
Centipede Venom: A Potential Source of Ion Channel Modulators



Abstract
:1. Introduction
2. Diversity of Centipede Venoms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Toxin | Organism | Number of Residues | PDB | Ion Channels Activity |
|---|---|---|---|---|
| μ-SLPTX-Ssm6a (μ-SLPTX3-Ssm6a) | Scolopendra mutilans | 46 | 2MUN\2MZ4 | NaV1.7, IC50 of 25 nM |
| μ-SLPTX-Ssm1a | Scolopendra subspinipes | 32 | - | TTX-S NaV channel, IC50 of 9 nM |
| ω-SLPTX-Ssm1a | Scolopendra subspinipes | 83 | - | activator of Cav channels in DRG |
| ω-SLPTX-Ssm2a | Scolopendra subspinipes | 54 | - | CaV channels in DRG, IC50 of 1590 nM |
| SsmTx-I [20] | Scolopendra subspinipes mutilans | 36 | K+ channels in DRG, IC50 of 200 nM; KV2.1, IC50 of 41.7 nM; No effect on other K+ channels | |
| SsTx | Scolopendra mutilans | 53 | 5X0S | KV1.3, IC50 of 5.26 μM |
| SsdTx1-3 | Scolopendra subspinipes dehaani. | 53 | blocking hKir6.2 with a Kd of 278\260\281 nM | |
| SpTx1 | Scolopendra subspinipes dehaani. | 54 | Inhibiting hKir6.2 with a Kd of 8.5 nM | |
| SsTx-4 | S.subspinipes mutilans | 53 | Kir6.2, IC50 of 42.5 nM at 140 mV and 75.4 nM at 40 mV; Kir1.1, IC50 of 89.2 nM at 140 mV and 209.7 nM at 40 mV; Kir4.1, 360.1 nM at 140 mV and 6.2 μM at 40 mV | |
| RhTx | Scolopendra mutilans | 27 | 2MVA | TRPV1, EC50 of 521.5 nM |
| RhTx2 | Scolopendra mutilans | 31 | 3J5P\3J5Q | TRPV1, EC50 of 38.35 μM |
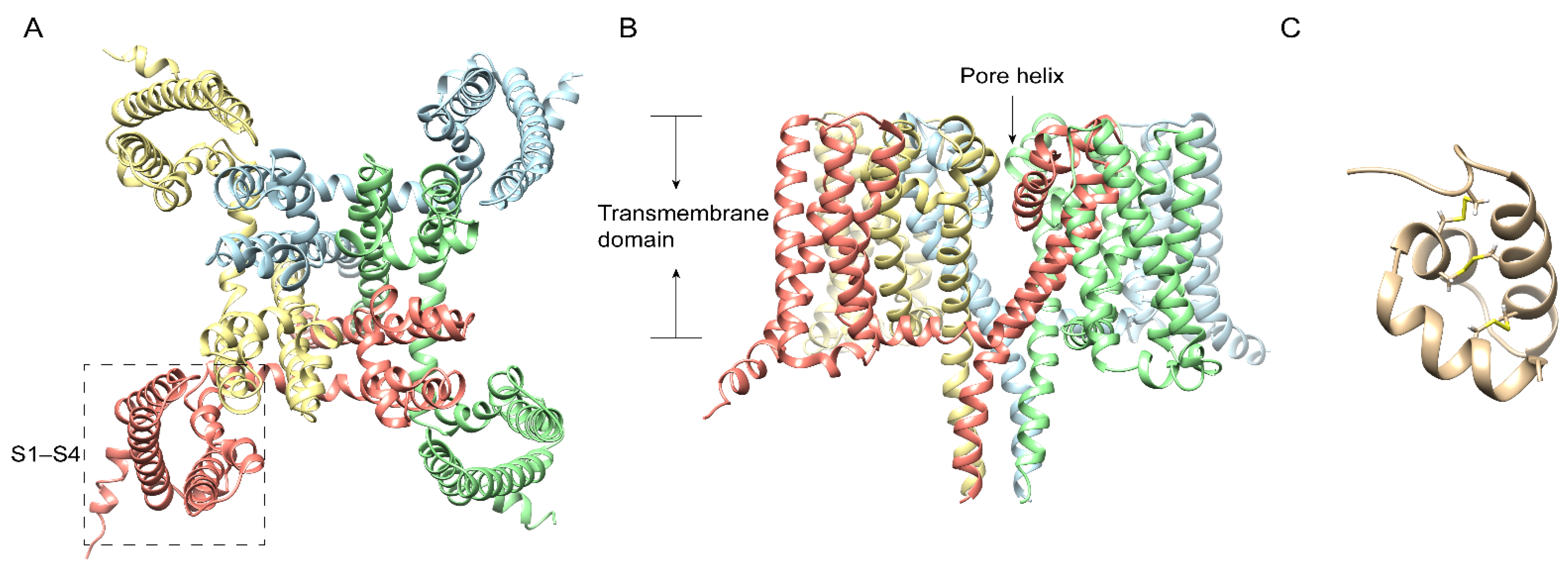
3. Centipede Venom Components Acting on NaV Channel
4. Centipede Venom Components Acting on Cav Channel
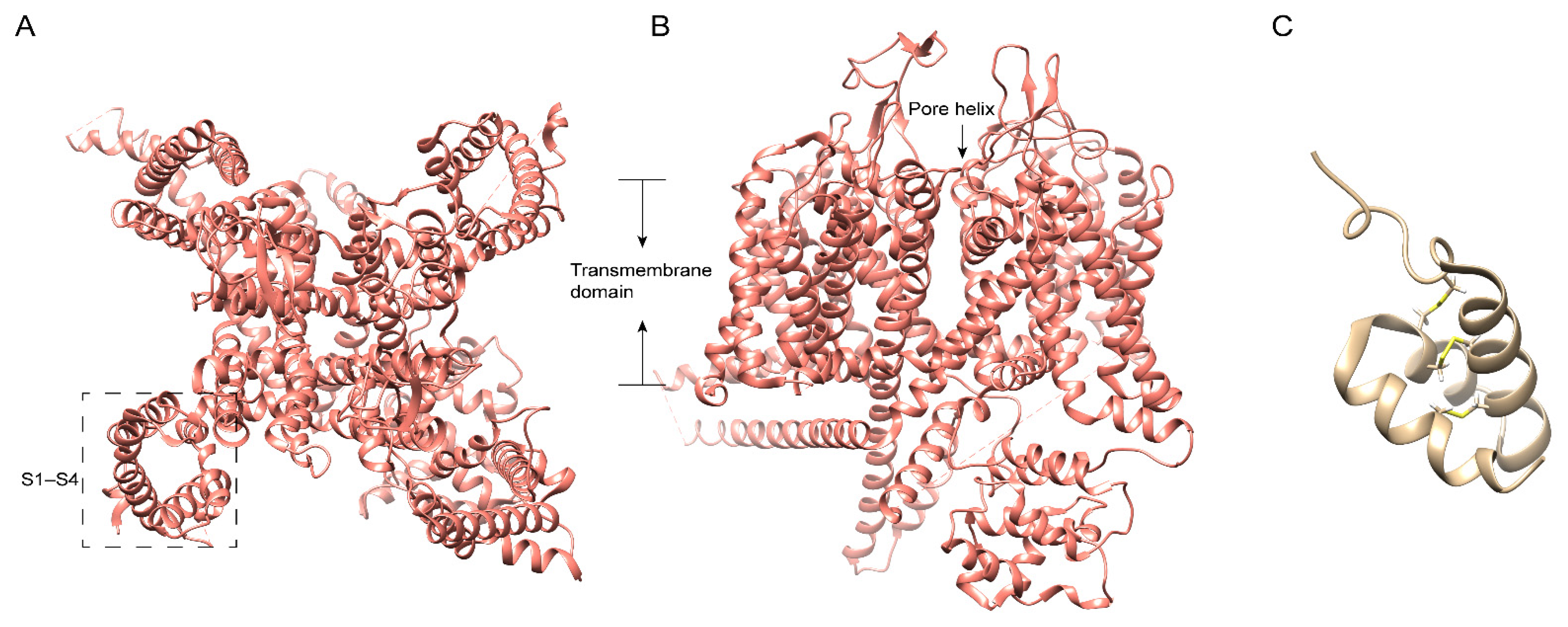
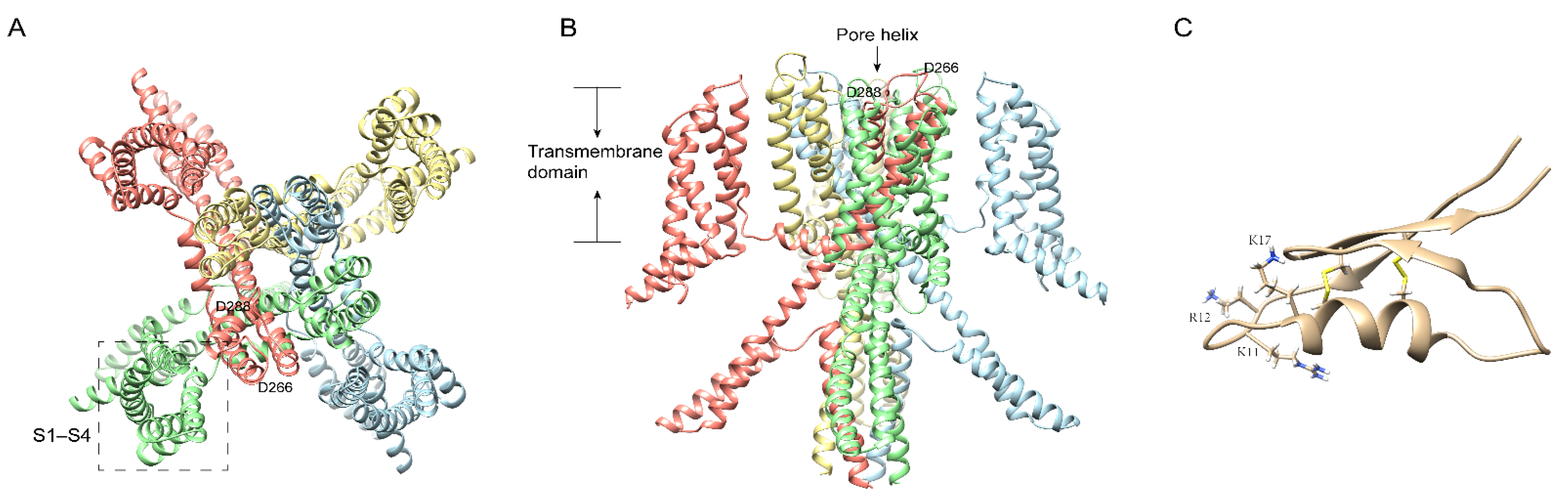
5. Centipede Venom Components Acting on the KV Channel
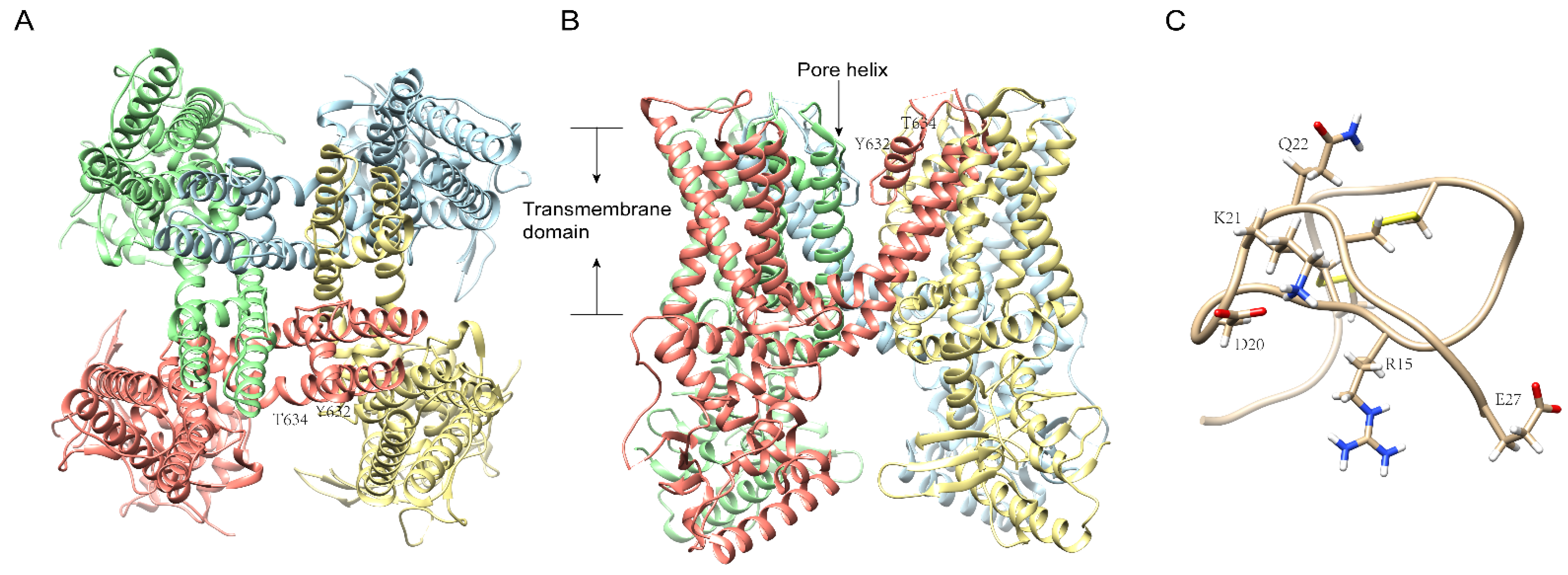
6. Centipede Venom Components Acting on TRP and Other Channels
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Edgecombe, G.D.; Giribet, G. Evolutionary biology of centipedes (Myriapoda: Chilopoda). Annu. Rev. Entomol. 2007, 52, 151–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veraldi, S.; Cuka, E.; Gaiani, F. Scolopendra bites: A report of two cases and review of the literature. Int. J. Dermatol. 2014, 53, 869–872. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, A.; Biceroglu, S.; Yakut, N.; Bilir, C.; Akdemir, R.; Akilli, A. Acute myocardial infarction in a young man caused by centipede sting. Emerg. Med. J. 2006, 23, e30. [Google Scholar] [CrossRef] [PubMed]
