
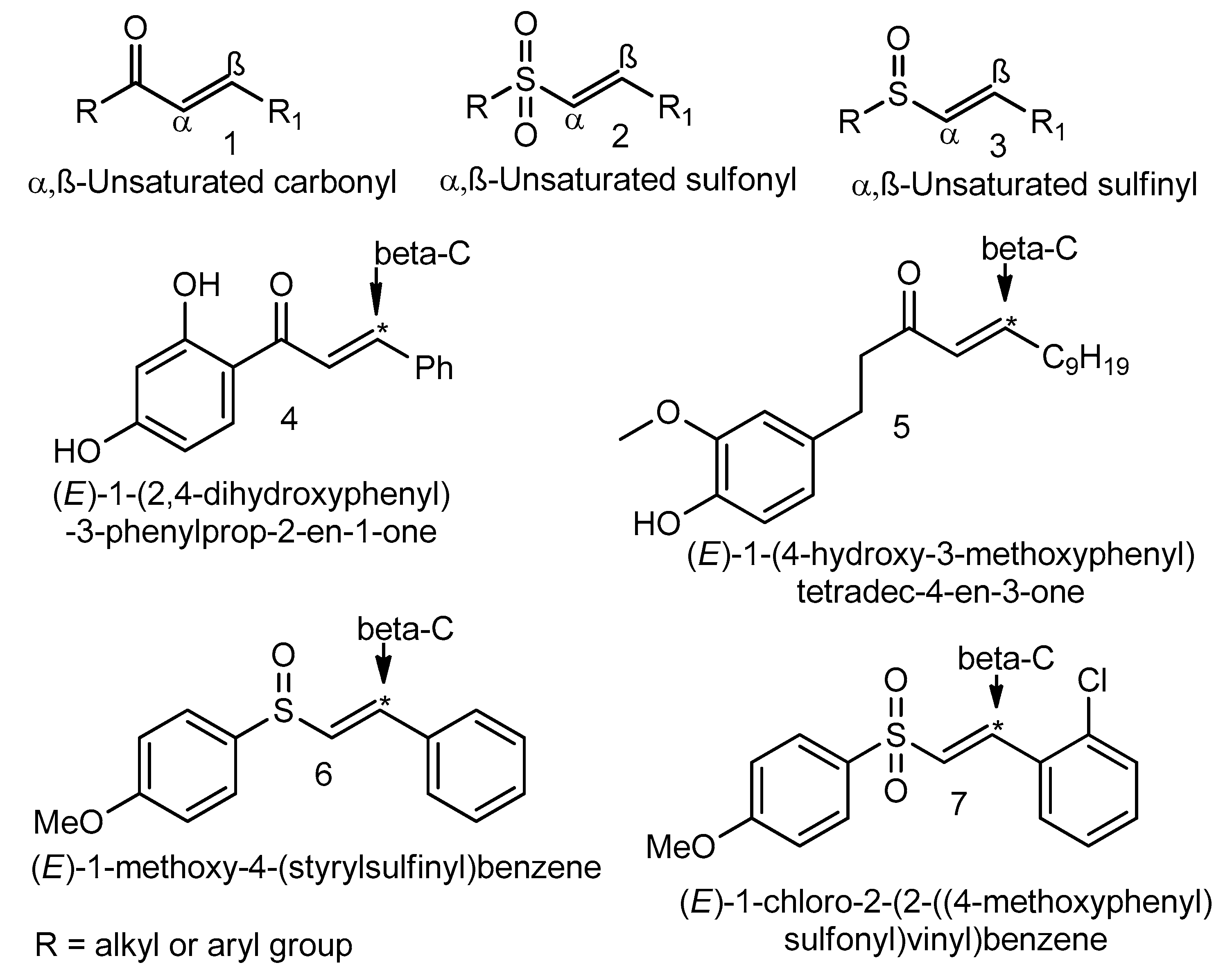
An Overview of NRF2-Activating Compounds Bearing α,β-Unsaturated Moiety and Their Antioxidant Effects

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Biologic Effects of NRF2/KEAP1 Signaling Pathway
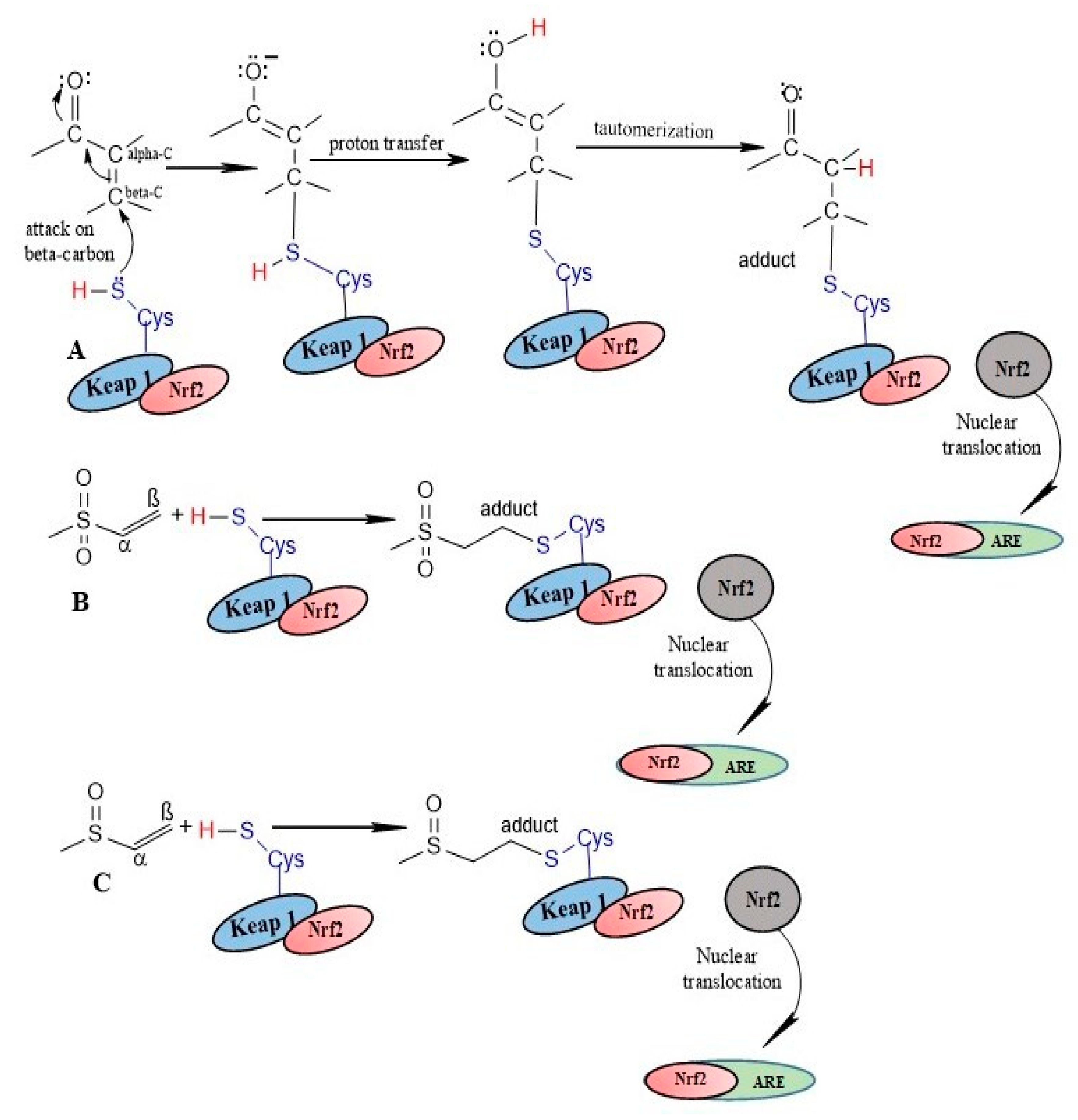
3. Modulation of NRF2/KEAP1 Signaling Pathway by α,β-Unsaturated Moiety-Bearing Compounds
4. α,β-Unsaturated Carbonyls
4.1. Sesquiterpene Lactones
4.1.1. Parthenolide
4.1.2. Helenalin
4.1.3. Alantolactone
4.1.4. Costunolide
4.2. Curcumin
4.3. J-Series Cyclopentenone Prostaglandin
4.4. Chalcone and Its Derivatives
4.5. Dimethyl Fumarate
5. α,β-Unsaturated Sulfonyls
5.1. Vinyl Sulfones
5.2. Vinyl Sulfonamides
5.3. Vinyl Sulfonates
6. α,β-Unsaturated Sulfinyls
Vinyl Sulfoxide

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S/N | Compound | Disease Studied | Model | NRF2 Activating Conc/Activity | Mechanism of Action | Biological Activity | Reference |
|---|---|---|---|---|---|---|---|
| 8 | Sesquiterpene lactones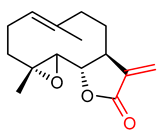 Parthenolide | Obesity | Mice, Adipocytes (3T3-L1), RAW264.7 | 1–8 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-adipogenesis | [124] |
| Obesity | 3T3-L1 Cell | 1–8 µM | NRF2 activation, Antioxidant, Anti-inflammatory | [125] | |||
| Osteoporosis | Human | 5–20 µM | NRF2 activation, Antioxidant, Anti-apoptosis | [126] | |||
| Breast cancer | Human breast cancer cell line MDA-MB 231 | 2.0 µM | NRF2 regulation, chemoresistance | [133] | |||
| Chronic lymphocytic leukemia | Human peripheral blood mononuclear cells (PBMCs) | 1.46 µM | NRF2 activation, Antioxidant, cytotoxicity | [111] | |||
| 9 | 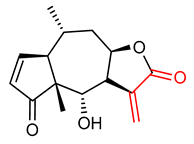 Helenalin | Acute hepatic injury | Male C57BL/6 Mice | 0.75–3.00 mg/kg | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [137] |
| 10 | 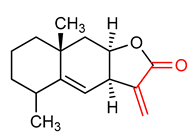 Alantolactone | Breast cancer | MCF-7 human breast cancer cells | 10–30 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, anticancer | [142] |
| Cancer | Heps1c1c7 cells | 1–10 µM | NRF2 activation, Antioxidant, anticancer | [143] | |||
| Chronic obstructive pulmonary disease (COPD) | Cigarette smoke-induced human bronchial epithelial cells | 1–10 µM | NRF2 activation, Antioxidant, Anti-inflammatory | [144] | |||
| 11 | 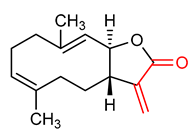 Costunolide | Acute liver injury | Mice | 20–40 mg/kg | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [146] |
| Oxidative damage | PC12 Cells | 5 µM | NRF2 activation, Antioxidant, neuroprotection | [150] | |||
| Tumor | RAW264.7 Macrophages | 0.1–1.0 µM | NRF2 activation, Anti-inflammatory | [147] | |||
| 12 | Curcumin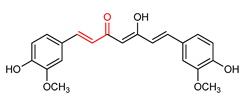 | Neurodegenerative diseases | Neuronal cells | 10 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant | [155] |
| Oxidative stress, inflammation | HepG2 Cells | 50 mg/kg | NRF2 activation, Antioxidant, Anti-inflammatory | [158] | |||
| Nasal diseases | Human nasal fibroblast | 0–5 µM | NRF2 activation, Antioxidant | [159] | |||
| Oxidative stress | Rats | 30 mg/kg | NRF2 activation, Antioxidant | [160] | |||
| 13 | Prostaglandin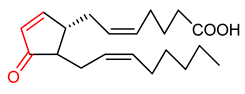 15-Deoxy-∆12,14-prostaglandin J2 | Breast cancer | Human breast cancer cells | 10 µmol/L | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant | [163] |
| Cancer | Mouse embryonic fibroblast (MEF) 293 cells | 0.5–10 µM | NRF2 activation, Antioxidant, Anticancer | [164] | |||
| 14 | Chalcones Trans-chalcone | Leishmannia amazonensis | L. amazonensis-infected macrophages | 2–12 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, | [169] |
| 15 | 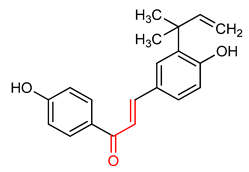 Licochalcone A | Cutaneous oxidative stress | UVA-irradiated human dermal fibroblast | 9 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [170] |
| 16 | 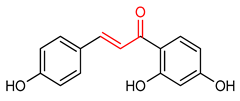 Isoliquiritigenin | Pancreatic injury | Mice | >3% | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [171] |
| 17 | 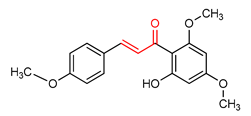 Chalcone flavokawain A | inflammation | Primary splenocytes | 2–30 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [172] |
| 18 | DMF Dimethyl fumarate | Oxidative stress | Mouse ovary | 20 mg/kg | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, | [175] |
| Multiple sclerosis | Multiple sclerosis patient | 0–400 | NRF2 activation, Antioxidant, | [173] | |||
| Parkinson’s disease | Mice | 0.05–80 µM | NRF2 activation, Antioxidant, Anti-inflammatory | [177] | |||
| 19 | Vinyl Sulfones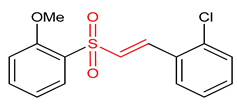 (E)-1-chloro-2-(2-((2-methoxyphenyl)sulfonyl)vinyl)benzene | Multiple sclerosis | HEK293 | 10 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, | [2] |
| Parkinson’s disease | PD animal model | 1–20 µM | NRF2 activation, Antioxidant, Anti-inflammatory | [23] | |||
| Parkinson’s disease | PD animal model | 1–10 µM | NRF2 activation, Antioxidant, Neuroprotection | [32] | |||
| 20 |  (E)-4-(3-(4-((2-(3-fluoropyridin-2-yl)vinyl)sulfonyl)phenoxy)propyl)morpholine hydrochloride | Parkinson’s disease | PD mice | 0.3–10 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Neuroprotection | [9] |
| 21 |  (E)-1-(2-((4-methoxyphenyl)sulfonyl)vinyl)2-(trifluoromethyl)benzene | Parkinson’s disease | PD mice | 20 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Neuroprotection | [32] |
| 22 | 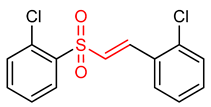 (E)-1-chloro-2-(2-((2-chlorophenyl)sulfonyl)vinyl)benzene | Oxidative stress | PC12 Cells | 2.5–1.0 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Neuroprotection | [194] |
| 23 | 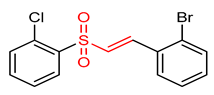 (E)-1-bromo-2-(2-((2-chlorophenyl)sulfonyl)vinyl)benzene | Oxidative stress | PC12 Cells | 0.5–1.0 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Neuroprotection | [194] |
| 24 | Vinyl Sulfonamides (E)-N,2-diphenylethenesulfonamide | Parkinson’s disease | PD mouse | >10 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [17] |
| 25 | 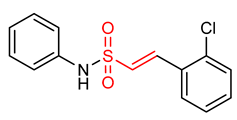 (E)-2-(2-chlorophenyl)-N-phenylethesulfonamide | Parkinson’s disease | PD mouse | >10 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [17] |
| 26 | 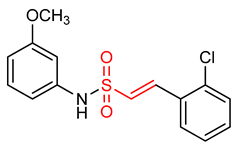 (E)-2-(2-chlorophenyl)-N-(2-methoxyphenyl)ethenesulfonamide | Parkinson’s disease | PD mouse | 6.35 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [17] |
| 27 | 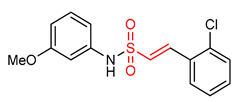 (E)-2-(2-chlorophenyl)-N-(3-methoxyphenyl)ethane sulfonamide | Parkinson’s disease | PD mouse | >10 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [17] |
| 28 | 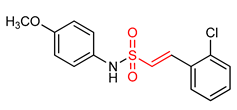 (E)-2-(2-chlorophenyl)-N-(4-methoxyphenyl)ethane sulfonamide | Parkinson’s disease | PD mouse | >10 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [17] |
| 29 | Vinyl Sulfonates (E)-2-methoxyphenyl 2-(2-chlorophenyl)ethenesulfonate | Parkinson’s disease | PD mouse | 0.076 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [17] |
| 30 | 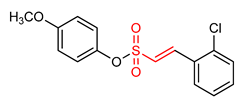 (E)-4-methoxyphenyl 2-(2-chlorophenyl)ethenesulfonate | Parkinson’s disease | PD animal model | 0.237 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [17] |
| 31 |  (E)-3-methoxyphenyl 2-(2-chlorophenyl)ethenesulfonate | Parkinson’s disease | PD mouse | 0.165 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, Anti-inflammatory | [17] |
| 32 | Vinyl Sulfoxides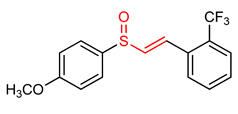 (E)-1-(2-((4-methoxyphenyl)sulfinyl)vinyl)-2-(trifluoromethyl)benzene | Parkinson’s disease | BV-2 Cells | 20 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Neuroprotection | [32] |
| 33 | 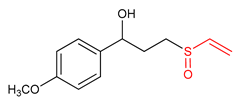 1-(4-methoxyphenyl)-3-(vinylsulfinyl)propan-1-ol | Parkinson’s disease | BV-2 Cells | 20 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, | [3] |
| 34 | 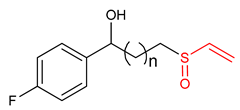 1-(4-fluorophenyl)-3-(vinylsulfinyl)propan-1-ol | Parkinson’s disease | BV-2 Cells | 20 µM | electrophilic modification of KEAP1 cysteine residues | NRF2 activation, Antioxidant, | [3] |
| Entry | Compound | Clinical Trial/FDA Approval | Targeted Disease | Reference |
|---|---|---|---|---|
| 8 | Parthenolide | Clinical trial | Cancer | NCT00133341 |
| 12 | Curcumin | Clinical trial | Impaired glucose tolerance and insulin resistance/ type 2 diabetes | NCT01052025 |
| 15 | Licochalcone A | Clinical trial | Human oral squamous cell carcinoma | NCT03292822 |
| 18 | Dimethyl fumarate | FDA approved | Multiple sclerosis | NCT00810836 |
7. Pharmacological Profile of α,β-Unsaturated Structure-Bearing NRF2 Activators
8. Structure–Activity Relationship of α,β-Unsaturated Structure-Bearing NRF2 Activators
9. α,β-Unsaturated-Based NRF2 Activator in Parkinson’s Disease
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sulsen, V.P.; Martino, V.S. Sesquiterpene Lactones: Advances in Their Chemistry and Biological Aspects, 1st ed.; Springer Nature: Cham, Switzerland, 2018. [Google Scholar]
- Carlstrom, K.E.; Chinthkindi, P.K.; Espinosa, B.; Nimer, F.A.; Arner, E.S.J.; Arvidsson, P.I.; Piehl, F.; Johnasson, K. Characterization of more selective central nervous system NRF2-activating novel vinyl sulfoxime compounds compared to dimethyl fumarate. Neurotherapeutics 2020, 17, 114–1152. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.Y.; Hwang, H.S.; Nam, G.; Choi, K. Synthesis and NRF2 activating ability of thiourea and vinyl sulfoxide derivatives. Bull. Korean Chem. Soc. 2013, 34, 2317–2320. [Google Scholar] [CrossRef] [Green Version]
