
Transport of Biologically Active Ultrashort Peptides Using POT and LAT Carriers


 ,
,
Abstract
:1. Introduction
2. POT Family Peptide Transporters (SCL15): PEPT1, PEPT2, PHT1, PHT2
3. Localization of the POT Family Members in Various Organs and Tissues
4. Participation of the POT Family Transporters in Various Physiological Processes under Normal and Pathological Conditions
5. Role of PEPT1 and PEPT2 in the Transport of Ultrashort Peptides
6. Amino-Acid Transporters LAT1 and LAT2: Possible Involvement in Peptide Transport
7. Participation of LAT1 and LAT2 in Various Physiological Processes during Ageing and Pathology
8. Possible Mechanisms for Ligand Transport by LAT1
9. Discussion
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Nomenclature and Symbolism for Amino Acids and Peptides. Recommendations 1983. Eur. J. Biochem. 1984, 138, 9–37. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulos, V.; Bojarska, J.; Chai, T.-T.; Elnagdy, S.; Kaczmarek, K.; Matsoukas, J.; New, R.; Parang, K.; Lopez, O.P.; Parhiz, H.; et al. A Global Review on Short Peptides: Frontiers and Perspectives. Molecules 2021, 26, 430. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry, 4th ed.; Freeman and Company: New York, NY, USA, 2015. [Google Scholar]
- Hoffknecht, B.C.; Albada, H.B.; Sturm, M.; Prochnow, P.; Bandow, J.E.; Metzler-Nolte, N. Synthesis and Antibacterial Activity of Trivalent Ultrashort Arg-Trp-Based Antimicrobial Peptides (AMPs). Med. Chem. Commun. 2015, 6, 372–376. [Google Scholar] [CrossRef]
- Ni, M.; Tresset, G.; Iliescu, C.; Hauser, C.A.E. Ultrashort Peptide Theranostic Nanoparticles by Microfluidic-Assisted Rapid Solvent Exchange. IEEE Trans. NanoBiosci. 2020, 19, 627–632. [Google Scholar] [CrossRef]
- Ghalayini, S.; Susapto, H.H.; Hall, S.; Kahin, K.; Hauser, C.A.E. Preparation and printability of ultrashort self-assembling peptide nanoparticles. Int. J. Bioprint. 2019, 5, 239. [Google Scholar] [CrossRef]
- Salvagni, E.; Garcia, C.; Manresa, A.; Müller-Sanchez, C.; Reina, M.; Rodriguez-Abreu, C.; Garcia-Celma, M.J.; Esquena, J. Short and ultrashort antimicrobial peptides anchored onto soft commercial contact lenses inhibit bacterial adhesion. Colloids Surf. B Biointerfaces 2020, 196, 111283. [Google Scholar] [CrossRef]
- Abuine, R.; Rathnayake, A.U.; HeeGuk, B. Biological Activity of Peptides Purified from Fish Skin Hydrolysates. Fish. Aquat. Sci. 2019, 22, 10. [Google Scholar] [CrossRef] [Green Version]
- Ciociola, T.; Giovati, L.; Conti, S.; Magliani, W.; Santinoli, C.; Polonelli, L. Natural and Synthetic Peptides with Antifungal Activity. Future Med. Chem. 2016, 8, 1413–1433. [Google Scholar] [CrossRef]
- Khavinson, V.K. Peptide medicines: Past, present, future. Clin. Med. 2020, 98, 165–177. [Google Scholar] [CrossRef]
- Chervyakova, N.A.; Linkova, N.S.; Chalisova, N.I.; Koncevaya, E.A.; Trofimova, S.V.; Khavinson, V.K. Molecular aspects of immunoprotective activity of peptides in spleen during the ageing process. Adv. Gerontol. 2014, 27, 224–228. [Google Scholar]
- Sinjari, B.; Diomede, F.; Khavinson, V.; Mironova, E.; Linkova, N.; Trofimova, S.; Trubiani, O.; Caputi, S. Short Peptides Protect Oral Stem Cells from Ageing. Stem. Cell Rev. Rep. 2020, 16, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.K.; Tarnovskaia, S.I.; Lin’kova, N.S.; Guton, E.O.; Elashkina, E.V. Epigenetic aspects of peptidergic regulation of vascular endothelial cell proliferation during aging. Adv. Gerontol. 2014, 27, 108–114. [Google Scholar] [PubMed]
- Khavinson, V.K.; Popovich, I.G.; Linkova, N.S.; Mironova, E.S.; Ilina, A.R. Peptide Regulation of Gene Expression: A Systematic Review. Molecules 2021, 26, 7053. [Google Scholar] [CrossRef] [PubMed]
- Zuglianello, C.; Lemos-Senna, E. The nanotechnological approach for nasal delivery of peptide drugs: A comprehensive review. J. Microencapsul. 2022, 39, 156–175. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Dong, X.; Yang, F.; Zhou, S.; Xiang, M.; Lou, L.; Yao, S.Q.; Gao, L. Engineered Cell-Penetrating Peptides for Mitochondrion-Targeted Drug Delivery in Cancer Therapy. Chemistry 2021, 27, 14721–14729. [Google Scholar] [CrossRef]
- Allolio, C.; Magarkar, A.; Jurkiewicz, P.; Baxová, K.; Javanainen, M.; Mason, P.E.; Šachl, R.; Cebecauer, M.; Hof, M.; Horinek, D.; et al. Arginine-Rich Cell-Penetrating Peptides Induce Membrane Multilamellarity and Subsequently Enter via Formation of a Fusion Pore. Proc. Natl. Acad. Sci. USA 2018, 115, 11923–11928. [Google Scholar] [CrossRef] [Green Version]
- Terada, T.; Sawada, K.; Irie, M.; Saito, H.; Hashimoto, Y.; Inui, K. Structural Requirements for Determining the Substrate Affinity of Peptide Transporters PEPT1 and PEPT2. Pflug. Arch. 2000, 440, 679–684. [Google Scholar] [CrossRef]
- Campos-Bedolla, P.; Walter, F.R.; Veszelka, S.; Deli, M.A. Role of the Blood-Brain Barrier in the Nutrition of the Central Nervous System. Arch. Med. Res. 2014, 45, 610–638. [Google Scholar] [CrossRef]
- del Amo, E.M.; Urtti, A.; Yliperttula, M. Pharmacokinetic Role of L-Type Amino Acid Transporters LAT1 and LAT2. Eur. J. Pharm. Sci. 2008, 35, 161–174. [Google Scholar] [CrossRef]
- Kamal, M.A.; Keep, R.F.; Smith, D.E. Role and Relevance of PEPT2 in Drug Disposition, Dynamics, and Toxicity. Drug Metab. Pharmacokinet. 2008, 23, 236–242. [Google Scholar] [CrossRef] [Green Version]
- Saito, H.; Okuda, M.; Terada, T.; Sasaki, S.; Inui, K. Cloning and Characterization of a Rat H+/Peptide Cotransporter Mediating Absorption of Beta-Lactam Antibiotics in the Intestine and Kidney. J. Pharmacol. Exp. Ther. 1995, 275, 1631–1637. [Google Scholar] [PubMed]
- Saito, H.; Terada, T.; Okuda, M.; Sasaki, S.; Inui, K. Molecular Cloning and Tissue Distribution of Rat Peptide Transporter PEPT2. Biochim. Biophys. Acta 1996, 1280, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Bröer, S. Amino Acid Transport across Mammalian Intestinal and Renal Epithelia. Physiol. Rev. 2008, 88, 249–286. [Google Scholar] [CrossRef]
- Daniel, H.; Kottra, G. The Proton Oligopeptide Cotransporter Family SLC15 in Physiology and Pharmacology. Pflug. Arch. 2004, 447, 610–618. [Google Scholar] [CrossRef]
- Smith, D.E.; Clémençon, B.; Hediger, M.A. Proton-Coupled Oligopeptide Transporter Family SLC15: Physiological, Pharmacological and Pathological Implications. Mol. Asp. Med. 2013, 34, 323–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, D.J.; Leibach, F.H.; Ganapathy, V.; Thwaites, D.T. Optimal Absorptive Transport of the Dipeptide Glycylsarcosine Is Dependent on Functional Na+/H+ Exchange Activity. Pflug. Arch. 2002, 445, 139–146. [Google Scholar] [CrossRef]