- Fung, H.T.; Lam, S.K.; Wong, O.F. Centipede bite victims: A review of patients presenting to two emergency departments in Hong Kong. Hong Kong Med. J. 2011, 17, 381–385. [Google Scholar]
- Ozsarac, M.; Karcioglu, O.; Ayrik, C.; Somuncu, F.; Gumrukcu, S. Acute coronary ischemia following centipede envenomation: Case report and review of the literature. Wilderness Environ. Med. 2004, 15, 109–112. [Google Scholar] [CrossRef]
- Du, C.; Li, J.; Shao, Z.; Mwangi, J.; Xu, R.; Tian, H.; Mo, G.; Lai, R.; Yang, S. Centipede KCNQ Inhibitor SsTx Also Targets KV1.3. Toxins 2019, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Liu, Z.; Xiao, Y.; Li, Y.; Rong, M.; Liang, S.; Zhang, Z.; Yu, H.; King, G.F.; Lai, R. Chemical punch packed in venoms makes centipedes excellent predators. Mol. Cell Proteom. 2012, 11, 640–650. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Xiao, Y.; Kang, D.; Liu, J.; Li, Y.; Undheim, E.A.; Klint, J.K.; Rong, M.; Lai, R.; King, G.F. Discovery of a selective NaV1.7 inhibitor from centipede venom with analgesic efficacy exceeding morphine in rodent pain models. Proc. Natl. Acad. Sci. USA 2013, 110, 17534–17539. [Google Scholar] [CrossRef] [Green Version]
- Rong, M.; Yang, S.; Wen, B.; Mo, G.; Kang, D.; Liu, J.; Lin, Z.; Jiang, W.; Li, B.; Du, C.; et al. Peptidomics combined with cDNA library unravel the diversity of centipede venom. J. Proteom. 2015, 114, 28–37. [Google Scholar] [CrossRef]
- Gonzalez-Morales, L.; Pedraza-Escalona, M.; Diego-Garcia, E.; Restano-Cassulini, R.; Batista, C.V.; Gutierrez Mdel, C.; Possani, L.D. Proteomic characterization of the venom and transcriptomic analysis of the venomous gland from the Mexican centipede Scolopendra viridis. J. Proteom. 2014, 111, 224–237. [Google Scholar] [CrossRef]