- Mayer, R.J.; Allihn, P.W.A.; Hampel, N.; Mayer, P.; Sieber, S.A.; Ofial, A.R. Electrophilic reactivities of cyclic enones and α,β-unsaturated lactones. Chem. Sci. 2021, 12, 4850–4865. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yazaki, R.; Ohsima, T. Chemo-and region selective direct functional group installation through catalytic hydroxyl group selective conjugation addition of amino alchohols to α,β-unsaturated sulfonyl compounds. Org. Lett. 2016, 18, 3350–3353. [Google Scholar] [CrossRef] [PubMed]
- Lipton, S.; Rezaie, T.; Nutter, A.; Lopez, K.M.; Parker, J.; Kosaka, K.; Satoh, T.; McKercher, S.R.; Masiliah, E.; Nakanishi, N. Therapeutic advantage of pro-electrophoilic drugs to activate the Nrf2/ARE pathway in Alzheimer’s disease models. Cell Death Dis. 2016, 7, e2499. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Saha, S.; Buttari, B.; Profumo, E.; Saso, L. Activation of NRF2 signaling pathway by natural and synthetic chalcones: A therapeutic road map for oxidative stress. Expert Rev. Clin. Pharmacol. 2021, 14, 465–480. [Google Scholar] [CrossRef]
- Robledinos-Anton, N.; Fernandez-Gines, R.; Manda, G.; Cuadrado, A. Activators and inhibitors of NRF2: A review of their potential for clinical development. Oxid. Med. Cell. Longev. 2019, 2019, 9372182. [Google Scholar] [CrossRef] [PubMed]
- Chio, J.W.; Kim, S.; Park, J.; Kim, H.J.; Shin, S.J.; Kim, J.W.; Woo, S.Y.; Lee, C.; Han, S.M.; Lee, J.; et al. Optimization of vinyl sulfone derivatives as potent nuclear factor erythroid 2-related factor 2 [NRF2] activities for disease therapy. J. Med. Chem. 2019, 62, 811–830. [Google Scholar] [CrossRef] [PubMed]
- Yagishita, Y.; Gatbonton-Schwager, T.N.; McCallum, M.L.; Kensler, T.W. Current landscape of NRF2 biomarkers in clinical trials. Antioxidants 2020, 9, 716. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, F.; Sun, F.; Gu, K.F.; Dong, S.; He, D. Dimethyl fumarate for multiple sclerosis Cochrane Database Syst. Rev. 2015, 4, CD011076. [Google Scholar]
- Fox, R.J.; Miller, D.H.; Phillips, J.T.; Hutchinson, M.; Havrdova, E.; Kita, M.; Yang, M.; Raghupathi, K.; Novas, M.; Sweetser, M.T.; et al. Placebo-controlled phase 3 study of oral BG-12 or glatiramer in multiple sclerosis. N. Engl.J. Med. 2012, 367, 1087–1097. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. therapeutic roles of curcumin: Lessons learned from clinical trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Rakesh, K.P.; Ravidar, L.; Fang, W.Y.; Qin, H.L. Pharmaceutical and medicinal significance of sulfur (SVI)-containing motifs for drug discovery: A critical review. Eur. J. Med. Chem. 2019, 162, 679–734. [Google Scholar] [CrossRef]
- Lu, L.; Feng, Q.; Su, T.; Cheng, Y.; Huang, Z.; Huang, Q.; Liu, Z. Chapter 35, Pharmacoepigenetics of Chinese herbal components in cancer. In Translational Epigenetics, Pharmacoepigenetics; Cacabelos, R., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 10, pp. 859–869. [Google Scholar]
- Rasul, A.; Parveen, S.; Ma, T. Costunolide: A novel anticancer sesquiterpene lactone. Bangladish J. Pharmacol. 2012, 7, 6–13. [Google Scholar]
- Choi, J.W.; Shin, S.J.; Kim, H.J.; Park, J.-H.; Kim, J.K.; Lee, E.H.; Pae, A.N.; Bahn, Y.S.; Park, K.D. Antioxidant anti-inflammatory, and neuroprotective effects of novel vinyl sulfonate compounds as NRF2 activator. ACS Med. Chem. Lett. 2019, 10, 1061–1067. [Google Scholar] [CrossRef]
- Silva, M.; Pruccoli, L.; Morronoi, F.; Sita, G.; Seghetti, f.; Viegas, C.; Tarozzi, A. The keap 1/NRF2-ARE pathway as a pharmacological target for chalcones. Molecules 2018, 23, 1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Testa, B.; Fahr, A. Lipophilicity and its relationship with passive drug permeation. Pharm. Res. 2011, 28, 962–977. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.A.; Barry, C.J.; Walker, D.K. Design of drugs involving the concepts and theories of drug metabolism and pharmacokinetics. Med. Res. Rev. 1996, 16, 243–266. [Google Scholar] [CrossRef]
- Remko, M. Molecular structure, Pka, lipophilicity, solubility and absorption of biologically active aromatic and heterocyclic sulphonamides. J. Mol. Struct. Theochem. 2010, 944, 34–42. [Google Scholar] [CrossRef]
- Bukhari, S.N.; Jantan, I.; Jasamai, M. Anti-inflammatory trends of 1,3-diphemyl-2- propene-1- one derivatives. Mini Rev. Med. Chem. 2013, 13, 87–94. [Google Scholar] [CrossRef]
- Lee, J.A.; Kim, J.H.; Woo, S.Y.; Son, H.Y.; Han, S.H.; Jang, B.K.; Choi, J.W.; Kim, D.J.; Park, K.D.; Hwang, O. A novel compound VSC2 has anti-inflammatory and antioxidant properties in microglia and in Parkinson’s disease model. Br. J. Pharmacol. 2015, 172, 1087–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankaran, S.; Balasubramanian, R. Insight into the lipophilicity of selected monosubstituted chalcones. Pak. J. Pharm. Sci. 2018, 31, 941–946. [Google Scholar]
- Smith, D.A.; Cucurull-Sanchez, L. The adaptive in combo strategy. In Comprehensive Medicinal Chemistry II; Pfizer Global Research and Developmental: Sandwich, UK, 2007; Volume 5, pp. 957–969. [Google Scholar]
- Di, L.; Kerns, E.H. Drug-Like Properties, Concepts, Structure Design and Methods from ADME to Toxicity Optimization, 2nd ed.; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Karlgen, M.; Bergstrom, C.A.S. How physicochemical properties of drugs affect their metabolism and clearance. In New Horizons in Predictive Drug Metabolism and Pharmacokinetics; Royal Society of Chemistry: Cambridge, UK, 2015; pp. 1–26. [Google Scholar]
- Randy, R.M.; Madeira, M.; Wood, H.B.; Geissler, W.M.; Raab, C.E.; Martin, I.J. Integrating the impact of lipophilicity on potency and pharmacokinetic parameters enables the use of diverse chemical space during small molecule drug optimization. J. Med. Chem. 2020, 63, 12156–12170. [Google Scholar]
- Alrubaie, L.A.; Muhasin, R.J.; Mousa, M.N. Synthesis, characterization and evaluation of anti-inflammatory properties of novel α,β-unsaturated ketones. Tropi. J. Pharm. Res. 2020, 19, 147–154. [Google Scholar] [CrossRef]
- Kim, S.S.; Gallaher, D.D.; Csallany, A.S. Aldehydes and related carbonyl compounds in rat and human urine. Lipids 1999, 34, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Sappanen, C.M.; Csallany, A.S. Simultaneous determination of lipophilic aldehydes by high-performance liquid chromatography in vegetable oil. J. Am. Chem. Soc. 2001, 78, 1253. [Google Scholar] [CrossRef]
- Woo, S.Y.; Kim, J.H.; Moon, M.K.; Han, S.-H.; Yeon, S.K.; Choi, J.W.; Jang, B.K.; Song, H.J.; Kang, Y.G.; Kim, J.W.; et al. Discovery of vinyl sulfones as novel class of neuroprotective agents towards Parkinson’s disease therapy. J. Med. Chem. 2014, 57, 1473–1487. [Google Scholar] [CrossRef]
- Sun, Z.; Chin, Y.E.; Zhang, D.D. Acetylation of Nrf2 by p300/CBP augments promoter-specific DNA binding of Nrf2 during the antioxidant response. Mol. Cell. Biol. 2009, 29, 2658–2672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodore, M.; Kawai, Y.; Yang, J.; Kleshchenko, Y.; Reddy, S.P.; Villalta, F.; Arinze, I.J. Multiple nuclear localization signals function in the nuclear import of the transcription factor Nrf2. J. Biol. Chem. 2008, 283, 8984–8988. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo, M.C.; Zhang, D.D. The emerging role of the Nrf2-Keap1 signaling pathway in cancer. Genes Dev. 2013, 27, 2179–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.-L. The KEAP1-NRF2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nioi, P.; Nguyen, T.; Sherratt, P.J.; Pickett, C.B. The carboxy-terminal Neh3 domain of Nrf2 is required for transcriptional activation. Mol. Cell. Biol. 2005, 25, 10895–10906. [Google Scholar] [CrossRef] [Green Version]
- Katoh, Y.; Itoh, K.; Yoshida, E.; Miyagishi, M.; Fukamizu, A.; Yamamoto, M. Two domains of Nrf2 cooperatively bind CBP, a CREB binding protein, and synergistically activate transcription. Genes Cells 2001, 6, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Rada, P.; Rojo, A.I.; Evrard-Todeschi, N.; Innamorato, N.G.; Cotte, A.; Jaworski, T.; Tobon-Velasco, J.C.; Devijver, H.; Garcia-Mayoral, M.F.; Van Leuven, F.; et al. Structural and functional characterization of Nrf2 degradation by the glycogen synthase kinase 3/beta-TrCP axis. Mol. Cell. Biol. 2012, 32, 3486–3499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Liu, K.; Geng, M.; Gao, P.; Wu, X.; Hai, Y.; Li, Y.; Li, Y.; Luo, L.; Hayes, J.D.; et al. RXRalpha inhibits the NRF2-ARE signaling pathway through a direct interaction with the Neh7 domain of NRF2. Cancer Res. 2013, 73, 3097–3108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. KEAP1 represses nuclear activation of antioxidant responsive elements by NRF2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, A.; Kang, M.I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative stress sensor KEAP1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of NRF2. Mol. Cell. Biol. 2004, 24, 7130–7139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, Y.; Suzuki, T.; Inoue, J.; Iso, T.; Wells, G.; Moore, T.W.; Mizushima, T.; Dinkova-Kostova, A.T.; Kasai, T.; Kamei, T.; et al. Molecular basis for the disruption of KEAP1–NRF2 interaction via Hinge & Latch mechanism. Commun. Biol. 2021, 4, 576. [Google Scholar] [PubMed]