- Daniel, H.; Spanier, B.; Kottra, G.; Weitz, D. From Bacteria to Man: Archaic Proton-Dependent Peptide Transporters at Work. Physiology 2006, 21, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Nabulsi, N.B.; Smith, D.E.; Kilbourn, M.R. [11C] Glycylsarcosine: Synthesis and in Vivo Evaluation as a PET Tracer of PepT2 Transporter Function in Kidney of PepT2 Null and Wild-Type Mice. Bioorg. Med. Chem. 2005, 13, 2993–3001. [Google Scholar] [CrossRef]
- Meissner, B.; Boll, M.; Daniel, H.; Baumeister, R. Deletion of the Intestinal Peptide Transporter Affects Insulin and TOR Signaling in Caenorhabditis Elegans. J. Biol. Chem. 2004, 279, 36739–36745. [Google Scholar] [CrossRef] [Green Version]
- Verri, T.; Kottra, G.; Romano, A.; Tiso, N.; Peric, M.; Maffia, M.; Boll, M.; Argenton, F.; Daniel, H.; Storelli, C. Molecular and Functional Characterisation of the Zebrafish (Danio Rerio) PEPT1-Type Peptide Transporter11Sequence Data Submission and Accession Numbers: Sequence Data Reported Are Available in the DDBJ/EMBL/GenBank Databases under Accession Nos. AY300011 (Zebrafish Pept1) and AY300010 (Zebrafish Transketolase). FEBS Lett. 2003, 549, 115–122. [Google Scholar] [CrossRef]
- Rønnestad, I.; Gavaia, P.J.; Viegas, C.S.B.; Verri, T.; Romano, A.; Nilsen, T.O.; Jordal, A.-E.O.; Kamisaka, Y.; Cancela, M.L. Oligopeptide Transporter PepT1 in Atlantic Cod (Gadus Morhua L.): Cloning, Tissue Expression and Comparative Aspects. J. Exp. Biol. 2007, 210, 3883–3896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Pan, Y.; Wong, E.A.; Bloomquist, J.R.; Webb, K.E. Molecular Cloning and Functional Expression of a Chicken Intestinal Peptide Transporter (CPepT1) in Xenopus Oocytes and Chinese Hamster Ovary Cells. J. Nutr. 2002, 132, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.J.; Kanai, Y.; Nussberger, S.; Ganapathy, V.; Leibach, F.H.; Romero, M.F.; Singh, S.K.; Boron, W.F.; Hediger, M.A. Expression Cloning of a Mammalian Proton-Coupled Oligopeptide Transporter. Nature 1994, 368, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Fei, Y.J.; Prasad, P.D.; Ramamoorthy, S.; Han, H.; Yang-Feng, T.L.; Hediger, M.A.; Ganapathy, V.; Leibach, F.H. Human Intestinal H+/Peptide Cotransporter. Cloning, Functional Expression, and Chromosomal Localization. J. Biol. Chem. 1995, 270, 6456–6463. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Aliaga, I.; Daniel, H. Peptide Transporters and Their Roles in Physiological Processes and Drug Disposition. Xenobiotica 2008, 38, 1022–1042. [Google Scholar] [CrossRef]
- Brandsch, M. Drug transport via the intestinal peptide transporter PepT1. Curr. Opin. Pharmacol. 2013, 13, 881–887. [Google Scholar] [CrossRef]
- Brandsch, M.; Knütter, I.; Bosse-Doenecke, E. Pharmaceutical and pharmacological importance of peptide transporters. J. Pharm. Pharmacol. 2008, 60, 543–585. [Google Scholar] [CrossRef]
- Zhou, S.; Shu, Y. Transcriptional Regulation of Solute Carrier (SLC) Drug Transporters. Drug Metab. Dispos. 2022, 50, DMD-MR-2021-000704. [Google Scholar] [CrossRef]
- Killer, M.; Wald, J.; Pieprzkthomas, J.; Marlovits, C.; Low, C. Structural snapshots of human PepT1 and PepT2 reveal mechanistic insights into substrate and drug transport across epithelial membranes. Sci. Adv. 2021, 7, eabk3259. [Google Scholar] [CrossRef]
- Nielsen, C.U.; Brodin, B. Di/tri-peptide transporters as drug delivery targets: Regulation of transport under physiological and patho-physiological conditions. Curr. Drug Targets 2003, 4, 373–388. [Google Scholar] [CrossRef]
- Spanier, B. Transcriptional and Functional Regulation of the Intestinal Peptide Transporter PEPT1. J. Physiol. 2014, 592, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Smith, D.E.; Keep, R.F.; Brosius, F.C. Immunolocalization of the Proton-Coupled Oligopeptide Transporter PEPT2 in Developing Rat Brain. Mol. Pharm. 2004, 1, 248–256. [Google Scholar] [CrossRef]
- Teuscher, N.S.; Shen, H.; Shu, C.; Xiang, J.; Keep, R.F.; Smith, D.E. Carnosine Uptake in Rat Choroid Plexus Primary Cell Cultures and Choroid Plexus Whole Tissue from PEPT2 Null Mice. J. Neurochem. 2004, 89, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, C.; Shen, H.; Teuscher, N.S.; Lorenzi, P.J.; Keep, R.F.; Smith, D.E. Role of PEPT2 in Peptide/Mimetic Trafficking at the Blood-Cerebrospinal Fluid Barrier: Studies in Rat Choroid Plexus Epithelial Cells in Primary Culture. J. Pharmacol. Exp. Ther. 2002, 301, 820–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Distribution of Peptide Transporter PEPT2 MRNA in the Rat Nervous System—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/10221455/ (accessed on 5 May 2022).
- Török, M.; Huwyler, J.; Drewe, J.; Gutmann, H.; Fricker, G. Transport of the Beta-Lactam Antibiotic Benzylpenicillin and the Dipeptide Glycylsarcosine by Brain Capillary Endothelial Cells in Vitro. Drug Metab. Dispos. 1998, 26, 1144–1148. [Google Scholar]
- Kudo, M.; Katayoshi, T.; Kobayashi-Nakamura, K.; Akagawa, M.; Tsuji-Naito, K. H(+)/Peptide Transporter (PEPT2) Is Expressed in Human Epidermal Keratinocytes and Is Involved in Skin Oligopeptide Transport. Biochem. Biophys. Res. Commun. 2016, 475, 335–341. [Google Scholar] [CrossRef] [Green Version]
- Terada, T.; Inui, K. Recent Advances in Structural Biology of Peptide Transporters. Curr. Top. Membr. 2012, 70, 257–274. [Google Scholar] [CrossRef]
- Shen, H.; Smith, D.E.; Yang, T.; Huang, Y.G.; Schnermann, J.B.; Brosius, F.C. Localization of PEPT1 and PEPT2 Proton-Coupled Oligopeptide Transporter MRNA and Protein in Rat Kidney. Am. J. Physiol. 1999, 276, F658–F665. [Google Scholar] [CrossRef]
- Smith, D.E.; Pavlova, A.; Berger, U.V.; Hediger, M.A.; Yang, T.; Huang, Y.G.; Schnermann, J.B. Tubular Localization and Tissue Distribution of Peptide Transporters in Rat Kidney. Pharm. Res. 1998, 15, 1244–1249. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Aliaga, I.; Frey, I.; Boll, M.; Groneberg, D.A.; Eichinger, H.M.; Balling, R.; Daniel, H. Targeted Disruption of the Peptide Transporter Pept2 Gene in Mice Defines Its Physiological Role in the Kidney. Mol. Cell Biol. 2003, 23, 3247–3252. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Smith, D.E.; Keep, R.F.; Xiang, J.; Brosius, F.C. Targeted Disruption of the PEPT2 Gene Markedly Reduces Dipeptide Uptake in Choroid Plexus. J. Biol. Chem. 2003, 278, 4786–4791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, T.; Shimada, S.; Guo, W.; Sato, K.; Kohmura, E.; Hayakawa, T.; Takagi, T.; Tohyama, M. Cloning and Functional Expression of a Brain Peptide/Histidine Transporter. J. Biol. Chem. 1997, 272, 10205–10211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakata, K.; Yamashita, T.; Maeda, M.; Moriyama, Y.; Shimada, S.; Tohyama, M. Cloning of a Lymphatic Peptide/Histidine Transporter. Biochem. J. 2001, 356, 53–60. [Google Scholar] [CrossRef]
