
miR-29c Increases Protein Synthesis in Skeletal Muscle Independently of AKT/mTOR

 ,
,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
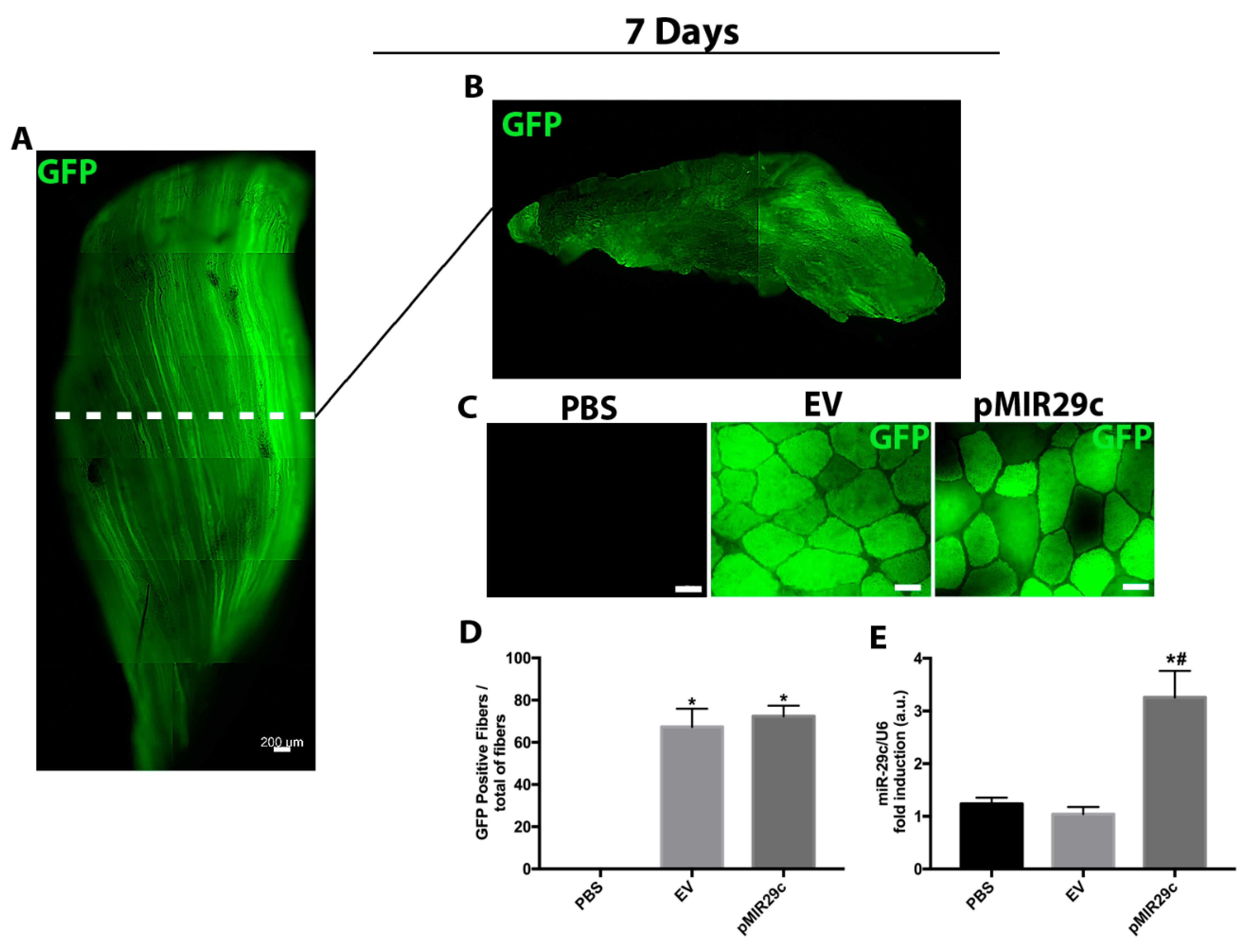
2.1. GFP Transfection Efficiency and Overexpression of miR-29c Results in Skeletal Muscle
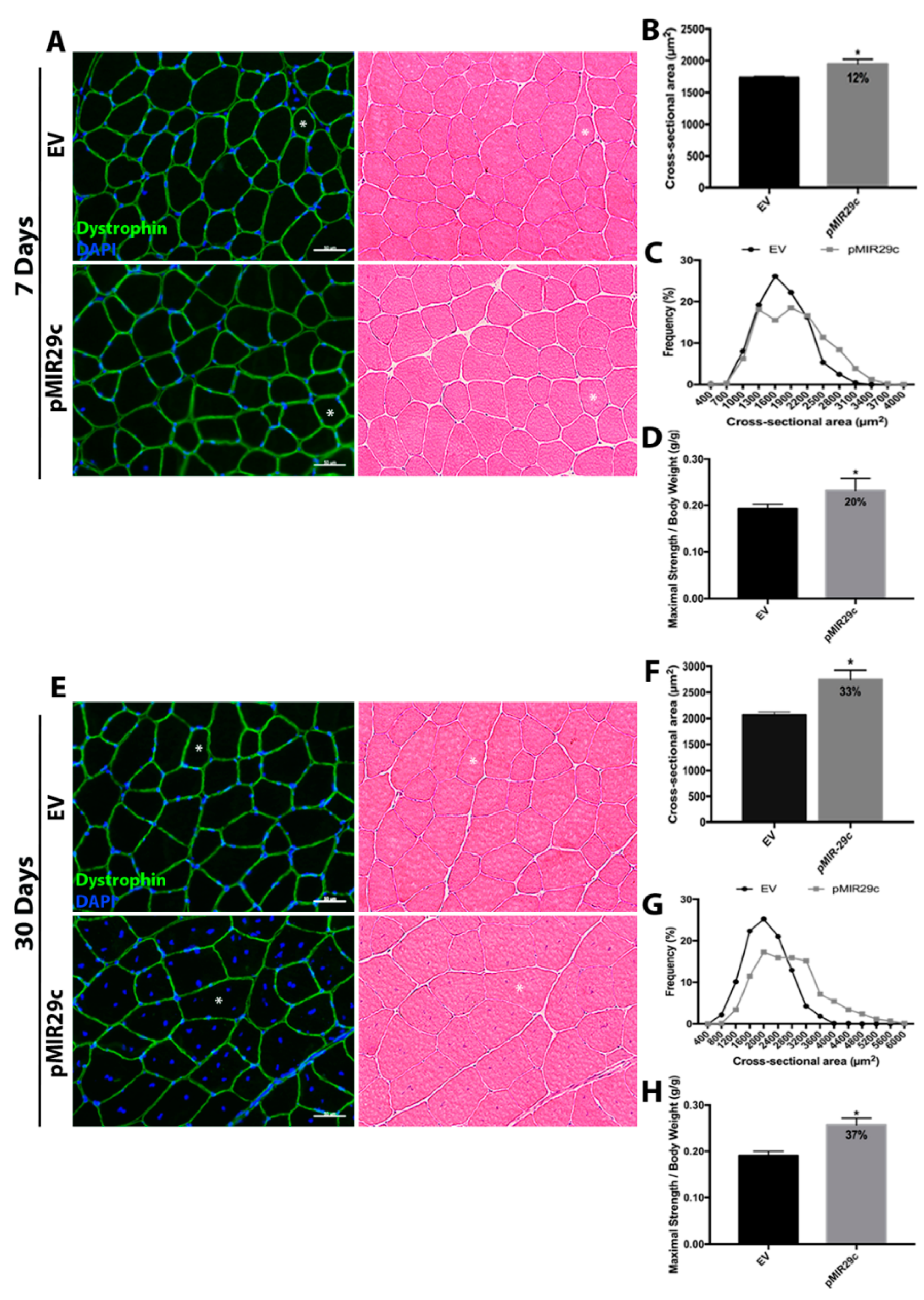
2.2. Impact of miR-29c Overexpression on Skeletal Muscle Mass and Function
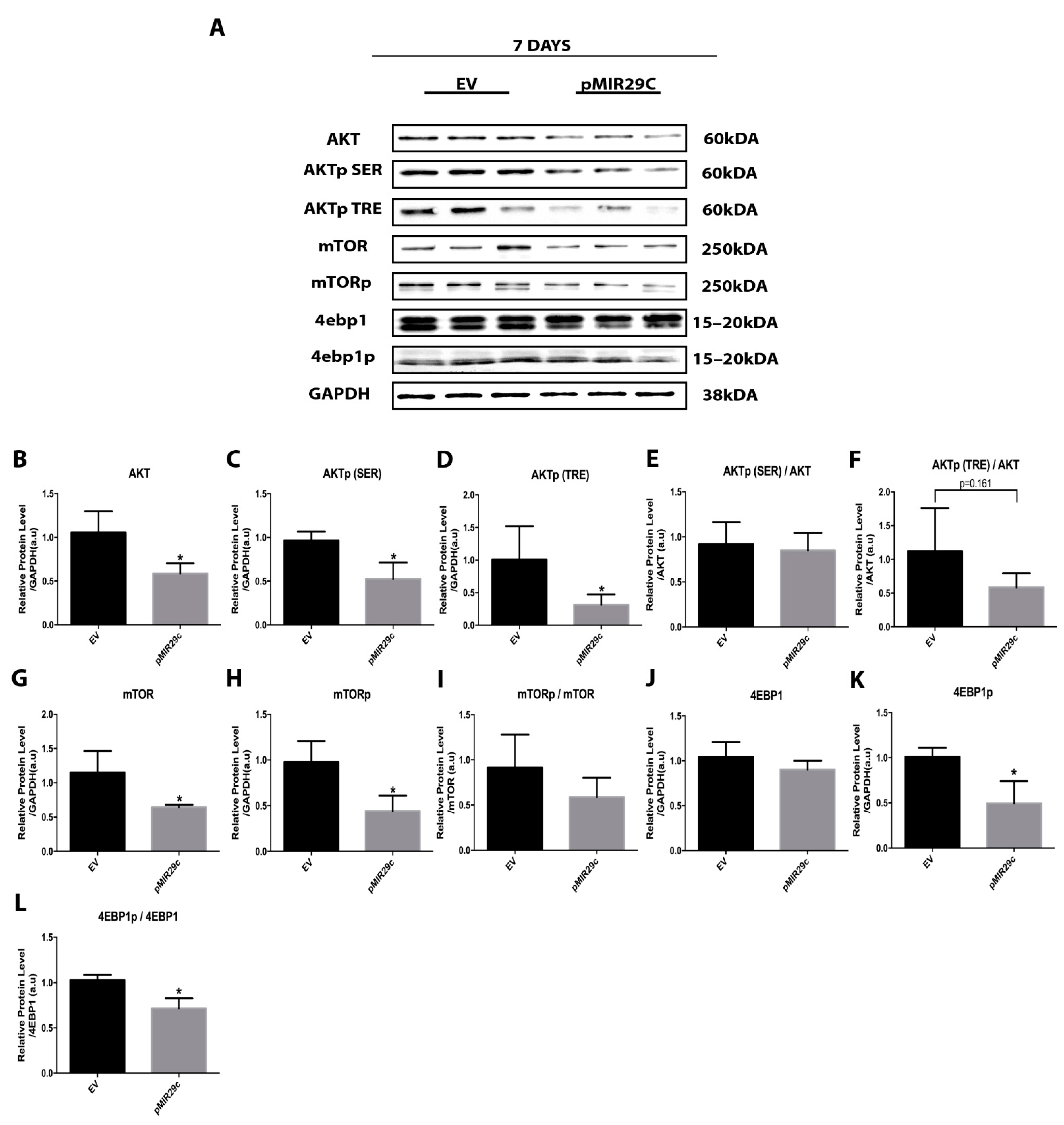
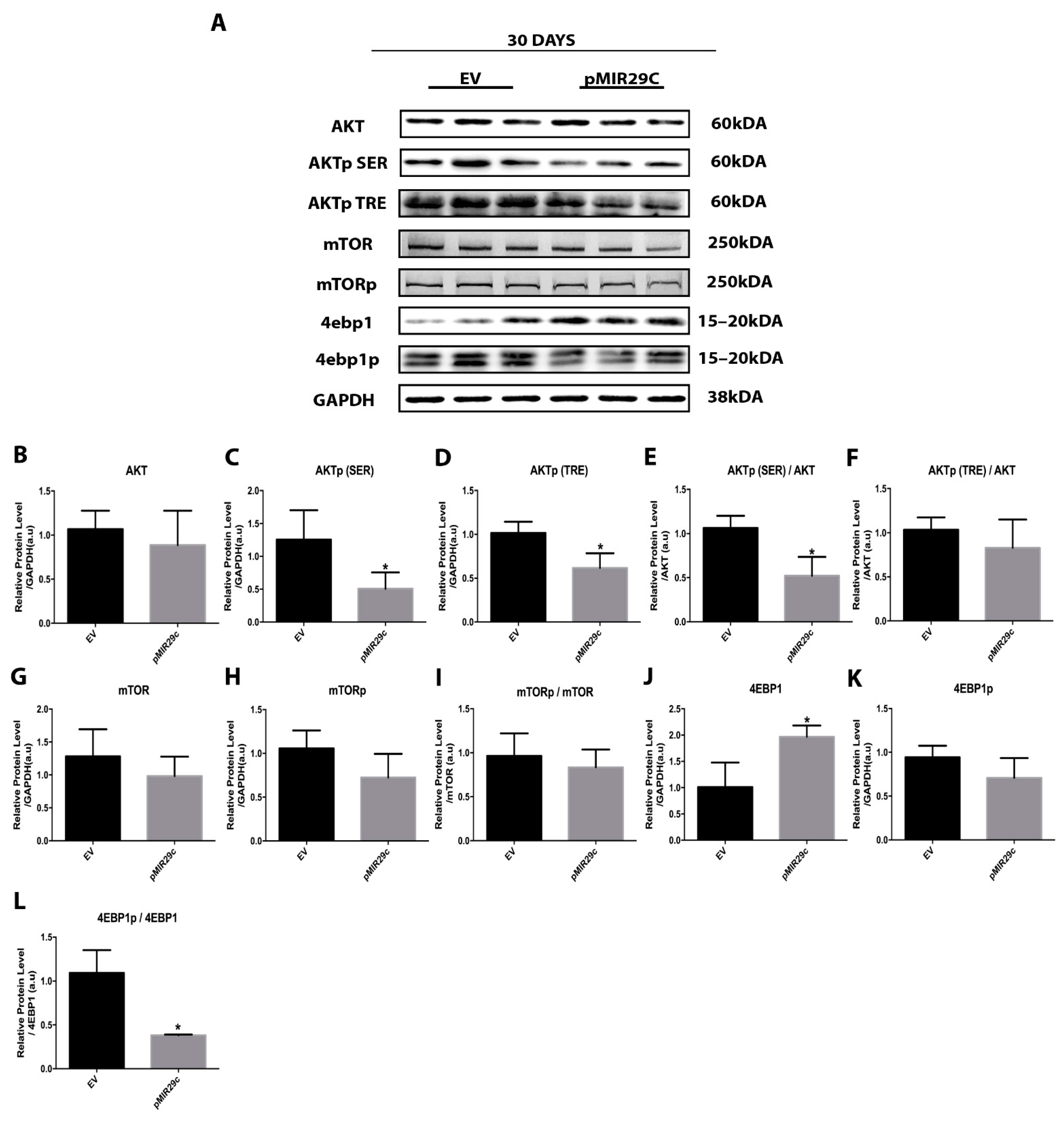
2.3. Impact of miR-29c Overexpression on the AKT-mTOR Pathway
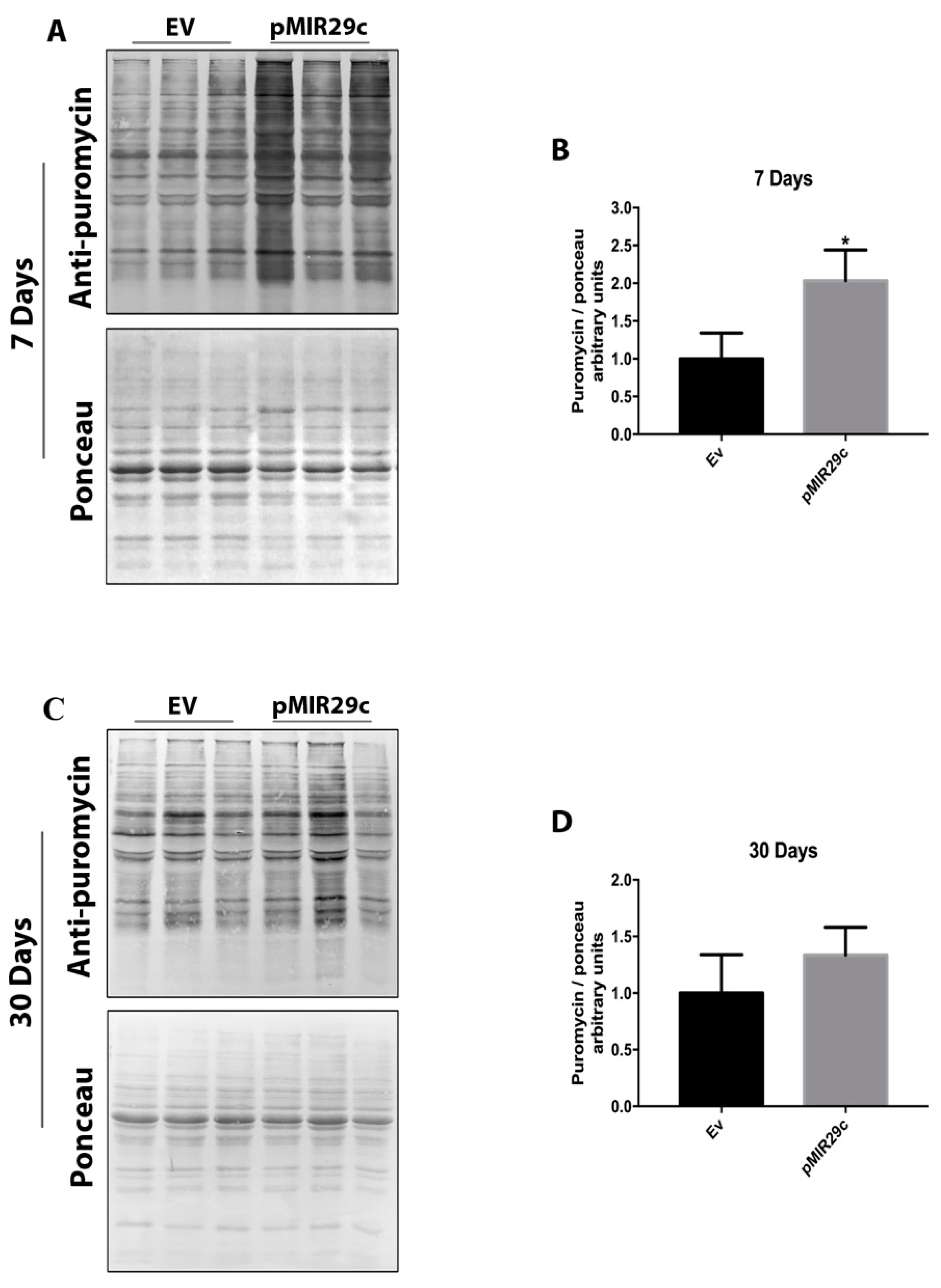
2.4. Impact of miR-29c Overexpression on Protein Synthesis
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Muscle Transfection by Eletrotransfer
4.3. TA Morphometric Analysis
4.4. Puromycin Assay
4.5. Maximal Strength Test (1RM)
4.6. Western Blotting
4.7. RNA Extraction and Real-Time PCR
4.8. Statistical Analysis
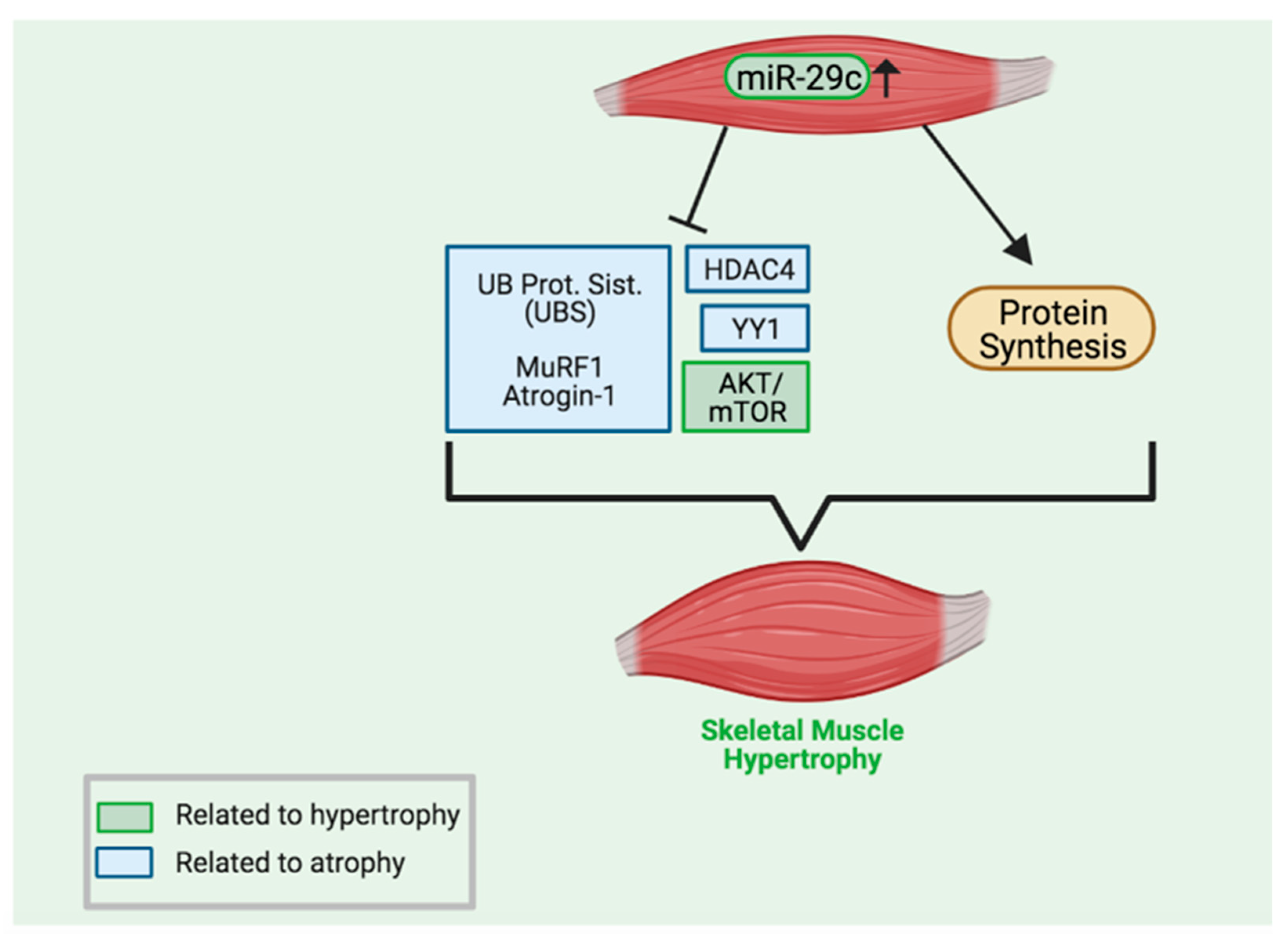