- Ma, Q. Role of NRF2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Baird, L.; Llères, D.; Swift, S.; Dinkova-Kostova, A.T. Regulatory flexibility in the NRF2-mediated stress response is conferred by conformational cycling of the KEAP1-NRF2 protein complex. Proc. Natl. Acad. Sci. USA 2013, 110, 15259–15264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertel, W.; Kremer, J.P.; Kenney, J.; Steckholzer, U.; Jarrar, D.; Trentz, O.; Schildberg, F.W. Downregulation of proinflammatorry cytokine release in whole blood from septic patients. Blood 1995, 85, 1341–1347. [Google Scholar] [CrossRef] [Green Version]
- Pietsch, E.C.; Chan, J.Y.; Torti, F.M.; Torti, S.V. NRF2 mediates the induction of ferritin H in response to xenobiotics and cancer chemopreventive dithiolethiones. J. Biol. Chem. 2003, 278, 2361–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An overview of NRF2 signaling pathway and its role inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Yamamoto, Y.; Wang, Q.M. The IKK NF-kappa B system: A treasure trove for drug development. Nat. Rev. Drug Discov. 2004, 3, 17–26. [Google Scholar] [CrossRef]
- Soares, M.P.; Seldon, M.P.; Gregoire, I.P.; Vassilevskaia, T.; Berberat, P.O.; Yu, J.; Tsui, T.Y.; Bach, F.H. Heme oxygenase-1 modulates the expression of adhesion molecules associated with endothelial cell activation. J. Immunol. 2004, 172, 3553–3563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerra, V.G.; Negi, G.; Sharma, S.S.; Kumar, A. Potential therapeutic effects of the simultaneous targeting of the Nrf2 and NF-B pathways in diabetic neuropathy. Redox Biol. 2013, 1, 394–397. [Google Scholar] [CrossRef] [Green Version]
- Kovac, S.; Angelova, P.R.; Holmström, K.M.; Zhang, Y.; Dinkova-Kostova, A.T.; Abramov, A.Y. Nrf2 regulates ROS production by mitochondria and NADPH oxidase. Biochim. Biophys. Acta 2015, 1850, 794–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strom, J.; Xu, B.; Tian, X.; Chen, Q.M. Nrf2 protects mitochondrial decay by oxidative stress. FASEB J. 2016, 30, 66–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.G.; Zhang, Y.Q.; Wu, Z.Z.; Hsieh, C.W.; Chu, C.S.; Wung, B.S. Peanut arachidin-1 enhances Nrf2-mediated protective mechanisms against TNF-α-induced ICAM-1 expression and NF-κB activation in endothelial cells. Int. J. Mol. Med. 2018, 41, 541–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filardi, E.S.; Vega, M.A.; Mateos, P.S.; Corbi, A.L.; Kroger, A.P. Heme Oxygenase-1 expression in M-CSF-polarized M2 macrophages contributes to LPS-induced IL-10 release. Immunobiology 2010, 215, 788–795. [Google Scholar] [CrossRef]
- Choi, K.M.; Kashyap, P.C.; Dutta, N.; Stoltz, G.J.; Ordog, T.; Donohue, T.S.; Bauer, A.J.; Linden, D.R.; Szurszewski, J.H.; Gibbons, S.J.; et al. CD206-positive M2 macrophages that express heme oxygenase-1 protect against diabetic gastroparesis in mice. Gastroenterology 2010, 138, 2399–2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.; Zhai, X.; Wang, G.; Tian, X.; Gao, D.; Shi, L.; Wu, H.; Fan, Q.; Peng, J.; Liu, K.; et al. Salvianolic acid B protects against acetaminophen hepatotoxicity by inducing NRF2 and phase II detoxification gene expression via activation of the P13K and PKC signaling pathway. J. Pharmacol. Sci. 2015, 127, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiss, E.H.; Schachner, D.; Zimmermann, K.; Dirsch, V.M. Glucose availability is a decisive factor for NRF2-mediated gene expression. Redox Biol. 2013, 1, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambel, S.S.; Santos-Goncalves, A.; Duarte, T.L. The dual of NRF2 in nonalcoholic fatty liver disease: Regulation of antioxidant defences and hepatic lipid metabolism. Biomed. Res. Int. 2015, 2015, 597134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Zaaba, N.E.; Yasin, J.; Hamadi, N.; Ali, B.H. Cardiac inflammation, oxidative stress, NRF2 expression, and coagulation events in mice with experimental chronic kidney disease. Oxid. Med. Cell. Longev. 2021, 2021, 8845607. [Google Scholar] [CrossRef] [PubMed]
- Egbujor, M.C.; Garrido, J.; Borges, F.; Saso, L. Sulfonamide a valid scaffold for antioxidant drug development. Mini-Rev. Org. Chem. 2022, 19. [Google Scholar] [CrossRef]
- Satoh, H.; Moriguchi, T.; Taguchi, K.; Takai, J.; Maher, J.M.; Suzuki, T.; Winnard, P.T., Jr.; Raman, V.; Ebina, M.; Nukiwa, T.; et al. Nrf2-deficiency creates a responsive microenvironment for metastasis to the lung. Carcinogenesis 2010, 31, 1833–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiramoto, K.; Satoh, H.; Suzuki, T.; Moriguchi, T.; Pi, J.; Shimosegawa, T.; Yamamoto, M. Myeloid lineage-specific deletion of antioxidant system enhances tumor metastasis. Cancer Prev. Res. 2014, 7, 835–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagishita, Y.; Fukutomi, T.; Sugawara, A.; Kawamura, H.; Takahashi, T.; Pi, J.; Uruno, A.; Yamamoto, M. Nrf2 protects pancreatic β-cells from oxidative and nitrosative stress in diabetic model mice. Diabetes 2014, 63, 605–618. [Google Scholar] [CrossRef] [Green Version]
- Uruno, A.; Farusawa, Y.; Yagishita, T.; Fukutomi, T.; Maramatsu, H.; Negishi, T.; Sugawara, A.; Kensler, T.W.; Yamamoto, M. The Keap1-Nrf2 system prevents onset of diabeties mellitus. Am. Soc. Microbiol. Mol. Cell. Biol. 2013, 33, 2996–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, A.J.; Lau, H.; Li, S.; Ichii, H. Potential benefits of Nrf2/Keap1 targeting in pancreatic islet cell transplantation. Antioxidants 2020, 9, 321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, C.M.; Faulenbach, M.; Vaag, A.; Vølund, A.; Ehses, J.A.; Seifert, B.; Mandrup-Poulsen, T.; Donath, M.Y. Interleukin-1-receptor antagonist in type 2 diabetes mellitus. N. Engl. J. Med. 2007, 356, 1517–1526. [Google Scholar] [CrossRef] [Green Version]
- Priddy, C.; Li, J. The role of the Nrf2/Keap1 signaling cascade in mechanobiology and bone health. Bone Rep. 2021, 15, 101149. [Google Scholar] [CrossRef] [PubMed]
- David, J.A.; Rifkin, W.J.; Rabbani, P.S.; Ceradini, D.J. The Nrf2/Keap1/ARE pathway and oxidative stress as a therapeutic target in type II diabetes mellitus. J. Diabetes Res. 2017, 2017, 4826724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chataigner, I.; Panel, C.; Gerard, H.; Piettre, S.R. Sulfonyl Vs. Carbonyl group: Which is the more electron–withdrawing? Chem. Commun. 2007, 31, 3288–3290. [Google Scholar] [CrossRef] [PubMed]
- Holland, R.; Fishbein, J.C. Chemistry of the cysteine sensors in Kelch-like ECH- associated protein 1. Antioxid. Redox Signal. 2010, 13, 1749–1761. [Google Scholar] [CrossRef]
- Pearson, R.G. Hard and soft acids and bases the evolution of a chemical concept. Coord. Chem. Rev. 1990, 100, 403–425. [Google Scholar] [CrossRef]
- Saverland, M.; Mertes, R.; Morozzi, C.; Eggler, A.L.; Gamon, L.F. Kinetic assessment of Michael addition reaction of alpha, beta-unsaturated carbonyl compounds to amino acid and protein thiols. Free Radic. Biol. Med. 2021, 169, 1–11. [Google Scholar] [CrossRef]
- Reusch, W. Nucleophilic addition to α,β-unsaturated carbonyl compounds. In Virtual Textbook of Organic Chemistry, 5th ed.; Michigan State University Press: East Lansing, MI, USA, 1999; p. 1162. [Google Scholar]
- Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Cole, R.N.; Itoh, K.; Wakabayashi, N.; Katoh, Y.; Yamamoto, M.; Talalay, P. Direct evidence that sulfhydryl groups of KEAP1 are the sensors regulating induction of pahse 2 enzymes that protect against carcinogens and oxidants. Proc. Natl. Acad. Sci. USA 2002, 99, 11908–11913. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, A.; Kang, M.I.; Watai, Y.; Tong, K.I.; Shibata, T.; Uchida, K.; Yamamoto, M. Oxidative and electrophilic stresses activate NRF2 through inhibition of ubiquitination activity of KEAP1. Mol. Cell. Biol. 2006, 26, 221–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggler, A.L.; Liu, G.; Pezzuto, J.M.; Van Breemen, R.B.; Mesecar, A.D. Modifying specific cysteines of the electrophile-sensing human Keap1 proteinis insufficient to disrupt binding to the Nrf2 domain Neh2. Proc. Natl. Acad. Sci. USA 2005, 102, 10070–10075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuadrado, A.; Manda, G.; Hassan, A.; Alcaraz, M.J.; Barbas, C.; Daiber, A.; Ghezzi, P.; Leon, R.; Lopez, M.G.; Oliva, B.; et al. Transcription factor NRF2 as a therapeutic target for chronic diseases: A system medicine approach. Pharmacol. Rev. 2018, 70, 348–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuadrado, A.; Rojo, A.I.; Wells, G.; Hayes, J.D.; Cousin, S.P.; Rumsey, W.L.; Attucks, O.C.; Franklin, S.; Levonen, A.L.; Kensler, T.W.; et al. Therapeutic targeting of the NRF2 and KEAP1 partnership in chronic diseases. Nat. Rev. Drug Discov. 2019, 18, 295–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the KEAP1–NRF2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Egbujor, M.C.; Petrosino, M.; Zuhra, K.; Saso, L. The Role Of Organosulfur Compounds As Nrf2 Activators And Their Antioxidant Effects. Antioxidants 2022, 11, 1255. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional regulation by NRF2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. NRF2 signaling pathaway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Cores, A.; Piquero, M.; Villacapa, M.; Leon, R.; Menendez, J.C. NRF2 regulation processes as a source of potential drug targets against neurodegenerative diseases. Biomolecules 2020, 10, 904. [Google Scholar] [CrossRef]