- Bhardwaj, R.K.; Herrera-Ruiz, D.; Eltoukhy, N.; Saad, M.; Knipp, G.T. The Functional Evaluation of Human Peptide/Histidine Transporter 1 (HPHT1) in Transiently Transfected COS-7 Cells. Eur. J. Pharm. Sci. 2006, 27, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Groneberg, D.A.; Fischer, A.; Chung, K.F.; Daniel, H. Molecular mechanisms of pulmonary peptidomimetic drug and peptide transport. Am. J. Respir. Cell Mol. Biol. 2004, 30, 251–260. [Google Scholar] [CrossRef]
- Boscutti, G.; Nardon, C.; Marchio, L.; Crisma, M.; Biondi, B.; Dalzoppo, D.; Dalla Via, L.; Formaggio, F.; Casini, A.; Fregona, D. Anticancer Gold (III) Peptidomimetics: From Synthesis to in vitro and ex vivo Biological Evaluations. Chem. Med. Chem. 2018, 13, 1131–1145. [Google Scholar] [CrossRef] [Green Version]
- Kouodom, M.N.; Boscutti, G.; Celegato, M.; Crisma, M.; Sitran, S.; Aldinucci, D.; Formaggio, F.; Ronconi, L.; Fregona, D. Rational design of gold (III)-dithiocarbamato peptidomimetics for the targeted anticancer chemotherapy. J. Inorg. Biochem. 2012, 117, 248–260. [Google Scholar] [CrossRef]
- Adibi, S.A. Regulation of Expression of the Intestinal Oligopeptide Transporter (Pept-1) in Health and Disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G779–G788. [Google Scholar] [CrossRef]
- Steffansen, B.; Nielsen, C.U.; Frokjaer, S. Delivery aspects of small peptides and substrates for peptide transporters. Eur. J. Pharm. Biopharm. 2005, 60, 241–245. [Google Scholar] [CrossRef]
- Viennois, E.; Pujada, A.; Zen, J.; Merlin, D. Function, Regulation, and Pathophysiological Relevance of the POT Superfamily, Specifically PepT1 in Inflammatory Bowel Disease. Compr. Physiol. 2018, 8, 731–760. [Google Scholar] [CrossRef]
- Agu, R.; Cowley, E.; Shao, D.; Macdonald, C.; Kirkpatrick, D.; Renton, K.; Massoud, E. Proton-Coupled Oligopeptide Transporter (POT) Family Expression in Human Nasal Epithelium and Their Drug Transport Potential. Mol. Pharm. 2011, 8, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Ayyadurai, S.; Charania, M.A.; Xiao, B.; Viennois, E.; Merlin, D. PepT1 Expressed in Immune Cells Has an Important Role in Promoting the Immune Response during Experimentally Induced Colitis. Lab. Investig. 2013, 93, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Merlin, D.; Si-Tahar, M.; Sitaraman, S.V.; Eastburn, K.; Williams, I.; Liu, X.; Hediger, M.A.; Madara, J.L. Colonic Epithelial HPepT1 Expression Occurs in Inflammatory Bowel Disease: Transport of Bacterial Peptides Influences Expression of MHC Class 1 Molecules. Gastroenterology 2001, 120, 1666–1679. [Google Scholar] [CrossRef] [PubMed]
- Viennois, E.; Ingersoll, S.A.; Ayyadurai, S.; Zhao, Y.; Wang, L.; Zhang, M.; Han, M.K.; Garg, P.; Xiao, B.; Merlin, D. Critical Role of PepT1 in Promoting Colitis-Associated Cancer and Therapeutic Benefits of the Anti-Inflammatory PepT1-Mediated Tripeptide KPV in a Murine Model. Cell Mol. Gastroenterol. Hepatol. 2016, 2, 340–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alghamdi, O.; King, N.; Jones, G.L.; Moens, P.D.J. Effect of Ageing and Hypertension on the Expression and Activity of PEPT2 in Normal and Hypertrophic Hearts. Amino Acids 2021, 53, 183–193. [Google Scholar] [CrossRef]
- Landowski, C.P.; Vig, B.S.; Song, X.; Amidon, G.L. Targeted Delivery to PEPT1-Overexpressing Cells: Acidic, Basic, and Secondary Floxuridine Amino Acid Ester Prodrugs. Mol. Cancer Ther. 2005, 4, 659–667. [Google Scholar] [CrossRef] [Green Version]
- Tai, W.; Chen, Z.; Cheng, K. Expression Profile and Functional Activity of Peptide Transporters in Prostate Cancer Cells. Mol. Pharm. 2013, 10, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Wu, X.; Wang, T.; Zhao, J.; Liu, X.; Yao, Z.; Zhang, Q.; Jian, X. Targeting PEPT1: A Novel Strategy to Improve the Antitumor Efficacy of Doxorubicin in Human Hepatocellular Carcinoma Therapy. Oncotarget 2017, 8, 40454–40468. [Google Scholar] [CrossRef] [Green Version]
- Neudeck, B.L.; Foster, D.R.; Li, L.Y.; Gonzales, J.P.; Welage, L.S. The Effects of Thermal Injury on Transcellular Permeability and Intestinal P-Glycoprotein in Rats. Burns 2003, 29, 803–809. [Google Scholar] [CrossRef]
- Hartmann, G.; Kim, H.; Piquette-Miller, M. Regulation of the Hepatic Multidrug Resistance Gene Expression by Endotoxin and Inflammatory Cytokines in Mice. Int. Immunopharmacol. 2001, 1, 189–199. [Google Scholar] [CrossRef]
- Kalitsky-Szirtes, J.; Shayeganpour, A.; Brocks, D.R.; Piquette-Miller, M. Suppression of Drug-Metabolizing Enzymes and Efflux Transporters in the Intestine of Endotoxin-Treated Rats. Drug Metab. Dispos. 2004, 32, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, D.R.; Landowski, C.P.; Zheng, X.; Amidon, G.L.; Welage, L.S. Interferon-Gamma Increases Expression of the Di/Tri-Peptide Transporter, h-PEPT1, and Dipeptide Transport in Cultured Human Intestinal Monolayers. Pharmacol. Res. 2009, 59, 215–220. [Google Scholar] [CrossRef]
- Foster, D.R.; Zheng, X. Cephalexin Inhibits N-Formylated Peptide Transport and Intestinal Hyperpermeability in CaCO2 Cells. J. Pharm. Pharm. Sci. 2007, 10, 299–310. [Google Scholar] [PubMed]
- Walker, D.; Thwaites, D.T.; Simmons, N.L.; Gilbert, H.J.; Hirst, B.H. Substrate Upregulation of the Human Small Intestinal Peptide Transporter, HPepT1. J. Physiol. 1998, 507, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Shiraga, T.; Miyamoto, K.; Tanaka, H.; Yamamoto, H.; Taketani, Y.; Morita, K.; Tamai, I.; Tsuji, A.; Takeda, E. Cellular and Molecular Mechanisms of Dietary Regulation on Rat Intestinal H+/Peptide Transporter PepT1. Gastroenterology 1999, 116, 354–362. [Google Scholar] [CrossRef]
- Erickson, R.H.; Gum, J.R.; Lindstrom, M.M.; McKean, D.; Kim, Y.S. Regional Expression and Dietary Regulation of Rat Small Intestinal Peptide and Amino Acid Transporter MRNAs. Biochem. Biophys. Res. Commun. 1995, 216, 249–257. [Google Scholar] [CrossRef]