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, B.; Lin, Z.; Pu, W.T. Mammalian Myocardial Regeneration. In Muscle; Elsevier: Amsterdam, The Netherlands, 2012; pp. 555–569. ISBN 978-0-12-381510-1. [Google Scholar]
- Janssen, I.; Heymsfield, S.B.; Wang, Z.M.; Ross, R. Skeletal muscle mass and distribution in 468 men and women aged 18–88 yr. J. Appl. Physiol. 2000, 89, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Béchet, D.; Tassa, A.; Combaret, L.; Taillandier, D.; Attaix, D. Regulation of skeletal muscle proteolysis by amino acids. J. Ren. Nutr. 2005, 15, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, A.L. Protein turnover in skeletal muscle: II. Effects of denervation and cortisone on protein catabolism in skeletal muscle. J. Biol. Chem. 1969, 244, 3223–3229. [Google Scholar] [CrossRef]
- Goldberg, A.L. Protein turnover in skeletal muscle: I. Protein catabolism during work-induced hypertrophy and growth induced with growth hormone. J. Biol. Chem. 1969, 244, 3217–3222. [Google Scholar] [CrossRef]
- Peris-Moreno, D.; Taillandier, D.; Polge, C. MuRF1/TRIM63, Master Regulator of Muscle Mass. Int. J. Mol. Sci. 2020, 21, 6663. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef] [PubMed]
- Vainshtein, A.; Sandri, M. Signaling Pathways That Control Muscle Mass. Int. J. Mol. Sci. 2020, 21, 4759. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.H.; Yuen, K.W.Y. Point centromere activity requires an optimal level of centromeric noncoding RNA. Proc. Natl. Acad. Sci. USA 2019, 116, 6270–6279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, N.; Kropp, J.; Khatib, H. MicroRNA Signaling in Embryo Development. Biology 2017, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.J.; Graça, F.A.; Cruz, A.; Silvestre, J.G.; Labeit, S.; Miyabara, E.H.; Yan, C.Y.I.; Wang, D.Z.; Moriscot, A.S. miR-29c improves skeletal muscle mass and function throughout myocyte proliferation and differentiation and by repressing atrophy-related genes. Acta Physiol. Oxf. Engl. 2019, 226, e13278. [Google Scholar] [CrossRef] [PubMed]