- Liu, Z.C.; Zhang, R.; Zhao, F.; Chen, Z.M.; Liu, H.W.; Wang, Y.J.; Jiang, P.; Zhang, Y.; Wu, Y.; Ding, J.P.; et al. Venomic and transcriptomic analysis of centipede Scolopendra subspinipes dehaani. J. Proteome. Res. 2012, 11, 6197–6212. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Qiu, P.; Yu, R. Centipede Venom Peptides Acting on Ion Channels. Toxins 2020, 12, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geron, M.; Hazan, A.; Priel, A. Animal Toxins Providing Insights into TRPV1 Activation Mechanism. Toxins 2017, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Vargas, J.M.; Possani, L.D.; Luna-Ramirez, K. Arthropod toxins acting on neuronal potassium channels. Neuropharmacology 2017, 127, 139–160. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yang, F.; Wei, N.; Hong, J.; Li, B.; Luo, L.; Rong, M.; Yarov-Yarovoy, V.; Zheng, J.; Wang, K.; et al. A pain-inducing centipede toxin targets the heat activation machinery of nociceptor TRPV1. Nat. Commun. 2015, 6, 8297. [Google Scholar] [CrossRef] [Green Version]
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A.B. The Diversity of Venom: The Importance of Behavior and Venom System Morphology in Understanding Its Ecology and Evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Undheim, E.A.B.; Jenner, R.A. Phylogenetic analyses suggest centipede venom arsenals were repeatedly stocked by horizontal gene transfer. Nat. Commun. 2021, 12, 818. [Google Scholar] [CrossRef]
- Ward, M.J.; Rokyta, D.R. Venom-gland transcriptomics and venom proteomics of the giant Florida blue centipede, Scolopendra viridis. Toxicon 2018, 152, 121–136. [Google Scholar] [CrossRef]
- Nystrom, G.S.; Ward, M.J.; Ellsworth, S.A.; Rokyta, D.R. Sex-based venom variation in the eastern bark centipede (Hemiscolopendra marginata). Toxicon 2019, 169, 45–58. [Google Scholar] [CrossRef]
- Chen, M.; Li, J.; Zhang, F.; Liu, Z. Isolation and characterization of SsmTx-I, a Specific Kv2.1 blocker from the venom of the centipede Scolopendra Subspinipes Mutilans L. Koch. J. Pept. Sci. 2014, 20, 159–164. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and molecular mechanisms of pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, G.F.; Escoubas, P.; Nicholson, G.M. Peptide toxins that selectively target insect Na(V) and Ca(V) channels. Channels 2008, 2, 100–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.H.; Catterall, W.A. Overview of the voltage-gated sodium channel family. Genome. Biol. 2003, 4, 207. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Shan, B.; Wang, Q.; Xu, Q.; Zhang, H.; Lei, H. Fusion of Ssm6a with a protein scaffold retains selectivity on NaV 1.7 and improves its therapeutic potential against chronic pain. Chem. Biol. Drug Des. 2017, 89, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Voltage-gated calcium channels. Cold Spring Harb. Perspect. Biol. 2011, 3, a003947. [Google Scholar] [CrossRef]
- Sousa, S.R.; Vetter, I.; Lewis, R.J. Venom peptides as a rich source of cav2.2 channel blockers. Toxins 2013, 5, 286–314. [Google Scholar] [CrossRef] [Green Version]
- Saegusa, H.; Kurihara, T.; Zong, S.; Kazuno, A.; Matsuda, Y.; Nonaka, T.; Han, W.; Toriyama, H.; Tanabe, T. Suppression of inflammatory and neuropathic pain symptoms in mice lacking the N-type Ca2+ channel. EMBO J. 2001, 20, 2349–2356. [Google Scholar] [CrossRef]
- Hatakeyama, S.; Wakamori, M.; Ino, M.; Miyamoto, N.; Takahashi, E.; Yoshinaga, T.; Sawada, K.; Imoto, K.; Tanaka, I.; Yoshizawa, T.; et al. Differential nociceptive responses in mice lacking the alpha(1B) subunit of N-type Ca(2+) channels. Neuroreport 2001, 12, 2423–2427. [Google Scholar] [CrossRef]
- Shieh, C.C.; Coghlan, M.; Sullivan, J.P.; Gopalakrishnan, M. Potassium channels: Molecular defects, diseases, and therapeutic opportunities. Pharmacol. Rev. 2000, 52, 557–594. [Google Scholar]
- Gutman, G.A.; Chandy, K.G.; Grissmer, S.; Lazdunski, M.; McKinnon, D.; Pardo, L.A.; Robertson, G.A.; Rudy, B.; Sanguinetti, M.C.; Stuhmer, W.; et al. International Union of Pharmacology. LIII. Nomenclature and molecular relationships of voltage-gated potassium channels. Pharmacol. Rev. 2005, 57, 473–508. [Google Scholar] [CrossRef]