- Guerrini, L.; Gong, S.S.; Mangasarian, K.; Basilico, C. Cis- and Trans-Acting Elements Involved in Amino Acid Regulation of Asparagine Synthetase Gene Expression. Mol. Cell Biol. 1993, 13, 3202–3212. [Google Scholar]
- Lochs, H.; Hübl, W.; Gasic, S.; Roth, E.; Morse, E.L.; Adibi, S.A. Glycylglutamine: Metabolism and Effects on Organ Balances of Amino Acids in Postabsorptive and Starved Subjects. Am. J. Physiol. 1992, 262, E155–E160. [Google Scholar] [CrossRef]
- Adibi, S.A.; Lochs, H.; Abumrad, N.N.; Daniel, H.; Vazquez, J.A. Removal of Glycylglutamine from Plasma by Individual Tissues: Mechanism and Impact on Amino Acid Fluxes in Postabsorption and Starvation. J. Nutr. 1993, 123, 325–331. [Google Scholar] [CrossRef]
- Perry, J.R.; Basrai, M.A.; Steiner, H.Y.; Naider, F.; Becker, J.M. Isolation and Characterization of a Saccharomyces Cerevisiae Peptide Transport Gene. Mol. Cell Biol. 1994, 14, 104–115. [Google Scholar]
- Turner, G.C.; Du, F.; Varshavsky, A. Peptides Accelerate Their Uptake by Activating a Ubiquitin-Dependent Proteolytic Pathway. Nature 2000, 405, 579–583. [Google Scholar] [CrossRef]
- Novotny, A.; Xiang, J.; Stummer, W.; Teuscher, N.S.; Smith, D.E.; Keep, R.F. Mechanisms of 5-Aminolevulinic Acid Uptake at the Choroid Plexus. J. Neurochem. 2000, 75, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Teuscher, N.S.; Keep, R.F.; Smith, D.E. PEPT2-Mediated Uptake of Neuropeptides in Rat Choroid Plexus. Pharm. Res. 2001, 18, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Tattoli, I.; Wojtal, K.A.; Vavricka, S.R.; Philpott, D.J.; Girardin, S.E. PH-Dependent Internalization of Muramyl Peptides from Early Endosomes Enables Nod1 and Nod2 Signaling. J. Biol. Chem. 2009, 284, 23818–23829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irie, M.; Terada, T.; Katsura, T.; Matsuoka, S.; Inui, K. Computational Modelling of H+-Coupled Peptide Transport via Human PEPT1. J. Physiol. 2005, 565, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, J.P.; Brayden, D.J.; Ryan, S.M. Evaluation of PepT1 Transport of Food-Derived Antihypertensive Peptides, Ile-Pro-Pro and Leu-Lys-Pro Using in Vitro, Ex Vivo and in Vivo Transport Models. Eur. J. Pharm. Biopharm. 2017, 115, 276–284. [Google Scholar] [CrossRef] [Green Version]
- Abousaab, A.; Warsi, J.; Salker, M.S.; Lang, F. β-Klotho as a Negative Regulator of the Peptide Transporters PEPT1 and PEPT2. Cell Physiol. Biochem. 2016, 40, 874–882. [Google Scholar] [CrossRef] [Green Version]
- Chang, Q.; Hoefs, S.; van der Kemp, A.W.; Topala, C.N.; Bindels, R.J.; Hoenderop, J.G. The Beta-Glucuronidase Klotho Hydrolyzes and Activates the TRPV5 Channel. Science 2005, 310, 490–493. [Google Scholar] [CrossRef]
- Warsi, J.; Abousaab, A.; Lang, F. Up-Regulation of Excitatory Amino Acid Transporters EAAT1 and EAAT2 by ß-Klotho. Neurosignals 2015, 23, 59–70. [Google Scholar] [CrossRef]
- Neumann, J.; Bruch, M.; Gebauer, S.; Brandsch, M. Transport of the Phosphonodipeptide Alafosfalin by the H+/Peptide Cotransporters PEPT1 and PEPT2 in Intestinal and Renal Epithelial Cells. Eur. J. Biochem. 2004, 271, 2012–2017. [Google Scholar] [CrossRef]
- He, Y.-Y.; Li, T.-T.; Chen, J.-X.; She, X.-X.; Ren, D.-F.; Lu, J. Transport of ACE Inhibitory Peptides Ile-Gln-Pro and Val-Glu-Pro Derived from Spirulina Platensis Across Caco-2 Monolayers. J. Food Sci. 2018, 83, 2586–2592. [Google Scholar] [CrossRef] [PubMed]
- Vij, R.; Reddi, S.; Kapila, S.; Kapila, R. Transepithelial Transport of Milk Derived Bioactive Peptide VLPVPQK. Food Chem. 2016, 190, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Oxender, D.L.; Christensen, H.N. Distinct mediating systems for the transport of neutral amino acids by the ehrlich cell. J. Biol. Chem. 1963, 238, 3686–3699. [Google Scholar] [CrossRef]
- Bodoy, S.; Martín, L.; Zorzano, A.; Palacín, M.; Estévez, R.; Bertran, J. Identification of LAT4, a Novel Amino Acid Transporter with System L Activity *. J. Biol. Chem. 2005, 280, 12002–12011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastroberardino, L.; Spindler, B.; Pfeiffer, R.; Skelly, P.J.; Loffing, J.; Shoemaker, C.B.; Verrey, F. Amino-Acid Transport by Heterodimers of 4F2hc/CD98 and Members of a Permease Family. Nature 1998, 395, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Pineda, M.; Fernández, E.; Torrents, D.; Estévez, R.; López, C.; Camps, M.; Lloberas, J.; Zorzano, A.; Palacín, M. Identification of a Membrane Protein, LAT-2, That Co-Expresses with 4F2 Heavy Chain, an L-Type Amino Acid Transport Activity with Broad Specificity for Small and Large Zwitterionic Amino Acids. J. Biol. Chem. 1999, 274, 19738–19744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, P.D.; Wang, H.; Huang, W.; Kekuda, R.; Rajan, D.P.; Leibach, F.H.; Ganapathy, V. Human LAT1, a Subunit of System L Amino Acid Transporter: Molecular Cloning and Transport Function. Biochem. Biophys. Res. Commun. 1999, 255, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Kanai, Y.; Endou, H. Heterodimeric Amino Acid Transporters: Molecular Biology and Pathological and Pharmacological Relevance. Curr. Drug Metab. 2001, 2, 339–354. [Google Scholar] [CrossRef]
- Yanagida, O.; Kanai, Y.; Chairoungdua, A.; Kim, D.K.; Segawa, H.; Nii, T.; Cha, S.H.; Matsuo, H.; Fukushima, J.; Fukasawa, Y.; et al. Human L-Type Amino Acid Transporter 1 (LAT1): Characterization of Function and Expression in Tumor Cell Lines. Biochim. Biophys. Acta 2001, 1514, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Segawa, H.; Fukasawa, Y.; Miyamoto, K.; Takeda, E.; Endou, H.; Kanai, Y. Identification and Functional Characterization of a Na+-Independent Neutral Amino Acid Transporter with Broad Substrate Selectivity. J. Biol. Chem. 1999, 274, 19745–19751. [Google Scholar] [CrossRef] [Green Version]
- Uchino, H.; Kanai, Y.; Kim, D.K.; Wempe, M.F.; Chairoungdua, A.; Morimoto, E.; Anders, M.W.; Endou, H. Transport of Amino Acid-Related Compounds Mediated by L-Type Amino Acid Transporter 1 (LAT1): Insights Into the Mechanisms of Substrate Recognition. Mol. Pharmacol. 2002, 61, 729–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanai, Y.; Endou, H. Functional Properties of Multispecific Amino Acid Transporters and Their Implications to Transporter-Mediated Toxicity. J. Toxicol. Sci. 2003, 28, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Whorton, A.R. Identification of Stereoselective Transporters for S-Nitroso-L-Cysteine. J. Biol. Chem. 2005, 280, 20102–20110. [Google Scholar] [CrossRef] [Green Version]