- Saito, R.; Suzuki, T.; Hiramoto, K.; Asami, S.; Naganuma, E.; Suda, H.; Iso, T.; Yamamoto, H.; Morita, M.; Baird, L.; et al. Characterizations of Three Major Cysteine Sensors of Keap1 in Stress Response. Mol. Cell. Biol. 2015, 36, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Levonen, A.L.; Landar, A.; Ramachandran, A.; Ceaser, E.K.; Dickinson, D.A.; Zanoni, G.; Morrow, J.D.; Darley-Usmar, V.M. Cellular mechanisms of redox cell signaling: Role of cysteine modification in controlling antioxidant defences in response to electrophilic lipid oxidation products. Biochem. J. 2004, 378, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Wakabayashi, N.; Dinkova-Kostova, A.T.; Holtzclaw, W.D.; Kang, M.; Kobayashi, A.; Yamamoto, M.; Kensler, T.W.; Talalay, P. Protection against electrophile and oxidant stress by induction of the phase 2 response: Fate of cysteines of the KEAP1 sensor modified by inducers. Proc. Natl. Acad. Sci. USA 2004, 101, 2040–2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Suzuki, T.; Kobayashi, A.; Wakabayashi, J.; Maher, J.; Motohashi, H.; Yamamoto, M. Physiological significance of reactive cysteine residues of KEAP1 in determining NRF2 activity. Mol. Cell. Biol. 2008, 28, 2758–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meadows, D.C.; Gervay-Hague, J. Vinyl sulfones: Synthetic preparations and medicinal chemistry applications. Med. Res. Rev. 2006, 26, 793–814. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.E.; Lyness, W.I. The effect of methyl-mercapto, methylsulfinyl, and methylsulfonyl groups on the equilibrium in three-carbon prototropic systems. J. Am. Chem. Soc. 1964, 86, 3840–3846. [Google Scholar] [CrossRef]
- Cilento, G. The expansion of the sulfur outer shell. Chem. Rev. 1960, 60, 147–167. [Google Scholar] [CrossRef]
- Inomata, K.; Hirata, T.; Suhara, H.; Kinoshita, H.; Kotake, H.; Senda, H. Stereochemistry of the conversion of γ-substituted (E)-vinyl sulfones to the corresponding allyl sulfones. Determination of the relative drgree of syn-effect. Chem. Lett. 1988, 17, 2009–2012. [Google Scholar] [CrossRef] [Green Version]
- Snyder, H.R.; Eliel, E.L.; Charnahan, R.E. Studies in the sulfone series. J. Am. Chem. Soc. 1951, 73, 3258. [Google Scholar] [CrossRef]
- Snyder, H.R.; Hallada, D.P. α,β-Unsaturated sulfonyl compounds in Diels-Alder reaction II. J. Am. Chem. Soc. 1952, 74, 5595–5597. [Google Scholar] [CrossRef]
- Saddler, J.C.; Conrad, P.C.; Fuchs, P.L. A new (3+3) annulations route to isoquinoline-3-carboxylates. Tetrahedron Lett. 1978, 19, 5079–5081. [Google Scholar] [CrossRef]
- Liu, L.K.; Chi, Y.; Jen, K.Y. Copper-catalyzed additions of sulfonyl iodides to simple and cyclic alkenes. J. Org. Chem. 1980, 45, 406–410. [Google Scholar] [CrossRef]
- Taber, D.F.; Saleh, S.A. Branching strategy in organic synthesis 2-Reveral of olefin polarization with concomitant carbon-carbon bond formation. J. Org. Chem. 1981, 46, 4817–4819. [Google Scholar] [CrossRef]
- Smith, M.B.; March, J. Advanced Organic Chemistry: Ractions, Mechanism and Structure, 6th ed.; Wiley-Interscience: New York, NY, USA, 2007. [Google Scholar]
- Rodrigues, T.; Recker, D.; Schneider, P.; Schneider, G. Counting on natural products for drug design. Nat. Chem. 2016, 8, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Arshad, L.; Jantan, T.; Bukhari, S.N.A.; Haque, A. Immunosuppressive effects of natural α,β-unsaturated carbonyl-based compounds, and their avalogs and derivatives, on immune cells: A review. Front. Pharmacol. 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, N.W.; Singh, B.; Buchan, G.; O’Brien, J.; Arroyo, A.D.; Liu, X.; Sobol, R.W.; Blair, I.A.; Mesaros, C.A.; Wendell, S.G. Primary saturation of α,β-unsaturated carbonyl containing fatty acid does not abolish electrophilicity. Chem.-Biol. Interact. 2021, 350, 109689. [Google Scholar] [CrossRef] [PubMed]
- Thapa, P.; Upadhyay, S.P.; Suo, W.Z.; Singh, V.; Gurung, P.; Lee, E.S.; Sharma, R.; Sharm, M. Chalcone and its analogs:therapeutic and diagnostic applications in Alzheimer’s disease. Bioorganic Chem. 2021, 108, 104681. [Google Scholar] [CrossRef] [PubMed]
- Naidu, S.D.; Dinkova-Kostova, A.T. KEAP1, a cysteine-based sensor and a drug target for the prevention and treatment of chronic disease. Open Biol. 2020, 10, 200105. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.W.; Chun, K.-S.; Kim, D.-H.; Kim, S.-J.; Kim, S.H.; Cho, N.-C.; Na, H.-K.; Surh, Y.-J. Curcumin induces stabilization of Nrf2 protein through Keap1 cysteine modification. Biochem. Pharmacol. 2020, 173, 113820. [Google Scholar] [CrossRef] [PubMed]
- Grimsrud, P.A.; Xie, H.; Griffin, T.J.; Bernlohr, D.A. Oxidative stress and covalent modification of protein withbioactive aldehydes. J. Biol. Chem. 2008, 283, 21837–21841. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.-H.; Hwang, Y.; Liu, H.; Wang, X.J.; Zhang, Y.; Stephenson, K.K.; Boronina, T.N.; Cole, R.N.; Dinkova-Kostova, A.T.; Talalay, P.; et al. Electrophilic tuning of the chemoprotective natural product sulforaphane. Proc. Natl. Acad. Sci. USA 2010, 107, 9590–9595. [Google Scholar] [CrossRef] [Green Version]
- Tirumalai, R.; Rajesh, K.T.; Mai, K.H.; Biswal, S. Acrolein causes transcriptional induction of phase II genes by activation of Nrf2 in human lung type II epithelial (A549) cells. Toxicol. Lett. 2002, 132, 27–36. [Google Scholar] [CrossRef]
- Randall, M.J.; Spiess, P.C.; Hristova, M.; Hondal, R.J.; Vander, A. Acrolein-induced activation of mitogen-activated protein kinase signalling is mediated by alkylation of thioredoxin reductase and thioredoxin1. Redox Biol. 2013, 1, 265–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poganik, J.R.; Aye, Y. Electrophilic signaling and emerging immune and neuromodulatory electrophilic pharmaceuticals. Front. Aging Neurosci. 2020, 12, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ploger, M.; Sendker, J.; Langer, K.; Schmidt, T.J. Covalent modification of human serum albumin by the natural sesquiterpene lactone partheolide. Molecules 2015, 20, 6211–6223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.C.N.; Rosenbach, M.D.; Corr, M.; Schwab, R.B. Selectivity of electrophilic compounds in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2010, 107, 7479–7484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghantous, A.; Gali-Muhtasib, H.; Vuorela, H.; Salibba, N.A.; Darwiche, N. What made sequiterpene lactones reach cancer clinical trials? Drug Discov. Today 2010, 15, 668–678. [Google Scholar] [CrossRef]
- Zhang, Q.; Chu, X.; Jiang, L.; Liu, M.; Mei, Z.; Zhang, H. Identification of non-electrophilic NRF2 activators from approved drugs. Molecules 2017, 22, 883. [Google Scholar] [CrossRef] [Green Version]
- Moujir, L.; Callies, O.; Sousa, P.M.C.; Sharopov, F.; Seca, A.M.L. Applications of sequiterpene lactones: A review of sow potential success cases. Appl. Sci. 2020, 10, 3001. [Google Scholar] [CrossRef]
- Gach, K.; Dlugosz, A.; Janecka, A. The role of oxidative stress in anticancer activity of sesquiterpene lactones. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 477–486. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Obermair, B.; Dorn, T.; Sims, K.; Rimbach, G.; Birringer, M. Sesquiterpene lactone composition and cellular Nrf2 induction of Taraxacum officinale leaves and roots and taraxinic acid β-d-glucopyranosyl ester. J. Med. Food 2017, 20, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Umemura, K.; Itoh, T.; Hamada, N.; Fujita, Y.; Ako, Y.; Nozawa, Y.; Matsuura, N.; Linuma, M.; Ito, M. Preconditioning by sesquiterpene lactone enhances H2O2-induced NRF2/ARE activation. Biochem. Biophys. Res. Commun. 2008, 368, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Fischedick, J.T.; Standford, M.; Johoson, D.A.; De Vos, R.C.H.; Todorovic, S.; Banjanac, T.; Verpoorte, R.; Johnson, J.A. Activation of antioxidant response element in mouse primary cortical cultures with sesquiterpene lactose isolated from Tanaetum parthenium. Planta Med. 2012, 78, 1725–1730. [Google Scholar] [PubMed] [Green Version]
- Formisano, C.; Rigano, D.; Millan, E.; Munoz, E.; Taglialatela-Scatati, O. Anti-inflammatory sesquiterpene lactones from Onopordum illyricum L. (Asteraceae) an Italian medicinal plant. Fitoterapia 2017, 116, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Nodwell, M.B.; Menz, H.; Kirsch, S.F.; Sieber, S.A. Rugulactone and its analogue exert antibacterial effects through multiple mechanisms including inhibition of thiamine biosynthesis. ChemBioChem 2012, 13, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Pareek, A.; Suthur, M.; Rathore, G.S.; Bansal, V. Feverfe (Tanacetum parthenium L): A systematic review. Pharmacogn. Rev. 2011, 5, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, M.; McGonnor, A.; DiNatale, G.J.; Chiramaneqong, T.; Cai, T.; Connor, R.E. Hsp72 is an intracellular target of the α,β-unsaturated sesquiterpene lactone, parthenolide. ACS Omega 2017, 2, 7267–7274. [Google Scholar] [CrossRef] [Green Version]
- Kupchan, S.M.; Fessler, D.C.; Eakin, M.A.; Giacobbe, T.J. Reactions of alpha methylene lactone tumor inhibitors with model biological neucleophiles. Science 1970, 168, 376–378. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Kang, B.; Hong, J.; Choi, H.-S. Parthenolide inhibits lipid accumulation via activation of NRF2/KEAP1 signaling during adipocyte differentiation. Food Sci. Biotechnol. 2019, 29, 431–440. [Google Scholar] [CrossRef]