- Hibino, H.; Inanobe, A.; Furutani, K.; Murakami, S.; Findlay, I.; Kurachi, Y. Inwardly rectifying potassium channels: Their structure, function, and physiological roles. Physiol. Rev. 2010, 90, 291–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.H.; Yarov-Yarovoy, V.; Gutman, G.A.; Catterall, W.A. Overview of molecular relationships in the voltage-gated ion channel superfamily. Pharmacol. Rev. 2005, 57, 387–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.N.; Herrington, J.; Petrov, A.; Ge, L.; Eiermann, G.; Xiong, Y.; Jensen, M.V.; Hohmeier, H.E.; Newgard, C.B.; Garcia, M.L.; et al. The role of voltage-gated potassium channels Kv2.1 and Kv2.2 in the regulation of insulin and somatostatin release from pancreatic islets. J. Pharmacol. Exp Ther. 2013, 344, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beeton, C.; Pennington, M.W.; Wulff, H.; Singh, S.; Nugent, D.; Crossley, G.; Khaytin, I.; Calabresi, P.A.; Chen, C.Y.; Gutman, G.A.; et al. Targeting effector memory T cells with a selective peptide inhibitor of Kv1.3 channels for therapy of autoimmune diseases. Mol. Pharmacol. 2005, 67, 1369–1381. [Google Scholar] [CrossRef]
- Wulff, H.; Calabresi, P.A.; Allie, R.; Yun, S.; Pennington, M.; Beeton, C.; Chandy, K.G. The voltage-gated Kv1.3 K(+) channel in effector memory T cells as new target for MS. J. Clin. Investig. 2003, 111, 1703–1713. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Li, B.; Wang, S.; Wu, F.; Wang, X.; Liang, P.; Ombati, R.; Chen, J.; Lu, X.; Cui, J.; et al. Centipedes subdue giant prey by blocking KCNQ channels. Proc. Natl. Acad. Sci. USA 2018, 115, 1646–1651. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Wang, Y.; Wang, L.; Kamau, P.; Zhang, H.; Luo, A.; Lu, X.; Lai, R. Target switch of centipede toxins for antagonistic switch. Sci. Adv. 2020, 6, eabb5734. [Google Scholar] [CrossRef]
- Ramu, Y.; Lu, Z. A family of orthologous proteins from centipede venoms inhibit the hKir6.2 channel. Sci. Rep. 2019, 9, 14088. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Xu, J.; Li, Y.; Zhao, P.; Kong, X.; Hu, H.; Liang, S.; Tang, C.; Liu, Z. Molecular mechanisms of centipede toxin SsTx-4 inhibition of inwardly rectifying potassium channels. J. Biol. Chem. 2021, 297, 101076. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Jordt, S.E.; Tominaga, M.; Julius, D. Acid potentiation of the capsaicin receptor determined by a key extracellular site. Proc. Natl. Acad. Sci. USA 2000, 97, 8134–8139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Winter, Z.; Buhala, A.; Otvos, F.; Josvay, K.; Vizler, C.; Dombi, G.; Szakonyi, G.; Olah, Z. Functionally important amino acid residues in the transient receptor potential vanilloid 1 (TRPV1) ion channel—An overview of the current mutational data. Mol. Pain 2013, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, M.Z.; Mtui, T.; Gao, Y.D.; Kohler, M.; Middleton, R.E. Resiniferatoxin binds to the capsaicin receptor (TRPV1) near the extracellular side of the S4 transmembrane domain. Biochemistry 2004, 43, 2501–2511. [Google Scholar] [CrossRef]
- Gao, R.; Meng, M.; Zhou, X.; Yu, M.; Li, Z.; Li, J.; Wang, X.; Song, Y.; Wang, H.; He, J. TRPV1, a novel biomarker associated with lung cancer via excluding immune infiltration. MedComm 2022, 3, e139. [Google Scholar] [CrossRef]
- Zhu, A.; Aierken, A.; Yao, Z.; Vu, S.; Tian, Y.; Zheng, J.; Yang, S.; Yang, F. A centipede toxin causes rapid desensitization of nociceptor TRPV1 ion channel. Toxicon 2020, 178, 41–49. [Google Scholar] [CrossRef]
- Ji, X.; Naito, Y.; Hirokawa, G.; Weng, H.; Hiura, Y.; Takahashi, R.; Iwai, N. P2X(7) receptor antagonism attenuates the hypertension and renal injury in Dahl salt-sensitive rats. Hypertens Res. 2012, 35, 173–179. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic Signalling: Therapeutic Developments. Front. Pharmacol. 2017, 8, 661. [Google Scholar] [CrossRef] [Green Version]
- Ford, A.P.; Undem, B.J. The therapeutic promise of ATP antagonism at P2X3 receptors in respiratory and urological disorders. Front. Cell Neurosci. 2013, 7, 267. [Google Scholar] [CrossRef] [Green Version]
- Burnstock, G. P2X ion channel receptors and inflammation. Purinergic. Signal. 2016, 12, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Cieslak, M.; Czarnecka, J.; Roszek, K.; Komoszynski, M. The role of purinergic signaling in the etiology of migraine and novel antimigraine treatment. Purinergic. Signal. 