- Su, T.Z.; Feng, M.R.; Weber, M.L. Mediation of Highly Concentrative Uptake of Pregabalin by L-Type Amino Acid Transport in Chinese Hamster Ovary and Caco-2 Cells. J. Pharmacol. Exp. Ther. 2005, 313, 1406–1415. [Google Scholar] [CrossRef] [Green Version]
- Duelli, R.; Enerson, B.E.; Gerhart, D.Z.; Drewes, L.R. Expression of Large Amino Acid Transporter LAT1 in Rat Brain Endothelium. J. Cereb. Blood Flow Metab. 2000, 20, 1557–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudo, Y.; Boyd, C.A.R. Characterisation of L-Tryptophan Transporters in Human Placenta: A Comparison of Brush Border and Basal Membrane Vesicles. J. Physiol. 2001, 531, 405–416. [Google Scholar] [CrossRef]
- Verrey, F. System L: Heteromeric Exchangers of Large, Neutral Amino Acids Involved in Directional Transport. Pflug. Arch. 2003, 445, 529–533. [Google Scholar] [CrossRef]
- Franca, R.; Veljkovic, E.; Walter, S.; Wagner, C.A.; Verrey, F. Heterodimeric Amino Acid Transporter Glycoprotein Domains Determining Functional Subunit Association. Biochem. J. 2005, 388, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Fraga, S.; Pinho, M.J.; Soares-da-Silva, P. Expression of LAT1 and LAT2 Amino Acid Transporters in Human and Rat Intestinal Epithelial Cells. Amino Acids 2005, 29, 229–233. [Google Scholar] [CrossRef]
- Dave, M.H.; Schulz, N.; Zecevic, M.; Wagner, C.A.; Verrey, F. Expression of Heteromeric Amino Acid Transporters along the Murine Intestine. J. Physiol. 2004, 558, 597–610. [Google Scholar] [CrossRef]
- Killian, D.M.; Chikhale, P.J. Predominant Functional Activity of the Large, Neutral Amino Acid Transporter (LAT1) Isoform at the Cerebrovasculature. Neurosci. Lett. 2001, 306, 1–4. [Google Scholar] [CrossRef]
- Soares-Da-Silva, P.; Serrão, M.P.; Pinho, M.J.; Bonifácio, M.J. Cloning and gene silencing of LAT2, the L-3, 4-dihydroxyphenylalanine (L-DOPA) transporter, in pig renal LLC-PK1 epithelial cells. FASEB J. 2004, 18, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.A.; Lang, F.; Bröer, S. Function and Structure of Heterodimeric Amino Acid Transporters. Am. J. Physiol. Cell Physiol. 2001, 281, C1077–C1093. [Google Scholar] [CrossRef] [PubMed]
- Asano, S.; Kameyama, M.; Oura, A.; Morisato, A.; Sakai, H.; Tabuchi, Y.; Chairoungdua, A.; Endou, H.; Kanai, Y. L-Type Amino Acid Transporter-1 Expressed in Human Astrocytomas, U343MGa. Biol. Pharm. Bull. 2007, 30, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Cibrian, D.; Saiz, M.L.; de la Fuente, H.; Sánchez-Díaz, R.; Moreno-Gonzalo, O.; Jorge, I.; Ferrarini, A.; Vázquez, J.; Punzón, C.; Fresno, M.; et al. CD69 Controls the Uptake of L-Tryptophan through LAT1-CD98 and AhR-Dependent Secretion of IL-22 in Psoriasis. Nat. Immunol. 2016, 17, 985–996. [Google Scholar] [CrossRef] [Green Version]
- Veldhoen, M.; Hirota, K.; Christensen, J.; O’Garra, A.; Stockinger, B. Natural Agonists for Aryl Hydrocarbon Receptor in Culture Medium Are Essential for Optimal Differentiation of Th17 T Cells. J. Exp. Med. 2009, 206, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Smirnova, A.; Wincent, E.; Vikström Bergander, L.; Alsberg, T.; Bergman, J.; Rannug, A.; Rannug, U. Evidence for New Light-Independent Pathways for Generation of the Endogenous Aryl Hydrocarbon Receptor Agonist FICZ. Chem. Res. Toxicol. 2016, 29, 75–86. [Google Scholar] [CrossRef]
- Cibrian, D.; Castillo-González, R.; Fernández-Gallego, N.; de la Fuente, H.; Jorge, I.; Saiz, M.L.; Punzón, C.; Ramírez-Huesca, M.; Vicente-Manzanares, M.; Fresno, M.; et al. Targeting L-Type Amino Acid Transporter 1 in Innate and Adaptive T Cells Efficiently Controls Skin Inflammation. J. Allergy Clin. Immunol. 2020, 145, 199–214. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, B.C.; Bode, B.P. Amino Acid Transporters ASCT2 and LAT1 in Cancer: Partners in Crime? Semin. Cancer Biol. 2005, 15, 254–266. [Google Scholar] [CrossRef]
- Wang, Q.; Holst, J. L-Type Amino Acid Transport and Cancer: Targeting the MTORC1 Pathway to Inhibit Neoplasia. Am. J. Cancer Res. 2015, 5, 1281–1294. [Google Scholar]
- Furuya, M.; Horiguchi, J.; Nakajima, H.; Kanai, Y.; Oyama, T. Correlation of L-Type Amino Acid Transporter 1 and CD98 Expression with Triple Negative Breast Cancer Prognosis. Cancer Sci. 2012, 103, 382–389. [Google Scholar] [CrossRef] [PubMed]
- El Ansari, R.; Craze, M.L.; Miligy, I.; Diez-Rodriguez, M.; Nolan, C.C.; Ellis, I.O.; Rakha, E.A.; Green, A.R. The Amino Acid Transporter SLC7A5 Confers a Poor Prognosis in the Highly Proliferative Breast Cancer Subtypes and Is a Key Therapeutic Target in Luminal B Tumours. Breast Cancer Res. 2018, 20, 21. [Google Scholar] [CrossRef] [PubMed]
- Maimaiti, M.; Sakamoto, S.; Yamada, Y.; Sugiura, M.; Rii, J.; Takeuchi, N.; Imamura, Y.; Furihata, T.; Ando, K.; Higuchi, K.; et al. Expression of L-Type Amino Acid Transporter 1 as a Molecular Target for Prognostic and Therapeutic Indicators in Bladder Carcinoma. Sci. Rep. 2020, 10, 1292. [Google Scholar] [CrossRef] [PubMed]
- Kaira, K.; Sunose, Y.; Arakawa, K.; Ogawa, T.; Sunaga, N.; Shimizu, K.; Tominaga, H.; Oriuchi, N.; Itoh, H.; Nagamori, S.; et al. Prognostic Significance of L-Type Amino-Acid Transporter 1 Expression in Surgically Resected Pancreatic Cancer. Br. J. Cancer 2012, 107, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Kaira, K.; Sunose, Y.; Ohshima, Y.; Ishioka, N.S.; Arakawa, K.; Ogawa, T.; Sunaga, N.; Shimizu, K.; Tominaga, H.; Oriuchi, N.; et al. Clinical Significance of L-Type Amino Acid Transporter 1 Expression as a Prognostic Marker and Potential of New Targeting Therapy in Biliary Tract Cancer. BMC Cancer 2013, 13, 482. [Google Scholar] [CrossRef] [Green Version]
- Oda, K.; Hosoda, N.; Endo, H.; Saito, K.; Tsujihara, K.; Yamamura, M.; Sakata, T.; Anzai, N.; Wempe, M.F.; Kanai, Y.; et al. L-Type Amino Acid Transporter 1 Inhibitors Inhibit Tumor Cell Growth. Cancer Sci. 2010, 101, 173–179. [Google Scholar] [CrossRef]
- Napolitano, L.; Scalise, M.; Koyioni, M.; Koutentis, P.; Catto, M.; Eberini, I.; Parravicini, C.; Palazzolo, L.; Pisani, L.; Galluccio, M.; et al. Potent Inhibitors of Human LAT1 (SLC7A5) Transporter Based on Dithiazole and Dithiazine Compounds for Development of Anticancer Drugs. Biochem. Pharmacol. 2017, 143, 39–52. [Google Scholar] [CrossRef]