- Kim, C.Y.; Kang, B.; Suh, H.J.; Choi, H.-S. Parthenolide, a fever few-derived phytochemical, ameliorates obesity and obesity-indvied infleammatory responses via the NRF2/Keap pathway. Pharmacol. Res. 2019, 145, 104259. [Google Scholar] [CrossRef]
- Mao, W.; Zhu, Z. Parthenolide inhibirs hydrogen peroxide-induced osteoblast apoptosis. Mol. Med. Rep. 2018, 17, 8369–8376. [Google Scholar] [PubMed] [Green Version]
- Uchi, H.; Arrighi, J.F.; Aubry, J.P.; Furue, M.; Hauser, C. The sesquiterpene lactone parthenolide inhibits LSP- but not TNF-alpha induced maturation of human monocyte-derived dendritic cells by inhibition of the P38 mitogen-activated protein kinase pathway. J. Allergy Clin. Immunol. 2002, 110, 269–276. [Google Scholar] [CrossRef]
- Hwang, D.; Fischer, N.H.; Jang, B.C.; Tak, H.; Kim, J.K. Inhibition of the expression of inducible cyclooxygenase and proinflammatory cytokines by sesquiterpene lactones in microphags correlates with the inhibition of MAP Kinases. Biochem. Biophys. Res. Commun. 1996, 226, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Bostwick, D.-G.; Alexander, E.E.; Singj, R.; Shan, A.; Qian, J.; Santella, R.M.; Oberley, L.W.; Yan, T.; Zhong, W.; Jiang, X.; et al. Antioxidant enzyme expression and reactive oxygen species damage in prostatic intraepithelial neoplasia and cancer. Cancer 2000, 89, 123–134. [Google Scholar] [CrossRef]
- Dey, S.; Sarkar, M.; Giri, B. Anti-inflammatory and anti-tumor activities of parthenolide: An update. J. Chem. Biol. Ther. 2016, 1, 107. [Google Scholar] [CrossRef] [Green Version]
- Li-Weber, M.; Palfi, K.; Giaisi, M.; Krammer, P.H. Dual role of the anti-inflammatory sesquiterpene lactone: Regulation of life and death by parthenolide. Cell. Death Differ. 2005, 12, 408–409. [Google Scholar] [CrossRef]
- Carlisi, D.; De Blasio, A.; Drago-Ferrante, R.; Fiore, R.D.; Buttitta, G.; Morreale, M.; Scerri, C.; Vento, R.; Tosoriere, G. Parthenolide Prevents resistance of MDA-MB231 cells to doxorubicin and mitoxantrone: The role of NRF2. Cell. Death Discov. 2017, 3, 17078. [Google Scholar] [CrossRef] [Green Version]
- Carlisi, D.; Buttitta, R.; Di Fiore, R.; Scerri, C.; Drago-Ferrante, R.; Vento, R.; Tesoriere, G. Parthenolide and DMAPT exert cytotoxic oxidative stress, mitochondrial dysfunction and necrosis. Cell. Death Dis. 2016, 7, e2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriplani, P.; Guarve, K. Recent patents on anti-cancer potential of helenalin. Recent Pat. Anticancer Drug Discov. 2020, 15, 132–142. [Google Scholar] [CrossRef]
- Shoaib, M.; Shah, I.; Ali, N.; Adhikari, A.; Tahir, M.N.; Shah, S.W.A.; Ishtiag, S.; Khan, J.; Khan, S.; Umer, M.N. Sesquiterpene lactone! A promising antioxidant, anticancer and moderate antinociceptive agent from artemesia macrocephala jacquem. BMC Complement. Altern. Med. 2017, 17, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyss, G.; Knorre, A.; Schmidt, T.J.; Pahl, H.L.; Merfort, I. The anti-inflammatory sesquiterpene lactone helenaline inhibits the transcription factor NF-kapper B by directly targeting p65. J. Biol. Chem. 1998, 273, 33508–33516. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zeng, Y.; Huang, Q.; Wen, S.; Wei, Y.; Chen, Y.; Zhang, X.; Bai, F.; Lu, Z.; Wei, J.; et al. Helenalin from centipede minima ameliorates acute hepatic injury by protecting mitochondria function, activating NRF2 pathway and inhibiting NF-kB activation. Biomed. Pharmacother. 2019, 119, 109435. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, S.; Huang, R.; Wei, L.; Tan, S.; Liang, S.; Tian, Y.; Wu, X.; Lu, Z.; Huang, Q. Helenalin enhancing ethanol metabolism, inhibiting oxidative stress and suppressiong HSC activation. Fitoterapia 2014, 95, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Li, R.-J.; Cheng, M.S.; Kim, Y.S. Alantolactone, a sesquiterpene lactone isolated from Inula helenium L. Selectrively suppress STAT3 actyivation and exhibits anticancer activity in MDA-MB-231 cells. Cancer Lett. 2015, 357, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahn, M.; Li, T.; Ahmad, K.M.K.; Rasul, A.; Nawaz, F.; Sun, M.; Zheng, Y.; Ma, T. Alantolactone induces apoptosis in HepG2 cells though GSH depletion, inhibition of STAT3 activation, and mitochondrial dysfunction. Biomed. Res. Int. 2013, 2013, 719858. [Google Scholar]
- Huo, J.; Shi, H.M.; Li, W.W.; Wang, M.Y.; Li, X.B. HPLC determination and NMR structural elucidatrion of sesquiterpene lactones in Inula helenuim. J. Pharm. Biomed. Anal. 2010, 51, 942–946. [Google Scholar] [CrossRef]
- Liu, J.; Liu, M.; Wang, S.; He, Y.; Huo, Y.; Yang, Z.; Cao, X. Alntolactone induces apoptosis and suppresses migration in MCF-7 human breast cancer cells via the p38 MAPK, NF-kB and NRF2 signaling pathways. Int. J. Mol. Med. 2018, 42, 1847–1856. [Google Scholar] [PubMed] [Green Version]
- Seo, J.Y.; Lim, S.S.; Kim, J.R.; Lim, J.-S.; Ha, Y.R.; Lee, I.A.; Kim, E.J.; Park, J.H.Y.; Kim, J.-S. NRF2-mediated Induction of detoxifying enxymes by alantolactone present in Invla helenium. Phytother. Res. 2008, 22, 1500–1505. [Google Scholar] [CrossRef]
- Dang, X.; Holt, B.; Ning, Q.; Liu, Y.; Guo, J.; Niu, G.; Chen, M. Alantolactone suppresses inflammation, apoptosis and oxidative stress in cigarette smoke-induced human bronchial epithelial cells through activation of NRF2/HO-1 and inhibition of the NF-kB pathways. Respirat. Res. 2020, 21, 95. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Choi, B.Y. Costunolide, a bioactive sesquiterpene lactone with diverse therapentic potential. Int. J. Mol. Sci. 2019, 20, 2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.; Yi, M.; Wang, R.; Huang, Y.; Chen, M. Protective effects of costunolide against D-galactos-amine and lipopolysaccharide-induced acute liver injury in mice. Front. Pharmacol. 2018, 9, 1469. [Google Scholar] [CrossRef] [Green Version]
- Pae, H.O.; Jeong, G.-S.; Kim, H.-S.; Woo, H.Y.; Rhew, H.S.; Kim, D.H.; Sohn, Y.C.; Kim, H.T.; Chung, H.-T. Costunolide inhibits production of tumor necrosis factor- K and interleukin-6 by inducing heme oxygenase-1 in RAW 264-7 Microphages. Inflamm. Res. 2007, 56, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Eliza, J.; Daisy, P.; Ignacimuthu, S. Antioxidant activity of costunolide and eremanthin isolated from costus speciosus (Ken ex. Retz) sm. Chem. Biol. Interact. 2010, 188, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chen, Y.; Zhang, J.; Wang, L.; Jin, Z.; Huang, H.; Man, S.; Gao, W. Evaluation of costunolide and dehydrocostulactone on ethano-induced gastric ulcer in mice based on multi-pathway regulation. Chem. Biol. Interact. 2016, 250, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Hou, Y.; Yao, J.; Fang, J. Activation of NRF2 by costunolide provides neuroprotective effect in PC12 Cells. Food Funct. 2019, 10, 4143. [Google Scholar] [CrossRef]
- Mythri, R.B.; Bharath, M.M. Curcumin: A potential neuroprotective agent in Parkinson’s disease. Curr. Pharm. Des. 2012, 18, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunnumakkara, A.B.; Bordoloi, D.; Padmavathi, G.; Monisha, J.; Roy, N.K.; Prasad, S.; Aggarwal, B.B. Curcumin, the golden neutraceutical: Multitargeting for multiple chronic disease. Br. J. Pharmacol. 2017, 174, 1325–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahban, M.; Habibi-Rezaei, M.; Mazaheri, M.; Saso, L.; Moosavi-Movahedi, A.A. Anti-viral potential and modulation of NRF2 by curcumin: Pharmacological implications. Antioxidants 2020, 9, 1228. [Google Scholar] [CrossRef] [PubMed]
- Madden, S.K.; Itzhaki, L.S. Structural and mechanistic insights into the KEAP1-NRF2 system as a route to drug discovery. Biochim. Et. Biophys. Acta (BBA)-Proteins Proteom. 2020, 1868, 140405. [Google Scholar] [CrossRef]
- Park, J.-Y.; Sohn, H.-Y.; Koh, Y.H.; Jo, C. Curcumin activates NRF2 through PKCᴕ-mediated p62 phosphorylation at Ser351. Sci. Rep. 2021, 11, 8430. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Ahmadi, Z.; Mohammadinejad, R.; Farkhondeh, T.; Samarghandian, S. Curcumin activates the NRF2 pathway and induces cellular protection against oxidative injury. Curr. Mol. Med. 2020, 20, 116–133. [Google Scholar] [CrossRef]
- Gonzalez-Reyes, S.; Guzman-Beltran, S.; Medina-Campos, O.N.; Pedraza-Chaverri, J. Curcumin Pretreatment induces NRF2 and an antioxidant response and prevents Hemin-induced toxicity in primary cultures of cerebellar granule neurons of rats. Oxid. Med. Cell. Longev. 2013, 2013, 801418. [Google Scholar] [CrossRef]
- Ren, L.; Zhan, P.; Wang, Q.; Wang, C.; Liu, Y.; Yu, Z.; Zhang, S. Curcumin unregulates the NRF2 system by repressing inflammatory signalling-mediated KEAP1 expression in insulin-resistant conditions. Biochem Biophys Res. Commun. 2019, 514, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Oh, J.-M.; Choi, H.; Kim, S.W.; Kim, B.G.; Cho, J.H.; Lee, J.; Lee, D.C. Activation of the NRF2/HO-1 pathway by curcumin inhibits oxidative stress in human nasal fibroblasts exposed to urban particulate matter. BMC Compl. Med. Ther. 2020, 20, 101. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Paital, B.; Jena, S.; Swain, S.S.; Kumar, S.; Yadav, M.K.; Chainy, G.B.N.; Samanta, L. Possible activation of NRF2 by vitamin E/curcumin against altered thyroid hormone induced oxidative stress via NFkB/AKT/Mtor/kap1 signaling in rat heart. Sci. Rep. 2019, 9, 7408. [Google Scholar] [CrossRef]