- Falcone, G. A new role of miR-29c as a potent inducer of skeletal muscle hypertrophy. Acta Physiol. Oxf. Engl. 2019, 226, e13320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takegaki, J.; Ogasawara, R.; Tamura, Y.; Takagi, R.; Arihara, Y.; Tsutaki, A.; Nakazato, K.; Ishii, N. Repeated bouts of resistance exercise with short recovery periods activates mTOR signaling, but not protein synthesis, in mouse skeletal muscle. Physiol. Rep. 2017, 5, e13515. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, K.; Yamaguchi, A. The Functional Role of Calcineurin in Hypertrophy, Regeneration, and Disorders of Skeletal Muscle. J. Biomed. Biotechnol. 2010, 2010, 721219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, D.D.; Wong, V.L.; Esser, K.A. Expression of beta-catenin is necessary for physiological growth of adult skeletal muscle. Am. J. Physiol. Cell Physiol. 2006, 291, C185–C188. [Google Scholar] [CrossRef] [PubMed]
- Lynch, G.S.; Ryall, J.G. Role of beta-adrenoceptor signaling in skeletal muscle: Implications for muscle wasting and disease. Physiol. Rev. 2008, 88, 729–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruas, J.L.; White, J.P.; Rao, R.R.; Kleiner, S.; Brannan, K.T.; Harrison, B.C.; Greene, N.P.; Wu, J.; Estall, J.L.; Irving, B.A.; et al. A PGC-1α isoform induced by resistance training regulates skeletal muscle hypertrophy. Cell 2012, 151, 1319–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarthy, J.J.; Murach, K.A. Chapter 24—Anabolic and Catabolic Signaling Pathways That Regulate Skeletal Muscle Mass. In Nutrition and Enhanced Sports Performance, 2nd ed.; Bagchi, D., Nair, S., Sen, C.K., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 275–290. ISBN 978-0-12-813922-6. [Google Scholar]
