11 |
| Amylopectin from maize | N.D. 2 |
| Amylopectin from potato | N.D. |
| Amylose from potato | N.D. |
| Substrates (Pullulan) | Production of Panose (mg/mL) | Conversion Yield (w/w) |
|---|---|---|
| 10 mg/mL | 10.42 | 104.2% |
| 40 mg/mL | 40.33 | 100.8% |
| 80 mg/mL | 72.74 | 90.9% |
| 100 mg/mL | 72.72 | 72.7% |
| Number of Amino Acids | Arg | Arg/(Arg+Lys) | Val, Ile and Thr | Asn and Gln |
|---|---|---|---|---|
| Amy117 | 4.24% | 0.40 | 4.58% | 8.15% |
| ThMA | 5.95% | 0.53 | 3.23% | 6.12% |
| BsNp | 5.95% | 0.55 | 4.08% | 6.97% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Hu, H.; Zhang, B.; Zheng, Y.; Wu, P.; Lu, Z.; Zhang, G. Discovery of a New Microbial Origin Cold-Active Neopullulanase Capable for Effective Conversion of Pullulan to Panose. Int. J. Mol. Sci. 2022, 23, 6928. https://doi.org/10.3390/ijms23136928
Wang M, Hu H, Zhang B, Zheng Y, Wu P, Lu Z, Zhang G. Discovery of a New Microbial Origin Cold-Active Neopullulanase Capable for Effective Conversion of Pullulan to Panose. International Journal of Molecular Sciences. 2022; 23(13):6928. https://doi.org/10.3390/ijms23136928
Chicago/Turabian StyleWang, Meixing, Huizhen Hu, Buyu Zhang, Yang Zheng, Pan Wu, Zhenghui Lu, and Guimin Zhang. 2022. "Discovery of a New Microbial Origin Cold-Active Neopullulanase Capable for Effective Conversion of Pullulan to Panose" International Journal of Molecular Sciences 23, no. 13: 6928. https://doi.org/10.3390/ijms23136928
APA StyleWang, M., Hu, H., Zhang, B., Zheng, Y., Wu, P., Lu, Z., & Zhang, G. (2022). Discovery of a New Microbial Origin Cold-Active Neopullulanase Capable for Effective Conversion of Pullulan to Panose. International Journal of Molecular Sciences, 23(13), 6928. https://doi.org/10.3390/ijms23136928