- Straus, D.S.; Glass, C.K. Cyclopentenone prostaglandins: New insights on biological activities and cellular targets. Med. Res. Rev. 2001, 21, 185–210. [Google Scholar] [CrossRef]
- Kim, E.H.; Surh, Y.J. 15deoxy-D12,14-prostaglandin J2 as a potential endogeneous regulator of redox-sensitive transcription factors. Biochem. Pharmacol. 2006, 72, 1516–1528. [Google Scholar] [CrossRef] [PubMed]
- Song, N.-Y.; Kim, E.-H.; Na, H.-K.; Surh, Y.-J. 15-Deoxy-delta 12, 14-prostaglandin J2 induces upregulation of multidrug resistance-associated protein 1 via NRF2 activation in human breast cancer cells. Natural compounds and their role in apoptotic cell signaling pathways. Am. N. Y. Acad. Sci. 2009, 1171, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Egner, P.A.; Wakabayashi, J.; Wakabayashi, N.; Yamamoto, M.; Kensler, T.W. NRF2-mediated induction of cytoprotective enzymes by 15-deoxyD1214 –prostaglandin J2 is attenuated by alkenal/one oxidoreductase. J. Biol. Chem. 2006, 281, 26245–26252. [Google Scholar] [CrossRef] [Green Version]
- Ugwu, D.I.; Ezema, B.E.; Okoro, U.C.; Eze, F.U.; Ekoh, O.C.; Egbujor, M.C.; Ugwuja, D. 1. ChemInform Abstract: Synthesis and pharmacological applications of chalcones: A review. ChemInform 2015, 13, 459–500. [Google Scholar] [CrossRef]
- Constantinescu, T.; Lungu, C.N. Anticancer activity of natural and synthetic chalcones. Int. J. Mol. Sci. 2021, 22, 11306. [Google Scholar] [CrossRef]
- Tang, Y.-L.; Zheng, X.; Qi, Y.; Pu, X.-J.; Liu, B.; Zhang, X.; Li, X.-S.; Xiao, W.-L.; Wan, C.-P.; Mao, Z.-W. Synthesis and anti-inflammatory evaluation of new chalcone derivatives bearing bispiperazine linker as IL-1β inhibitors. Bioorganic Chem. 2020, 98, 103748. [Google Scholar] [CrossRef]
- Ngameni, B.; Cedric, K.; Mbaveng, A.T.; Erdogan, M.; Simo, I.; Kuete, V.; Dastan, A. Design, synthesis, characterization, and anticancer activity of a novel series of O-Substituted chalcone derivatives. Bioorganic Med. Chem. Lett. 2021, 35, 127827. [Google Scholar] [CrossRef] [PubMed]
- Miranda-sapla, M.M.; Tomiotto-Pellissier, F.; Assolini, J.P.; Carloto, A.C.M.; Bortoleti, B.T.D.S.; Goncalves, M.D.; Tavares, E.R.; Rodrigues, J.H.D.S.; Simao, A.N.C.; Yamauchi, L.M.; et al. Trans-chalcone modulates Leishmania amazonensis infection in vitro by NRF2 over expression affecting iron availability. Eur. J. Pharmacol. 2019, 853, 275–288. [Google Scholar] [CrossRef]
- Kuhnl, J.; Roggenkamp, D.; Gehrke, S.A.; Stab, F.; Wenck, H.; Kolbe, L.; Neufang, G. Lichochalcone A activates NRF2 in vitro and contributes to licorice extract-induced lowered cutaneous oxidative stress. Exp. Damatol. 2015, 24, 42–47. [Google Scholar]
- Zhang, M.; Wu, Y.-Q.; Xie, L.; Wu, J.; Xu, K.; Xiao, J.; Chen, D.Q. Isoliquiritigenin protects against pancreatic injury and intestinal dysfunction after severe acute pancreatitis via NRF2 signaling. Front. Pharmacol. 2018, 9, 936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.-L.; Yang, T.-Y.; Gowrisankar, Y.V.; Liao, C.-H.; Liao, J.-W.; Huang, P.-J.; Hseu, Y.-C. Suppression of LPS-induced inflammation by chalcone flavokawain A through activation of NRF2/ARE-mediated antioxidant genes and inhibition of ROS/NFkB signaling pathways in primary splenocytes. Oxid. Med. Cell. Longev. 2020, 2020, 3476212. [Google Scholar] [CrossRef] [PubMed]
- Bomprezzi, R. Dimethyl fumarate in the treatment of relapsing-remitting multiple sclerosis: An overview. Ther. Adv. Neurol. Disord. 2015, 8, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Bista, P.; Zeng, W.; Ryan, S.; Lukashev, M.; Yamamoto, M. Dimethyl fumarate suppresses inflammation in vitro via both NRF2-dependent and NRF2-independent pathways. Neurology 2012, 78, P02.108. [Google Scholar] [CrossRef]
- Akino, N.; Wada-Hiraike, O.; Isono, W.; Terao, H.; Honjo, H.; Miyamoto, Y.; Tanikawa, M.; Sone, K.; Hirano, M.; Harada, M.; et al. Activation of NRF2/KEAP1 pathway by oral dimethylfumarate administration alleviates oxidative stress and age-associated infertility might be delayed in the mouse ovary. Reprod. Biol. Endocrinol. 2019, 17, 23. [Google Scholar] [CrossRef] [PubMed]
- Gopal, S.; Mikulskis, A.; Gold, R.; Fox, R.J.; Dawson, K.T.; Amaravadi, L. Evidance of activation of the NRF2 pathway in multiple sclerosis patients treated with delayed –release dimethyl fumarate in the phase 3 define and confirm studies. Mult. Scler. J. 2017, 2017, 1875–1883. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, M.; Kaidery, N.A.; Yang, L.; Calingasan, N.; Smirnova, N.; Gaisin, A.; Gaisina, I.N.; Gazaryan, I.; Hushpulian, D.M.; Kaddour-Djebbar, I.; et al. Distinct NRF2 signaling mechanisms of fumaric acid esters and their role in neuroprotection against 1-methyl-4-phenyl-1,2,3,6- tetrahydropyridine-induced experimental Parkinson’s like disease. J. Neurosci. 2016, 36, 6332–6351. [Google Scholar] [CrossRef] [Green Version]
- Rosto, M.; Testi, C.; Parisi, G.; Cortese, B.; Baiocco, P.; Di Angelantonio, S. Exploring the use of dimethyl fumarate as microglia modulator for neurodegenerative diseases treatment. Antioxidants 2020, 9, 700. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Okoro, U.C.; Egu, A.S.; Okonkwo, V.I.; Okafor, S.N.; Emeruwa, C.N.; Egwuatu, P.I.; Umeh, O.R.; Eziafakaego, M.I.; Amasiatu, I.S.; et al. Synthesis and biological evaluation of sulfamoyl carboxamide derivatives from sulfur-containing α-amino acids. Chiang Mai J. Sci. 2022, 49, 1100–1115. [Google Scholar] [CrossRef]
- Strating, J. Organic Sulfur Compounds, 1st ed.; Pergamon Press: Oxford, UK, 1961. [Google Scholar]
- Egbujor, M.C.; Okoro, U.C.; Okafor, S. Design, synthesis, molecular docking, antimicrobial and antioxidant activities of new phenylsulfamoyl carboxylic acids of pharmacological interest. Med. Chem. Res. 2019, 28, 2118–2127. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Okoro, U.C. New methionine-based p-toluenesulphonamoyl carboxamide derivatives as antimicrobial and antioxidant agents: Design, synthesis. J. Pharm. Res. Int. 2019, 28, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Egbujor, M.C.; Okoro, U.C.; Okafor, S.; Nwankwo, N.E. Design, synthesis and molecular docking of novel serine-based sulphonamide bioactive compounds as potential antioxidant and antimicrobial agents. Attach. Am. J. Pharm. Sci. 2019, 6, 12232–12240. [Google Scholar]
- Abdel-Aziz, A.A.; Angeli, A.; El-Azab, A.S.; Hammouda, M.E.A.; El-Sherbeny, M.A.; Supuran, C.T. Synthesis and anti-inflammatory activity of sulphonamides and carboxylates incorporating trimellitimides: Dual cyclooxygenase/carbonic anhydrase inhibitory actions. Bioorganic Chem. 2019, 84, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Simpkins, N.S. Sulfone in Organic Synthesis, 1st ed.; Pergamon Press: Oxford, UK, 1993. [Google Scholar]
- Forristal, I. The chemistry of α,β-unsaturated sulfoxides and sulfones: An update. J. Sulfur Chem. 2005, 26, 163–195. [Google Scholar] [CrossRef]
- Palmer, J.T.; Rasnick, D.; Klaus, J.L.; Bromme, D. Vinyl sulfones as mechanism-based cysteine protease inhibitor. J. Med. Chem. 1995, 38, 3193–3196. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hanzlik, R.P. Structure-activity relationship for inhibition of papain by peptide Michael acceptors. J. Med. Chem. 1992, 35, 1067–1075. [Google Scholar] [CrossRef]
- Engel, J.C.; Doyle, P.S.; Hsieh, I.; Mckerrow, J.H. Cysteine protease inhibitors cure an exprermental trypanosome cruzi infection. J. Exp. Med. 1998, 188, 725–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankel, B.A.; Bentley, M.; Kruger, R.G.; McCatterty, D.G. Vinyl sulfones: Inhibitors of SrtA, a transpeptidase required for cell wall protein anchoring and virvlence in staphylococcus aureus. J. Am. Chem. Soc. 2004, 126, 3404–3405. [Google Scholar] [CrossRef] [PubMed]
- Roush, W.R.; Gwaltney, S.L.; Cheng, J.; Scheidt, K.A.; Mckerrow, J.H.; Hansell, E. Vinyl sulfonates esters and vinyl sulphonamides: Potent, irreversible inhibitors of cystein proteases. J. Am. Chem. Soc. 1998, 120, 10994–10995. [Google Scholar] [CrossRef]
- Kover, G.E.; Kam, C.M.; Powers, J.C.; Hudig, D. Dipeptide vinyl sulfones suitable for intracellular inhibition of dipeptidyl peptidase I. Int. J. Immunopharmacol. 2001, 1, 21–32. [Google Scholar] [CrossRef]
- Kam, C.M.; Gotz, M.G.; Koot, G.; McGuire, M.; Thiele, D.; Hudig, D.; Powers, J.C. Design and evaluation of inhibitors for dipeptidyl peptidase I (Cathepsin C). Arch. Biochem. Biophys. 2004, 427, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.-L.; Hou, Y.; Bai, F.; Fang, J. Generation of potent NRF2 activators via tuning the electrophilicity and steric hinderance of vinyl sulfones for neuroprotection. Bioorganic Chem. 2020, 107, 104520. [Google Scholar] [CrossRef] [PubMed]
- Egbujor, M.C.; Okoro, U.C.; Okafor, S. Novel alanine-based antimicrobial and antioxidant agents: Synthesis and molecular docking. Indian J. Sci. Technol. 2020, 13, 1003–1014. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Okoro, U.C.; Nwobodo, D.C.; Ezeagu, C.U.; Amadi, U.B.; Okenwa-Ani, C.G.; Ugwu, J.I.; Okoye, I.G.; Abu, I.P.; Egwuatu, P.I. Design, synthesis, antimicrobial and antioxidant activities of novel threonine-based sulfonamide derivatives. J. Pharm. Res. Int. 2020, 32, 51–61. [Google Scholar] [CrossRef]