- Kongpracha, P.; Nagamori, S.; Wiriyasermkul, P.; Tanaka, Y.; Kaneda, K.; Okuda, S.; Ohgaki, R.; Kanai, Y. Structure-Activity Relationship of a Novel Series of Inhibitors for Cancer Type Transporter L-Type Amino Acid Transporter 1 (LAT1). J. Pharmacol. Sci. 2017, 133, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Li, L.; Coyaud, E.; Luna, A.; Sander, C.; Raught, B.; Asara, J.M.; Brown, M.; Muthuswamy, S.K. LLGL2 Rescues Nutrient Stress by Promoting Leucine Uptake in ER+ Breast Cancer. Nature 2019, 569, 275–279. [Google Scholar] [CrossRef]
- Häfliger, P.; Graff, J.; Rubin, M.; Stooss, A.; Dettmer, M.S.; Altmann, K.-H.; Gertsch, J.; Charles, R.-P. The LAT1 Inhibitor JPH203 Reduces Growth of Thyroid Carcinoma in a Fully Immunocompetent Mouse Model. J. Exp. Clin. Cancer Res. 2018, 37, 234. [Google Scholar] [CrossRef]
- Kume, E.; Mutou, T.; Kansaku, N.; Takahashi, H.; Wempe, M.F.; Ikegami, M.; Kanai, Y.; Endou, H.; Wakui, S. Ultrastructural Immunohistochemical Study of L-Type Amino Acid Transporter 1-4F2 Heavy Chain in Tumor Microvasculatures of N-Butyl-N-(4-Hydroxybutyl) Nitrosamine (BBN) Induced Rat Bladder Carcinoma. Microscopy 2017, 66, 198–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haining, Z.; Kawai, N.; Miyake, K.; Okada, M.; Okubo, S.; Zhang, X.; Fei, Z.; Tamiya, T. Relation of LAT1/4F2hc Expression with Pathological Grade, Proliferation and Angiogenesis in Human Gliomas. BMC Clin. Pathol. 2012, 12, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, L.; Ohgaki, R.; Hara, S.; Okuda, S.; Wei, L.; Okanishi, H.; Nagamori, S.; Endou, H.; Kanai, Y. Amino Acid Transporter LAT1 in Tumor-Associated Vascular Endothelium Promotes Angiogenesis by Regulating Cell Proliferation and VEGF-A-Dependent MTORC1 Activation. J. Exp. Clin. Cancer Res. 2020, 39, 266. [Google Scholar] [CrossRef] [PubMed]
- Lu, X. The Role of Large Neutral Amino Acid Transporter (LAT1) in Cancer. Curr. Cancer Drug Targets 2019, 19, 863–876. [Google Scholar] [CrossRef]
- Vumma, R.; Wiesel, F.-A.; Flyckt, L.; Bjerkenstedt, L.; Venizelos, N. Functional Characterization of Tyrosine Transport in Fibroblast Cells from Healthy Controls. Neurosci. Lett. 2008, 434, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, A.; Gautam, S.; Hyeon, S.J.; Ryu, H.; Kang, Y.-S. L-Citrulline Level and Transporter Activity Are Altered in Experimental Models of Amyotrophic Lateral Sclerosis. Mol. Neurobiol. 2021, 58, 647–657. [Google Scholar] [CrossRef]
- Huttunen, J.; Peltokangas, S.; Gynther, M.; Natunen, T.; Hiltunen, M.; Auriola, S.; Ruponen, M.; Vellonen, K.-S.; Huttunen, K.M. L-Type Amino Acid Transporter 1 (LAT1/Lat1)-Utilizing Prodrugs Can Improve the Delivery of Drugs into Neurons, Astrocytes and Microglia. Sci. Rep. 2019, 9, 12860. [Google Scholar] [CrossRef] [Green Version]
- Knöpfel, E.B.; Vilches, C.; Camargo, S.M.R.; Errasti-Murugarren, E.; Stäubli, A.; Mayayo, C.; Munier, F.L.; Miroshnikova, N.; Poncet, N.; Junza, A.; et al. Dysfunctional LAT2 Amino Acid Transporter Is Associated With Cataract in Mouse and Humans. Front. Physiol. 2019, 10, 688. [Google Scholar] [CrossRef] [Green Version]
- Poncet, N.; Mitchell, F.E.; Ibrahim, A.F.M.; McGuire, V.A.; English, G.; Arthur, J.S.C.; Shi, Y.-B.; Taylor, P.M. The Catalytic Subunit of the System L1 Amino Acid Transporter (Slc7a5) Facilitates Nutrient Signalling in Mouse Skeletal Muscle. PLoS ONE 2014, 9, e89547. [Google Scholar] [CrossRef] [Green Version]
- Markowicz-Piasecka, M.; Huttunen, J.; Montaser, A.; Huttunen, K.M. Hemocompatible LAT1-Inhibitor Can Induce Apoptosis in Cancer Cells without Affecting Brain Amino Acid Homeostasis. Apoptosis 2020, 25, 426–440. [Google Scholar] [CrossRef]
- Higuchi, K.; Sakamoto, S.; Ando, K.; Maimaiti, M.; Takeshita, N.; Okunushi, K.; Reien, Y.; Imamura, Y.; Sazuka, T.; Nakamura, K.; et al. Characterization of the Expression of LAT1 as a Prognostic Indicator and a Therapeutic Target in Renal Cell Carcinoma. Sci. Rep. 2019, 9, 16776. [Google Scholar] [CrossRef] [PubMed]
- Hansson, S.R.; Nääv, Å.; Erlandsson, L. Oxidative Stress in Preeclampsia and the Role of Free Fetal Hemoglobin. Front. Physiol. 2014, 5, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rani, N.; Dhingra, R.; Arya, D.S.; Kalaivani, M.; Bhatla, N.; Kumar, R. Role of Oxidative Stress Markers and Antioxidants in the Placenta of Preeclamptic Patients. J. Obstet. Gynaecol. Res. 2010, 36, 1189–1194. [Google Scholar] [CrossRef]
- Aiko, Y.; Askew, D.J.; Aramaki, S.; Myoga, M.; Tomonaga, C.; Hachisuga, T.; Suga, R.; Kawamoto, T.; Tsuji, M.; Shibata, E. Differential Levels of Amino Acid Transporters System L and ASCT2, and the MTOR Protein in Placenta of Preeclampsia and IUGR. BMC Pregnancy Childbirth 2014, 14, 181. [Google Scholar] [CrossRef] [Green Version]
- Myatt, L. Reactive Oxygen and Nitrogen Species and Functional Adaptation of the Placenta. Placenta 2010, 31, S66–S69. [Google Scholar] [CrossRef] [Green Version]
- Straka, E.; Ellinger, I.; Balthasar, C.; Scheinast, M.; Schatz, J.; Szattler, T.; Bleichert, S.; Saleh, L.; Knöfler, M.; Zeisler, H.; et al. Mercury Toxicokinetics of the Healthy Human Term Placenta Involve Amino Acid Transporters and ABC Transporters. Toxicology 2016, 340, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Luo, W.; Tan, D.; Liang, H.; Zhang, Q.; Tian, N.; Cao, K.; Tan, Y.; Ma, J.; Han, R. Positive Regulation of Placentation by L-Amino Acid Transporter-1 (Lat1) in Pregnant Mice. Int. J. Clin. Exp. Pathol. 2017, 10, 9551–9558. [Google Scholar] [PubMed]
- Ohgaki, R.; Ohmori, T.; Hara, S.; Nakagomi, S.; Kanai-Azuma, M.; Kaneda-Nakashima, K.; Okuda, S.; Nagamori, S.; Kanai, Y. Essential Roles of L-Type Amino Acid Transporter 1 in Syncytiotrophoblast Development by Presenting Fusogenic 4F2hc. Mol. Cell Biol. 2017, 37, e00427-16. [Google Scholar] [CrossRef] [Green Version]
- Jungnickel, K.E.J.; Parker, J.L.; Newstead, S. Structural Basis for Amino Acid Transport by the CAT Family of SLC7 Transporters. Nat. Commun. 2018, 9, 550. [Google Scholar] [CrossRef] [Green Version]
- Yan, R.; Zhao, X.; Lei, J.; Zhou, Q. Structure of the Human LAT1-4F2hc Heteromeric Amino Acid Transporter Complex. Nature 2019, 568, 127–130. [Google Scholar] [CrossRef]
- Singh, N.; Scalise, M.; Galluccio, M.; Wieder, M.; Seidel, T.; Langer, T.; Indiveri, C.; Ecker, G.F. Discovery of Potent Inhibitors for the Large Neutral Amino Acid Transporter 1 (LAT1) by Structure-Based Methods. Int. J. Mol. Sci. 2018, 20, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagamori, S.; Wiriyasermkul, P.; Okuda, S.; Kojima, N.; Hari, Y.; Kiyonaka, S.; Mori, Y.; Tominaga, H.; Ohgaki, R.; Kanai, Y. Structure-Activity Relations of Leucine Derivatives Reveal Critical Moieties for Cellular Uptake and Activation of MTORC1-Mediated Signaling. Amino Acids 2016, 48, 1045–1058. [Google Scholar] [CrossRef] [PubMed]