2015, 11, 307–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuda, M. P2 receptors, microglial cytokines and chemokines, and neuropathic pain. J. Neurosci. Res. 2017, 95, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.R.; Castelo-Branco, M.T.; Figliuolo, V.R.; Bernardazzi, C.; Buongusto, F.; Yoshimoto, A.; Nanini, H.F.; Coutinho, C.M.; Carneiro, A.J.; Coutinho-Silva, R.; et al. Overexpression of ATP-activated P2X7 receptors in the intestinal mucosa is implicated in the pathogenesis of Crohn’s disease. Inflamm. Bowel. Dis. 2014, 20, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Rassendren, F.; Audinat, E. Purinergic signaling in epilepsy. J. Neurosci. Res. 2016, 94, 781–793. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, D.; Vitiello, L.; Idzko, M.; la Sala, A. Purinergic signaling in atherosclerosis. Trends. Mol. Med. 2015, 21, 184–192. [Google Scholar] [CrossRef]
- Vassilevski, A.A.; Kozlov, S.A.; Grishin, E.V. Molecular diversity of spider venom. Biochemistry 2009, 74, 1505–1534. [Google Scholar] [CrossRef]
- Bibic, L.; Herzig, V.; King, G.F.; Stokes, L. Development of High-Throughput Fluorescent-Based Screens to Accelerate Discovery of P2X Inhibitors from Animal Venoms. J. Nat. Prod. 2019, 82, 2559–2567. [Google Scholar] [CrossRef]
- Kularatne, S.A.; Senanayake, N. Venomous snake bites, scorpions, and spiders. Handb. Clin. Neurol. 2014, 120, 987–1001. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, A.; Wang, A.; Kamau, P.M.; Lai, R.; Luo, L. Centipede Venom: A Potential Source of Ion Channel Modulators. Int. J. Mol. Sci. 2022, 23, 7105. https://doi.org/10.3390/ijms23137105
Luo A, Wang A, Kamau PM, Lai R, Luo L. Centipede Venom: A Potential Source of Ion Channel Modulators. International Journal of Molecular Sciences. 2022; 23(13):7105. https://doi.org/10.3390/ijms23137105
Chicago/Turabian StyleLuo, Anna, Aili Wang, Peter Muiruri Kamau, Ren Lai, and Lei Luo. 2022. "Centipede Venom: A Potential Source of Ion Channel Modulators" International Journal of Molecular Sciences 23, no. 13: 7105. https://doi.org/10.3390/ijms23137105
APA StyleLuo, A., Wang, A., Kamau, P. M., Lai, R., & Luo, L. (2022). Centipede Venom: A Potential Source of Ion Channel Modulators. International Journal of Molecular Sciences, 23(13), 7105. https://doi.org/10.3390/ijms23137105