- Onoabedje, E.A.; Ibezim, A.; Okoro, U.C.; Batra, S. Synthesis, molecular docking, antiplasmodial and antioxidant activities of new sulfonamido-peptode derivatives. Heliyon 2020, 6, e04958. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Okoro, U.C.; Egu, S.A.; Egwuatu, P.I.; Eze, F.U.; Amasiatu, I.S. Design, synthesis and biological evaluation of alanine-based sulphonamide derivatives. Int. J. Res. Pharm. Sci. 2020, 11, 6449–6458. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Okoro, U.C.; Okafor, S.; Nwankwo, N.E. Synthesis, characterization and in silico studies of novel alkanoylated 4-methylphenyl sulphonamoyl carboxylic acids as potential antimicrobial and antioxidant agents. Int. J. Pharm. Phytopharm. Res. 2019, 9, 89–97. [Google Scholar]
- Egbujor, M.C.; Egu, S.A.; Okonkwo, V.I.; Jacob, A.D.; Egwuatu, P.I.; Amasiatu, I.S. Antioxidant drug design:historical and recent developments. J. Pharm. Res. Int. 2021, 32, 36–56. [Google Scholar] [CrossRef]
- Egbujor, M.C.; Okoro, U.C.; Okafor, S.N.; Amasiatu, I.S.; Amadi, U.B.; Egwuatu, P.I. Synthesis, molecular docking and pharmacological evaluation of new 4-methylphenylsulphamoyl carboxylic acids analogs. Int. J. Res. Pharm. Sci. 2020, 11, 5357–5366. [Google Scholar] [CrossRef]
- Luan, F.; Wei, L.; Zhang, J.; MI, Y.; Dong, F.; Li, Q.; Guo, Z. Autioxidant activity of chitosan derivatives with propane sulfonate groups. Polymer 2018, 10, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arshia, A.J.; Faheem, A.; Khan, K.M.; Shah, S.; Perveen, S. Benzophenone esters and sulfonates synthesis and their potential as anti-inflammatory agents. Med. Chem. 2019, 15, 162–174. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, T.L. Synthesis: Carbon with three or four attached heteroatoms. In Comprehensive Organic Functional Group Transformations; Elsevier Science: Oxford, UK, 2017; Volume 6. [Google Scholar]
- Otacka, S.; Kwiatknowska, M.; Madalinska, L.; Kielbasinski, P. Chiral organosulfur ligands/catalysts with a stereogenic sulfur atom: Applications in asymmetric synthesis. Chem. Rev. 2017, 117, 414–4181. [Google Scholar] [CrossRef] [PubMed]
- Elisia, I.; Nakamura, H.; Lam, V.; Hofs, E.; Cederberg, R.; Cait, J.; Hughes, M.R.; Lee, L.; Jia, W.; Adomat, H.H.; et al. DMSO represses inflammatory cytokine production from human blood cells and reduces autoimmune arthritis. PLoS ONE 2016, 11, e0152538. [Google Scholar]
- Sanmartin-Suarez, C.; Soto-Otero, R.; Sanchez-Sellero, I.; Mendez-Alvarez, E. Antioxidant properties of dimethyl sulfoxide and its viability as a solvent in the evaluation of neuroprotective antioxidants. J. Pharmacol. Toxicol. Methods 2011, 63, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Forristal, I.; Rayner, C.M. Advances in Sulfur Chemistry. In Recent Advances in the Chemistry of α,β-Unsaturated Sulfoxides and Sulfones; Rayner, C.M., Ed.; JAI Press: Greenwich, CT, USA, 2000; Volume 2, pp. 155–213. [Google Scholar]
- Posner, G.H. Asymmetric Synthesis; Morrison, J., Ed.; Academic Press: New York, NY, USA, 1983; Volume 2, p. 225. [Google Scholar]
- Lyzwa, P.; Jankowiak, A.; Kwiatowska, M.; Mikolajczyk, M.; Kielbasinski, P.; Betz, A.; Jaffres, P.-A.; Gavmont, A.-C.; Gulea, M. Diastereoselective Michael additions to α,β-unsaturated sulfinyl phosphonates in the thiolane series. Tetrahedron Lett. 2007, 48, 351–355. [Google Scholar] [CrossRef]
- Unnikrishria, M.K.; Rao, M.N.A. Anti-inflammatory activity of methionine, methionine sulfoxide and methionone sulfone. Agents Actions 1990, 31, 110–112. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, D. Sesquiterpene Lactones: Diversity and Their Biological Activities in Opportunity, Challenge and Scope of Natural Products in Medicinal Chemistry; Tiwari, V.K., Mishra, B.B., Eds.; Research Signpost: Thiruvananthapuram, India, 2011; pp. 313–334. [Google Scholar]
- Ivanescu, B.; Miron, A.; Corciova, A. Sesquiterpene lactones from Artemisia genus: Biological activities and methods of analysis. J. Anal. Meth. Chem. 2015, 2015, 247685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cesare Mannelli, L.; Tenci, B.; Zanardelli, M.; Maidecchi, A.; Lugli, A.; Mattoli, L.; Ghelardini, C. Widespread pain reliever profile of a flower extract of Tanacetum parthenium. Phytomedicine 2015, 22, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.L.; Rossi, R.M.; Neelakantan, S.; Li, X.; Corbrtt, C.A.; Hassane, D.C.; Becker, M.W.; Bennett, J.M.; Sullivan, E.; Lachowicz, J.L.; et al. An orally bioavailable parthenolide analog selectively eradicates acute myelogenous leukemia stem and progenitor cells. Blood 2007, 110, 4427–4435. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Kinghorn, D. Development of potential antitumor agents from the scaffolds of plant-derived terpenoid lactones. J. Med. Chem. 2020, 63, 15410–15448. [Google Scholar] [CrossRef]
- Janecka, A.; Wyrebska, A.; Gach, K.; Fichna, J.; Janecki, T. Natural and synthetic α-methylenelactams with anticancer potential. Drug Discov. Today 2012, 17, 561–572. [Google Scholar] [CrossRef]
- Hall, I.H.; Lee, K.H.; Starnes, C.O.; Muraoka, O.; Sumida, Y.; Waddell, T.G. Antihyper lipidemic activity of sesquiterpene lactones and related compounds. J. Pharm. Sci. 1980, 69, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Witzell, D.A.; Ivie, W.; Dollahite, J.W. Mammalian toxicity of helenalin, the toxic principle of Helenium microcephalum CD (smallhead sneezeweed). Am. J. Vet. Res. 1976, 37, 859–861. [Google Scholar]
- Rumack, B.H. POISINDEX(R) Information System Micromedex; Inc., CCIS: Englewood, CO, USA, 2017; Volume 172. [Google Scholar]
- Hall, A.H.; Rumack, B.H. TOMES(R) Information System Micromedex; Inc., CCIS: Englewood, CO, USA, 2017; Volume 172. [Google Scholar]
- Khan, M.; Yi, F.; Rasul, A.; Li, T.; Wang, N.; Gao, R.; Ma, T. Alantolactone induces apoptosis in glioblastoma cells via GSH depletion, ROS generation and mitochondrial dysfunction. IUBMB Life 2012, 64, 783–794. [Google Scholar] [CrossRef]
- Sandberg, M.; Patil, J.; D’Angelo, B.; Weber, S.G.; Mallard, C. NRF2-regulation in brain health and disease: Implication of cerebral inflammation. Neuropharmacology 2014, 79, 298–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, C.; Zhang, S.; Teng, S.; Niu, K. Simultaneous determination of sesquiterpene lactones isoalantolactone and alantolactone isomers in rat plasma by liquid chromatography with tandem mass spectrometry: Application to a pharmacokinetic study. J. Sep. Sci. 2014, 37, 950–956. [Google Scholar] [CrossRef]
- Xu, R.; Zhou, G.; Peng, Y.; Wang, M.L.; Li, X. Pharmacokinetics, tissue distribution and excretion of isoalantolactone and alantolactone in rats after oral administration of radix inulae extract. Molecules 2015, 20, 7719–7736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Ye, J.; Yang, N.; Chen, L.; Zhuo, Z.; Mao, L.; Liu, Q.; Lan, G.; Ning, J.G.; Ge, G.; et al. Metabolism and pharmacokinetics of alantolactone and isoalantolactone in rats: Thiol conjugation as a potential metabolic pathway. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2018, 1072, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.; Tower, G.H.N.; Mitchell, J.C. Biological activities of sesquiterpene lactones. Phytochemistry 1976, 15, 1573–1580. [Google Scholar] [CrossRef]
- Liu, Q.; Majdi, M.; Cankar, K.; Goedbloed, M.; Charnikhova, T.; Verstappen, F.W.A.; De vos, R.C.H.; Beekwilder, J.; Van der Krol, S.; Bouwmeester, H.J. Reconstitution of the costunolide biosynthetic pathway in yeast and nicotiana benthamiana. PLoS ONE 2011, 6, e23255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hu, X.; Gao, W.; Qu, Z.; Guo, H.; Liu, Z.; Liu, C. Pharmacokinetic study of costunolide and dehydrocostus lactone after oral administration of traditional medicine Aucklandic lappa Dcne by LC/MS/MS. J. Ethnopharmacol. 2014, 151, 191–197. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, Y.; Gu, X.; Yang, C. Study on the the pharmacokinetics and metabolism of costunolide and hydocostus lactone in rats by HPLC-UV and UPLC-Q-TOF/MS. Biomed. Chromatogr. 2014, 28, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Chen, O.; Ray, S.D.; Ji, J.; Bucci, L.R.; Preuss, H.G. Highly bioavailable forms of curcumin and promising avenues for curcumin-based research and application: A review. Molecules 2020, 25, 1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Ji, J.; Bucci, L.R.; Preuss, R.G. A comparative pharmacokinetic assessment of novel highly bioavailable curcumin formulation with 95% curcumin: A randomized, double-blind, cross-over study. J. Am. Coll. Nutr. 2018, 37, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Schuster, V.L.; Chi, Y.; Lu, R. The prostaglandin transporter: Eicosanoid reuptake, control of signaling and development of high affinity inhibitors as drug candidates. Trans. Am. Clin. Climatol. Assoc. 2015, 126, 248–257. [Google Scholar] [PubMed]
- Razeghinejad, M.R. The effect of latanaprost on intraocular inflammation and macular edema. Ocul. Immunol. Inflamm. 2019, 27, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Akkinapally, S.; Hundalani, S.G.; Kulkarni, M.; Fernandes, C.J.; Cabrera, A.G.; Shivanna, B.; Pammi, M. Prostaglandin E1 for maintaining ductual potency in neonate with ductual-dependent cardiac lesions. Cochrane Database Syst Rev. 2018, 27, CD011417. [Google Scholar]