- Ylikangas, H.; Malmioja, K.; Peura, L.; Gynther, M.; Nwachukwu, E.O.; Leppänen, J.; Laine, K.; Rautio, J.; Lahtela-Kakkonen, M.; Huttunen, K.M.; et al. Quantitative Insight into the Design of Compounds Recognized by the L-Type Amino Acid Transporter 1 (LAT1). Chem. Med. Chem. 2014, 9, 2699–2707. [Google Scholar] [CrossRef] [PubMed]
- Zur, A.A.; Chien, H.-C.; Augustyn, E.; Flint, A.; Heeren, N.; Finke, K.; Hernandez, C.; Hansen, L.; Miller, S.; Lin, L.; et al. LAT1 Activity of Carboxylic Acid Bioisosteres: Evaluation of Hydroxamic Acids as Substrates. Bioorg. Med. Chem. Lett. 2016, 26, 5000–5006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ylikangas, H.; Peura, L.; Malmioja, K.; Leppänen, J.; Laine, K.; Poso, A.; Lahtela-Kakkonen, M.; Rautio, J. Structure-Activity Relationship Study of Compounds Binding to Large Amino Acid Transporter 1 (LAT1) Based on Pharmacophore Modeling and in Situ Rat Brain Perfusion. Eur. J. Pharm. Sci. 2013, 48, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Yunger, L.M.; Cramer, R.D. Measurement of Correlation of Partition Coefficients of Polar Amino Acids. Mol. Pharmacol. 1981, 20, 602–608. [Google Scholar]
- Chien, H.-C.; Colas, C.; Finke, K.; Springer, S.; Stoner, L.; Zur, A.A.; Venteicher, B.; Campbell, J.; Hall, C.; Flint, A.; et al. Reevaluating the Substrate Specificity of the L-Type Amino Acid Transporter (LAT1). J. Med. Chem. 2018, 61, 7358–7373. [Google Scholar] [CrossRef]
- Lee, Y.; Wiriyasermkul, P.; Jin, C.; Quan, L.; Ohgaki, R.; Okuda, S.; Kusakizako, T.; Nishizawa, T.; Oda, K.; Ishitani, R.; et al. Cryo-EM structure of the human L-type amino acid transporter 1 in complex with glycoprotein CD98hc. Nat. Struct. Mol. Biol. 2019, 26, 510–517. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Lin’kova, N.S.; Dudkov, A.V.; Polyakova, V.O.; Kvetnoi, I.M. Peptidergic Regulation of Expression of Genes Encoding Antioxidant and Anti-Inflammatory Proteins. Bull. Exp. Biol. Med. 2012, 152, 615–618. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Grigoriev, E.I.; Malinin, V.V.; Ryzhak, G.A. Peptide Substance Revealing a Stress Protective Effect, Pharmaceutical Composition on Its Base and the Method of Its Application. U.S. Patent 8,071,556, 6 December 2011. [Google Scholar]
- Khavinson, V.; Linkova, N.; Kozhevnikova, E.; Trofimova, S. EDR Peptide: Possible Mechanism of Gene Expression and Protein Synthesis Regulation Involved in the Pathogenesis of Alzheimer’s Disease. Molecules 2020, 26, 159. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Ryzhak, G.A.; Grigoriev, E.I.; Ryadnova, I.Y. Peptide Substance Restoring Myocardium Function. U.S. Patent 7,662,789, 16 February 2010. [Google Scholar]
- Zamorskii, I.I.; Shchudrova, T.S.; Lin’kova, N.S.; Nichik, T.E.; Khavinson, V.K. Nephroprotective Effect of EDL Peptide at Acute Injury of Kidneys of Different Genesis. Bull. Exp. Biol. Med. 2017, 163, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.K.; Grigoriev, E.I.; Malinin, V.V.; Ryzhak, G.A. Peptide Stimulating Regeneration of Liver Tissue, Pharmaceutical Composition Based on Thereof and Method for Its Using. WO2007139430, 4 December 2006. [Google Scholar]
- Khavinson, V.K.; Grigoriev, E.I.; Malinin, V.V.; Ryzhak, G.A. Peptide Normalizing Netabolism in Bone and Cartilaginous Tissues, Pharmaceutical Composition Based Thereon and Method for Use Thereof. RU2299741C1, 27 May 2008. [Google Scholar]
- Fridman, N.V.; Linkova, N.S.; Kozhevnikova, E.O.; Gutop, E.O.; Khavinson, V.K. Comparison of the Effects of KE and AED Peptides on Functional Activity of Human Skin Fibroblasts during Their Replicative Aging. Bull. Exp. Biol. Med. 2020, 170, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Stasenko, A.A.; Zhulaĭ, V.V.; Novopol’tseva, I.I.; Dontsova, L.S.; Zaiats, N.V.; Negievich, V.I. Sensitivity of patients to immunomodulators in immunodeficiency diseases. Klin. Khirurhiia 2014, 8, 52–54. [Google Scholar]
- Khavinson, V.; Linkova, N.; Diatlova, A.; Trofimova, S. Peptide Regulation of Cell Differentiation. Stem. Cell Rev. Rep. 2020, 16, 118–125. [Google Scholar] [CrossRef]
- Kuznik, B.I.; Khasanova, N.B.; Ryzhak, G.A.; Mezsheriakova, I.E.; Khavinson, V.K. The influence of polypeptide liver complex and tetrapeptide KEDA on organism physiological function in norm and age-related pathology. Adv. Gerontol. 2020, 33, 159–164. [Google Scholar]
- Ivko, O.M.; Linkova, N.S.; Ilina, A.R.; Sharova, A.A.; Ryzhak, G.A. AEDG peptide regulates human circadian rhythms genes expression during pineal gland accelerated aging. Adv. Gerontol. 2020, 33, 429–435. [Google Scholar]
- Khavinson, V.; Diomede, F.; Mironova, E.; Linkova, N.; Trofimova, S.; Trubiani, O.; Caputi, S.; Sinjari, B. AEDG Peptide (Epitalon) Stimulates Gene Expression and Protein Synthesis during Neurogenesis: Possible Epigenetic Mechanism. Molecules 2020, 25, 609. [Google Scholar] [CrossRef] [Green Version]
- Khavinson, V.K.; Grigoriev, E.I.; Malinin, V.V.; Ryzhak, G.A. Peptide Substance Revealing an Immunogeroprotective Effect, Pharmaceutical Composition on Its Base and the Method of Its Application. U.S. Patent 8,057,810, 15 November 2011. [Google Scholar]
- Kozlov, K.L.; Bolotov, I.I.; Linkova, N.S.; Drobintseva, A.O.; Khavinson, V.K.; Dyakonov, M.M.; Kozina, L.S. Molecular aspects of vasoprotective peptide KED activity during atherosclerosis and restenosis. Adv. Gerontol. 2016, 29, 646–650. [Google Scholar]
- Khavinson, V.K.; Lin’kova, N.S.; Umnov, R.S. Peptide KED: Molecular-Genetic Aspects of Neurogenesis Regulation in Alzheimer’s Disease. Bull. Exp. Biol. Med. 2021, 171, 190–193. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Malinin, V.V.; Grigoriev, E.I. Tetrapeptide Regulating Prostate Functions and Its Compositions and Methods. European Patent EP1353939, 25 August 2004. [Google Scholar]
- Khavinson, V.K.; Pronyaeva, V.E.; Linkova, N.S.; Trofimova, S.V. Peptidegic Regulation of Differentiation of Embryonic Retinal Cells. Bull. Exp. Biol. Med. 2013, 155, 172–174. [Google Scholar] [CrossRef]