- Wang, H.; Garzon, R.; Sun, H.; Ladner, K.J.; Singh, R.; Dahlman, J.; Cheng, A.; Hall, B.M.; Qualman, S.J.; Chandler, D.S.; et al. NF-κB–YY1–miR-29 Regulatory Circuitry in Skeletal Myogenesis and Rhabdomyosarcoma. Cancer Cell 2008, 14, 369–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, C.A.; Mabrey, D.M.; Frey, J.W.; Miu, M.H.; Schmidt, E.K.; Pierre, P.; Hornberger, T.A. Novel insights into the regulation of skeletal muscle protein synthesis as revealed by a new nonradioactive in vivo technique. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 1028–1039. [Google Scholar] [CrossRef] [Green Version]
- Adams, V.; Gußen, V.; Zozulya, S.; Cruz, A.; Moriscot, A.; Linke, A.; Labeit, S. Small-Molecule Chemical Knockdown of MuRF1 in Melanoma Bearing Mice Attenuates Tumor Cachexia Associated Myopathy. Cells 2020, 9, 2272. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves, P.K.N.; Cruz, A.; Silva, W.J.; Labeit, S.; Moriscot, A.S. miR-29c Increases Protein Synthesis in Skeletal Muscle Independently of AKT/mTOR. Int. J. Mol. Sci. 2022, 23, 7198. https://doi.org/10.3390/ijms23137198
Alves PKN, Cruz A, Silva WJ, Labeit S, Moriscot AS. miR-29c Increases Protein Synthesis in Skeletal Muscle Independently of AKT/mTOR. International Journal of Molecular Sciences. 2022; 23(13):7198. https://doi.org/10.3390/ijms23137198
Chicago/Turabian StyleAlves, Paula Ketilly Nascimento, André Cruz, William J. Silva, Siegfried Labeit, and Anselmo Sigari Moriscot. 2022. "miR-29c Increases Protein Synthesis in Skeletal Muscle Independently of AKT/mTOR" International Journal of Molecular Sciences 23, no. 13: 7198. https://doi.org/10.3390/ijms23137198
APA StyleAlves, P. K. N., Cruz, A., Silva, W. J., Labeit, S., & Moriscot, A. S. (2022). miR-29c Increases Protein Synthesis in Skeletal Muscle Independently of AKT/mTOR. International Journal of Molecular Sciences, 23(13), 7198. https://doi.org/10.3390/ijms23137198