- Krishna, U.; Ganguli, A.C.; Mandlekar, A.V.; Purandane, V.N. Administration of prostaglandins by various routes for induction of abortion, merits and demerits. Prostanglandins 1978, 15, 685–693. [Google Scholar] [CrossRef]
- Malik, K.; Dua, A. Prostaglandins. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Abbasi, S.; Kajimoto, K.; Harashima, H. Elimination of the biphasic pharmacodynamics of 15d-PGJ2 by controlling its release from a nanoemulsion. Int. J. Nanomed. 2016, 11, 2685–2694. [Google Scholar]
- Emi, M.; Maeyema, K. The biphasic effects of cyclopentenone prostaglandins, prostaglandin J(2) and 15-deoxy-Delta(12,14)-prostaglandin J(2) on proliferation and apoptosis in rat basophilic leukemia (RBL-2H3) cells. Biochem. Pharmacol. 2004, 67, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Napimoga, M.H.; da Siva, C.A.; Carregaro, V. Exogenous administration of 15d-PGJ2-loaded nanocapsules inhibit bone resorption in a mouse periodontitis model. J. Immunol. 2012, 189, 1043–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homen de Bittencourt, P.I., Jr.; Lagranha, D.J.; Maslinkiewicz, A.; Senna, S.M.; Tavares, A.M.; Baldissera, L.P.; Janner, D.D.; Peralta, J.S.; Bock, P.M.; Gutierrez, L.L.P.; et al. Lipocardium endothelium-directed cyclopentenone prostaglandin-based liposome formulation that completely reverse atherosclerotic lesions. Atherosclerosis 2007, 193, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Hagens, W.L.; Mattos, A.; Greupink, R. Targeting 15d-prostaglandin J2 to hepatic stellates cells: Two options evaluated. Pharm. Res. 2007, 24, 566–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A. Chalcone: A valuable insight into the recent advances and potential pharmacological activities. Chem. Sci. J. 2011, 2011, CSJ-29. [Google Scholar] [CrossRef]
- Sinha, S.; Prakash, A.; Medhi, B. Pharmacokinetic evaluation of chalcone derivatives with antimalarial activity in New Zealand white rabbits. BMC Res Notes. 2021, 14, 264. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.; Beshbishy, A.M.; Tayebwa, D.S.; Adeyemi, O.S.; Shaheen, H.; Yokoyama, N.; Igarashi, I. The effects of trans-chalcone and chalcone 4 hydrate on the growth of Babesia and Theileria. PloS Negl Trop Dis. 2019, 13, e0007030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancino, K.; Castro, I.; Yauri, C.; Julian, V.; Arevalo, J.; Sauvain, M.; Adaul, V.; Castillo, D. Toxicity assessment of synthetic chalcones with antileishmanial potential in BALB/c mice. Rev. Peru. Med. Exp. Salud Publica 2021, 38, 424–433. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. The Chemical-Biological Coordination Centre of the National Research Council; The national Academies Press: Washington, DC, USA, 1953. [Google Scholar] [CrossRef] [Green Version]
- Weng, Q.; Chen, L.; Ye, L.; Lu, X.; Yu, Z.; Wen, C.; Chen, Y.; Huang, G. Determination of licochalcone A in rat plasma by UPLC-MS/MS and its pharmacokinetics. Acta Chromatogr. 2019, 31, 262–265. [Google Scholar] [CrossRef]
- Cao, M.; Zhan, M.; Wang, Z.; Li, X.M.; Miao, M. Development of orally bioavailable isoliquiritigenin self-nanoemulsifying drug delivery system to effectively treat ovalbumin-induced asthma. Int. J. Nanomed. 2020, 15, 8945–8961. [Google Scholar] [CrossRef]
- Xu, X.; Liu, Z.; Li, X.; Liu, S.; Zi, X. Abstract 4629: Ultra performance liquid chromatography (UPLC)-tandem mass spectrometry (MS/MS) method for analysis and pharmacokinetic study of flavokawain A, a novel chalcone from the kava plant, in mice. Cancer Res. 2011, 71, 4629. [Google Scholar]
- Sweetser, M.T.; Dawson, K.T.; Bozic, C. Manufacturer’s response to case reports of PML. N. Engl. J. Med. 2013, 368, 1659–1661. [Google Scholar] [CrossRef]
- Li, L.; Tsai, S.-W.; Anderson, A.-L.; Keire, D.A.; Raubitschek, A.A.; Shively, J.E. Vinyl sulfone bifunctional derivatives of DOTA allow sulfhydryl- or amino-directed coupling to antibodies. Conjugates retain immunoreactivity and have similar biodistributions. Bioconjugate Chem. 2002, 13, 110–115. [Google Scholar] [CrossRef]
- Smyth, H.F.; Carpenter, C.P.; Well, C.S.; Pozzani, U.C.; Striegel, J.A. Range-finding toxicity data: List VI. AIHA 2007, 23, 95–107. [Google Scholar] [CrossRef]
- McKerrow, J.H.; Rosenthal, P.J.; Swenerton, R.; Doyle, P. Development of protease inhibitors for protozoan infections. Curr. Opin. Infect. Dis. 2008, 21, 668–672. [Google Scholar] [CrossRef] [PubMed]
- Renslo, A.R.; McKerrow, J.H. Drug discovery and development for neglected parasiticdisease. Nat. Chem. Biol. 2006, 2, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Prado, M.A.F. Determination of lipophilicity constants of sulfonamide substituents. J. Chem. Educ. 2001, 78, 533. [Google Scholar] [CrossRef]
- Mita, M.M.; Mita, A.C.; Goldston, M.; Chu, O.S.; Tolcher, A.W.; Desai, A.R.; Okereke, C.S.; Rowinsky, E.K.; Takimoto, C. Pharmacokinetics(PK) and pharmacodynamics (PD) of E7820 an oral sulfonamide with novel, alpha-2 integrin mediated antiangiogenic properties: Results of a phase 1 study. J. Clin. Oncol. 2005, 23, 3082. [Google Scholar] [CrossRef]
- Vree, T.B.; Nouws, J.F.M.; Hekster, Y.A. Comparative Pharmacokinetic Studies of Sulphonamides. In Comparative Veterinary Pharmacology, Toxicology and Therapy; Van Miert, A.S.J.P.A.M., Bogaert, M.G., Debackere, M., Eds.; Springer: Dordrecht, The Netherlands, 1986. [Google Scholar] [CrossRef]
- Jing, P.; Rodgers, P.J.; Amemiya, S. High lipophilicity of perfluoroalkyl carboxylate and sulfonate: Implications for their membrane permeability. J. Am. Chem. Soc. 2009, 131, 2290–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neelakantan, S.; Nasim, S.; Guzmzn, M.L.; Jordan, C.T.; Crooks, P.A. Aminoparthenolides as novel anti-leukemic agents: Discovery of the NF-KappaB inhibitor, DMAPT (LC-1). Bioorganic Med. Chem. Lett. 2009, 19, 4346–4349. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.S.; Pae, H.O.; Jeong, S.O.; Kim, Y.C.; Kwon, T.O.; Lee, H.S.; Kim, N.S.; Park, S.D.; Chung, H.T. The alpha-methyl-gamma-butyrolactone moiety in dehydrocostus lactone is responsible for cytoprotective heme oxygenase-1 expression through activation of the nuclear factor E2-related factor 2 in HepG2 Cells. Eur. J. Pharmacol. 2007, 565, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.R.; Mo, H.; Bieberich, A.A.; Alavanja, T.; Colby, D.A. Fluorinated amino-derivatives of the sesquiterpene lactone, parthenolide as (19)f NMR probes in deuterium-free environments. J. Med. Chem. 2011, 54, 7934–7941. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Payne, D.T.; Ampolu, B.; Bland, N.; Brown, J.T.; Dutton, M.J.; Fitton, C.A.; Gulliver, A.; Hale, L.; Hamza, D.; et al. Derivatisation of parthenolide to address chemoresistant chronic lymphocytic leukaemia. MedChemComm 2019, 10, 1379–1390. [Google Scholar] [CrossRef] [Green Version]
- Beekman, A.C.; Woerdenbag, H.J.; van Uden, W.; Pras, N.; Konings, A.W.T.; Wikstrom, H.V.; Schmidt, T.J. Structure-cytotoxicity relationships of some helenanolide type sesquiterpene lactones. J. Nat. Prodt. 1997, 60, 252–257. [Google Scholar] [CrossRef]
- Liu, X.-N.; Li, H.-M.; Wang, S.-P.; Zhang, J.-Z.; Liu, D.-L. Sesquiterpene lactones of Aucklandia lappa: Pharmacology, pharmacokinetics, toxicity, and structure-activity relationship. Chin. Herb. Med. 2021, 13, 167–176. [Google Scholar] [CrossRef]
- Kumar, V.; Kumar, S.; Hassan, M.; Wu, H.; Thimmulappa, R.K.; Kumar, A.; Sharma, S.K.; Parmar, V.S.; Biswal, S.; Malhotra, S.V. Novel Chalcone Derivatives as potent NRF2 activators in mice and human lungs epithelial cells. J. Med. Chem. 2011, 54, 4147–4159. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, L.; Cheng, C.; Li, G.; Xie, J.; Shen, M.; Chen, Q.; Li, W.; He, W.; Qiu, P.; et al. Design, synthesis and biological evaluation of chalcone analogues with novel dual antioxidant mechanisms as potential anti-ischemic stroke agents. Acta Pharm. Sin. B 2019, 9, 335–350. [Google Scholar] [CrossRef]
- Sorrenti, V.; Vanella, L.; Plantania, C.B.M.; Greish, K.; Bucolo, C.; Pittala, V.; Salerno, L. Novel heme oxygenase-1(HO-1) inducers based on dimethyl fumarate structure. Int. J. Mol. Sci. 2020, 21, 9541. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egbujor, M.C.; Buttari, B.; Profumo, E.; Telkoparan-Akillilar, P.; Saso, L. An Overview of NRF2-Activating Compounds Bearing α,β-Unsaturated Moiety and Their Antioxidant Effects. Int. J. Mol. Sci. 2022, 23, 8466. https://doi.org/10.3390/ijms23158466
Egbujor MC, Buttari B, Profumo E, Telkoparan-Akillilar P, Saso L. An Overview of NRF2-Activating Compounds Bearing α,β-Unsaturated Moiety and Their Antioxidant Effects. International Journal of Molecular Sciences. 2022; 23(15):8466. https://doi.org/10.3390/ijms23158466
Chicago/Turabian StyleEgbujor, Melford Chuka, Brigitta Buttari, Elisabetta Profumo, Pelin Telkoparan-Akillilar, and Luciano Saso. 2022. "An Overview of NRF2-Activating Compounds Bearing α,β-Unsaturated Moiety and Their Antioxidant Effects" International Journal of Molecular Sciences 23, no. 15: 8466. https://doi.org/10.3390/ijms23158466
APA StyleEgbujor, M. C., Buttari, B., Profumo, E., Telkoparan-Akillilar, P., & Saso, L. (2022). An Overview of NRF2-Activating Compounds Bearing α,β-Unsaturated Moiety and Their Antioxidant Effects. International Journal of Molecular Sciences, 23(15), 8466. https://doi.org/10.3390/ijms23158466