- Yu, C.-Y.; Huang, W.; Li, Z.-P.; Lei, X.-Y.; He, D.-X.; Sun, L. Progress in Self-Assembling Peptide-Based Nanomaterials for Biomedical Applications. Curr. Top. Med. Chem. 2016, 16, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zheng, C.; Webster, T.J. Self-Assembled Peptide Nanomaterials for Biomedical Applications: Promises and Pitfalls. Int. J. Nanomed. 2016, 12, 73–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galdiero, S.; Gomes, P.A.C. Peptide-Based Drugs and Drug Delivery Systems. Molecules 2017, 22, 2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foltz, M.; van der Pijl, P.C.; Duchateau, G.S. Current in Vitro Testing of Bioactive Peptides Is Not Valuable. J. Nutr. 2010, 140, 117–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, T. Are Peptides Absorbable Compounds? J. Agric. Food Chem. 2018, 66, 393–394. [Google Scholar] [CrossRef]
- Foltz, M.; van Buren, L.; Klaffke, W.; Duchateau, G.S. Modeling of the relationship between dipeptide structure and dipeptide stability, permeability, and ACE inhibitory activity. J. Food Sci. 2009, 74, H243–H251. [Google Scholar] [CrossRef]
- Timofeeva, N.M.; Khavinson, V.K.; Malinin, V.V.; Nikitina, A.A.; Egorova, V.V. Effect of peptide Livagen on activity of digestive enzymes in gastrointestinal tract and non-digestive organs in rats of different ages. Adv. Gerontol. 2005, 16, 92–96. [Google Scholar]
- Shen, W.; Matsui, T. Intestinal Absorption of Small Peptides: A Review. Int. J. Food Sci. Technol. 2019, 54, 1942–1948. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Aliaga, I.; Daniel, H. Mammalian Peptide Transporters as Targets for Drug Delivery. Trends Pharmacol. Sci. 2002, 23, 434–440. [Google Scholar] [CrossRef]
- Hutterer, M.; Nowosielski, M.; Putzer, D.; Waitz, D.; Tinkhauser, G.; Kostron, H.; Muigg, A.; Virgolini, I.J.; Staffen, W.; Trinka, E.; et al. O-(2-18F-Fluoroethyl)-L-Tyrosine PET Predicts Failure of Antiangiogenic Treatment in Patients with Recurrent High-Grade Glioma. J. Nucl. Med. 2011, 52, 856–864. [Google Scholar] [CrossRef] [Green Version]
- Furuse, M. Molecular Basis of the Core Structure of Tight Junctions. Cold Spring Harb. Perspect. Biol. 2010, 2, a002907. [Google Scholar] [CrossRef] [PubMed]
- Namikawa, M.; Kakizaki, S.; Kaira, K.; Tojima, H.; Yamazaki, Y.; Horiguchi, N.; Sato, K.; Oriuchi, N.; Tominaga, H.; Sunose, Y.; et al. Expression of Amino Acid Transporters (LAT1, ASCT2 and XCT) as Clinical Significance in Hepatocellular Carcinoma. Hepatol. Res. 2015, 45, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Friesema, E.C.; Docter, R.; Moerings, E.P.; Verrey, F.; Krenning, E.P.; Hennemann, G.; Visser, T.J. Thyroid Hormone Transport by the Heterodimeric Human System L Amino Acid Transporter. Endocrinology 2001, 142, 4339–4348. [Google Scholar] [CrossRef] [PubMed]
- Fagerberg, L.; Hallström, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the Human Tissue-Specific Expression by Genome-Wide Integration of Transcriptomics and Antibody-Based Proteomics. Mol. Cell Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Puris, E.; Gynther, M.; Auriola, S.; Huttunen, K.M. L-Type Amino Acid Transporter 1 as a Target for Drug Delivery. Pharm. Res. 2020, 37, 88. [Google Scholar] [CrossRef] [PubMed]



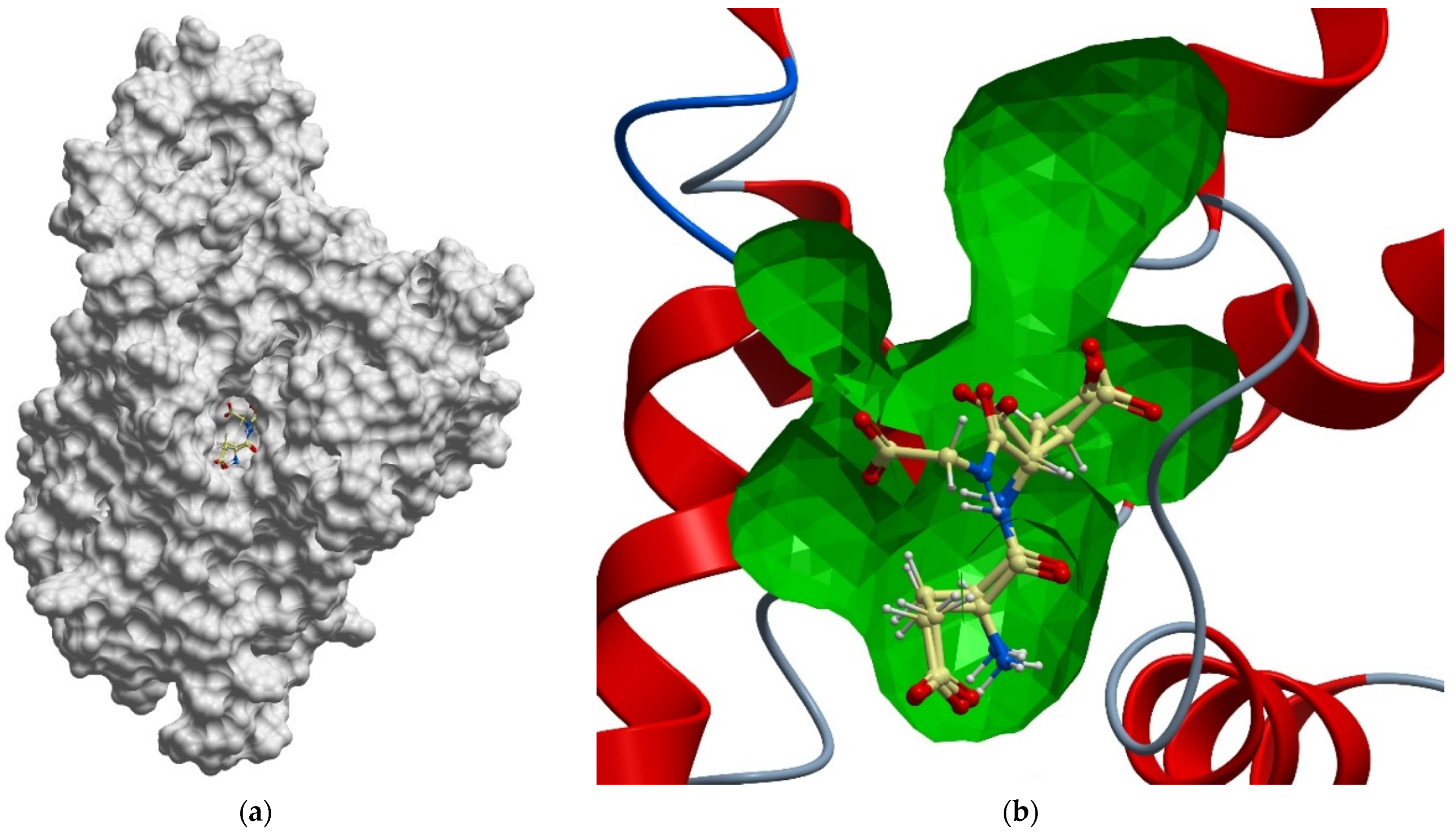{kind=link}
{kind=link}
| Ligand Structure | Mol. Name | ICM-Score | IC50, µM | Reference |
|---|---|---|---|---|
 | Tryptophan | −19.67 | 20.16 | [151] |
 | S-(3-bromo-4-methoxybenzyl)-l-cysteine | −19.31 | 33.20 | [152] |
 | l-2-amino-4-(3,5-dichlorophenyl)butanoic acid | −17.56 | 0.64 | [152] |
 | 5-(benzyloxy)-tryptophan | −17.59 | 1.48 | [152] |
 | 2-Amino-2-norbornanecarboxylic acid (BCH) | −15.00 | 155.10 | [152] |
| 2D Structure of Peptide | Name and Structure of Peptide | ICM-Score | Biological Activity |
|---|---|---|---|
 | Vesilut (ED) | −34.32 | Bladder regulator |
 | Chonluten (EDG) | −30.30 | Gastroprotector [161] Stress protector [162] |
 | Pinealon (EDR) | −30.29 | Neuroprotector [163] |
 | Cardiogen (AEDR) | −29.81 | Cardioprotector [164] |
 | Prostataget (DR) | −28.88 | Regulation of prostatic functions |
 | Ovagen (EDL) | −28.27 | Nephroprotector [165] Hepatoprotector [166] |
 | Cartalax (AED) | −26.65 | Chondroprotector [167] Regulation of skin fibroblast functions [168] |
 | Thymogen (EW) | −24.72 | Immunoprotector, medicinal product [14,169] |
 | Pancragen (KEDW-NH2) | −23.47 | Regulator of pancreatic functions [170] |
 | Livagen (KEDA) | −22.01 | Hepatoprotector [171] |
 | Testagen (KEDG) | −21.92 | Regulator of the male reproductive system |
 | Kistiget (DG) | −21.03 | Bladder regulator |
 | Epitalon (AEDG) | −20.38 | Geroprotector [12] Regulator of the epiphysis functions [172] Neuroprotector [173] |
 | Crystagen (EDP) | −20.22 | Immunoprotector [174] |
 | Vesselget (DS) | −19.82 | Vasoprotector [174] |
 | Nefroget (H-Asp(Pro)-OH) | −19.66 | Nephroprotector |
 | Vesugen (KED) | −18.91 | Vasoprotector [13] Neuroprotector [175] Geroprotector [176] |
 | Subcortiget (H-Asp(Ser)-OH) | −18.78 | Neuroprotector |
 | Bronchogen (AEDL) | −17.26 | Bronchoprotector [170] |
 | Fuaget (DL) | −16.01 | Hepatoprotector |
 | Prostomax (KEDP) | −13.58 | Regulation of prostatic functions [177] |
 | Orchiget (AE) | −13.03 | Regulation of prostatic functions |
 | Retiget (DA) | −12.82 | Retinaprotector |
 | Vilon (KE) | −12.31 | Immunoprotector [14,170] |
 | Thyroidget (DW) | −12.12 | Thyroid regulator |
 | Korthagen (AEDP) | −12.11 | Neuroprotector [13,170] |
 | Normoftal (H-Lys(H-Glu-OH)-OH) | −10.49 | Retinaprotector [178] Immunoprotector [170] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khavinson, V.; Linkova, N.; Kozhevnikova, E.; Dyatlova, A.; Petukhov, M. Transport of Biologically Active Ultrashort Peptides Using POT and LAT Carriers. Int. J. Mol. Sci. 2022, 23, 7733. https://doi.org/10.3390/ijms23147733
Khavinson V, Linkova N, Kozhevnikova E, Dyatlova A, Petukhov M. Transport of Biologically Active Ultrashort Peptides Using POT and LAT Carriers. International Journal of Molecular Sciences. 2022; 23(14):7733. https://doi.org/10.3390/ijms23147733
Chicago/Turabian StyleKhavinson, Vladimir, Natalia Linkova, Ekaterina Kozhevnikova, Anastasiia Dyatlova, and Mikhael Petukhov. 2022. "Transport of Biologically Active Ultrashort Peptides Using POT and LAT Carriers" International Journal of Molecular Sciences 23, no. 14: 7733. https://doi.org/10.3390/ijms23147733
APA StyleKhavinson, V., Linkova, N., Kozhevnikova, E., Dyatlova, A., & Petukhov, M. (2022). Transport of Biologically Active Ultrashort Peptides Using POT and LAT Carriers. International Journal of Molecular Sciences, 23(14), 7733. https://doi.org/10.3390/ijms23147733