
Sea Anemones, Actinoporins, and Cholesterol

 ,
,  , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Historical Context
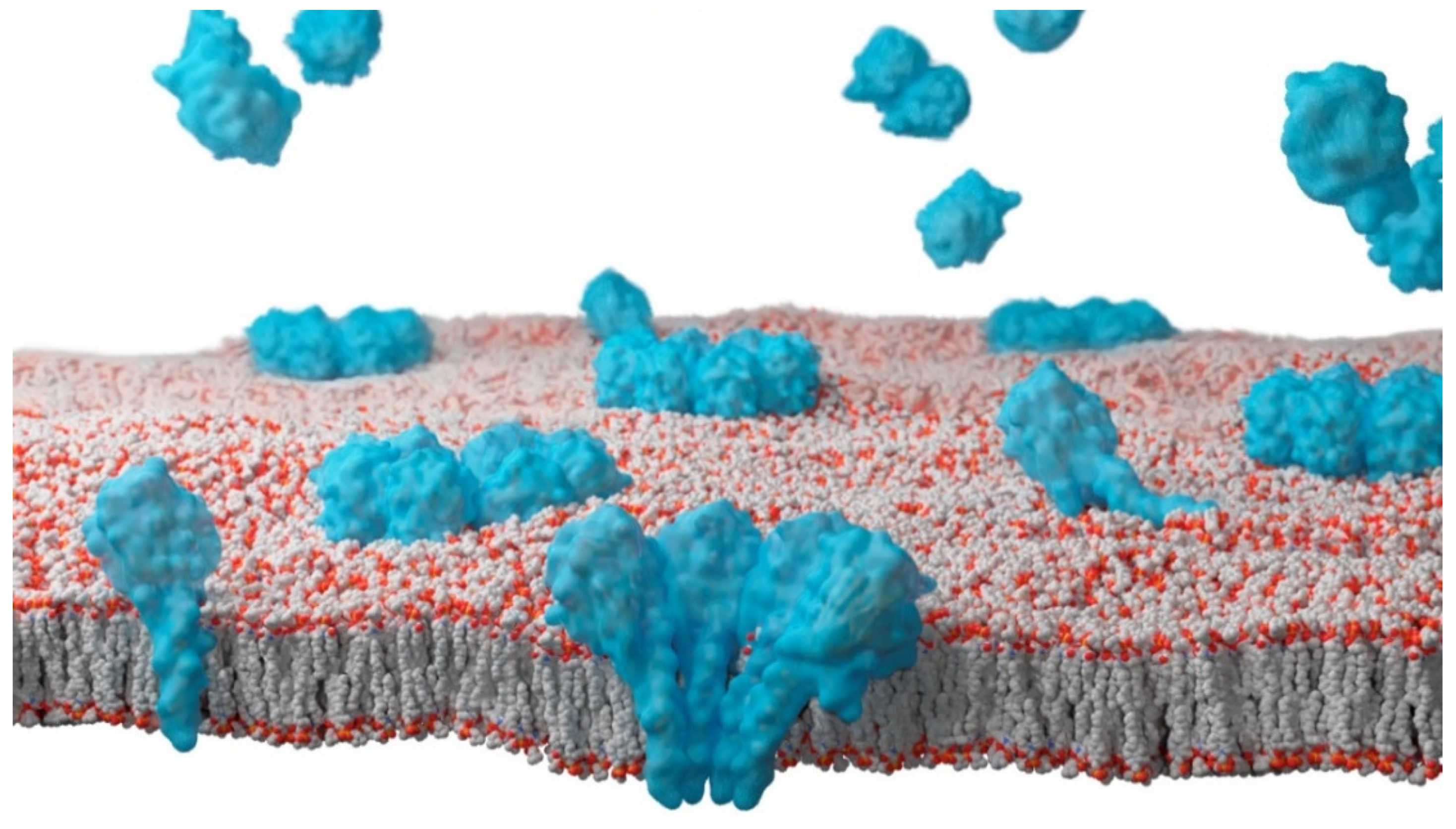
2. Sea Anemones Actinoporins
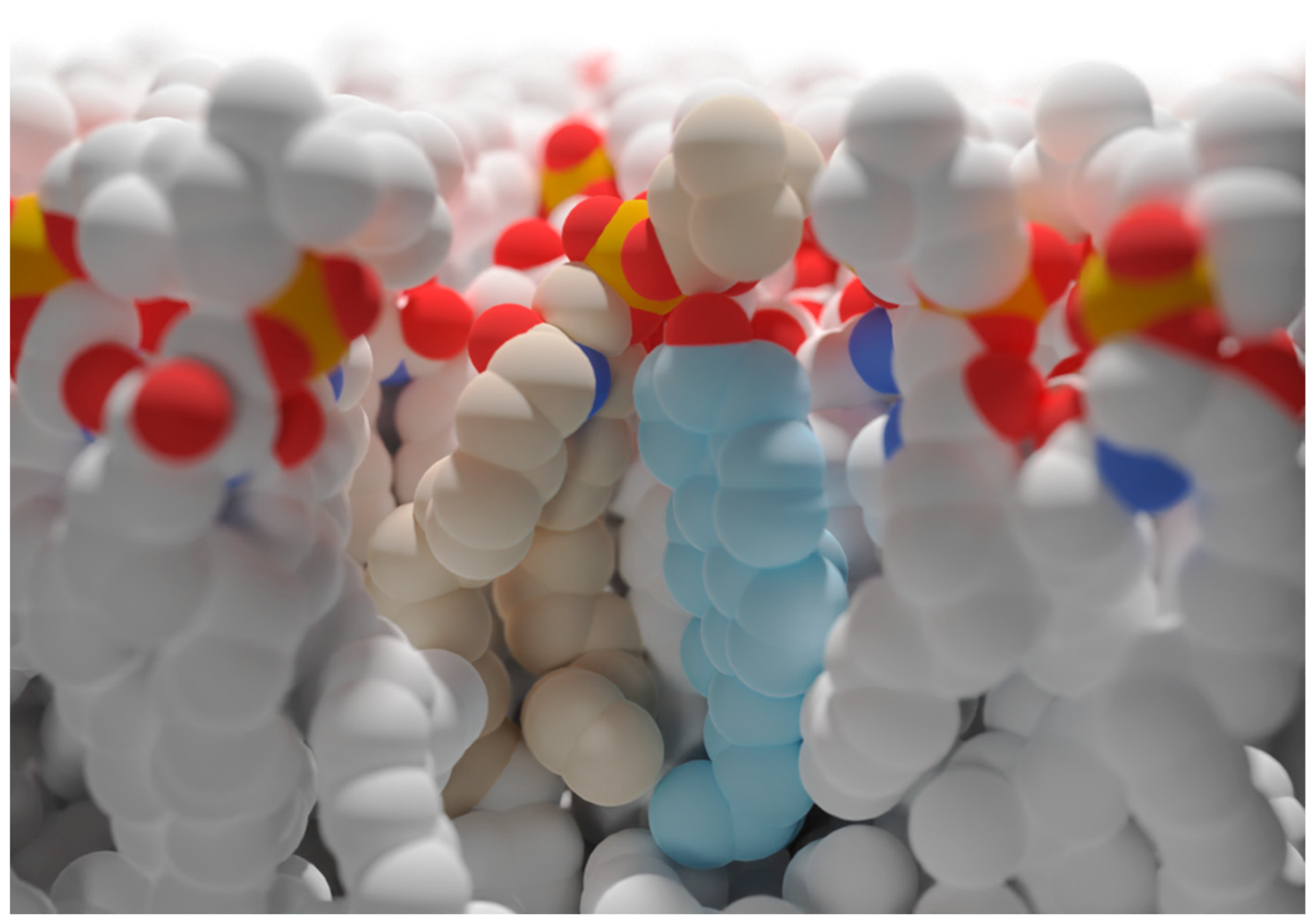
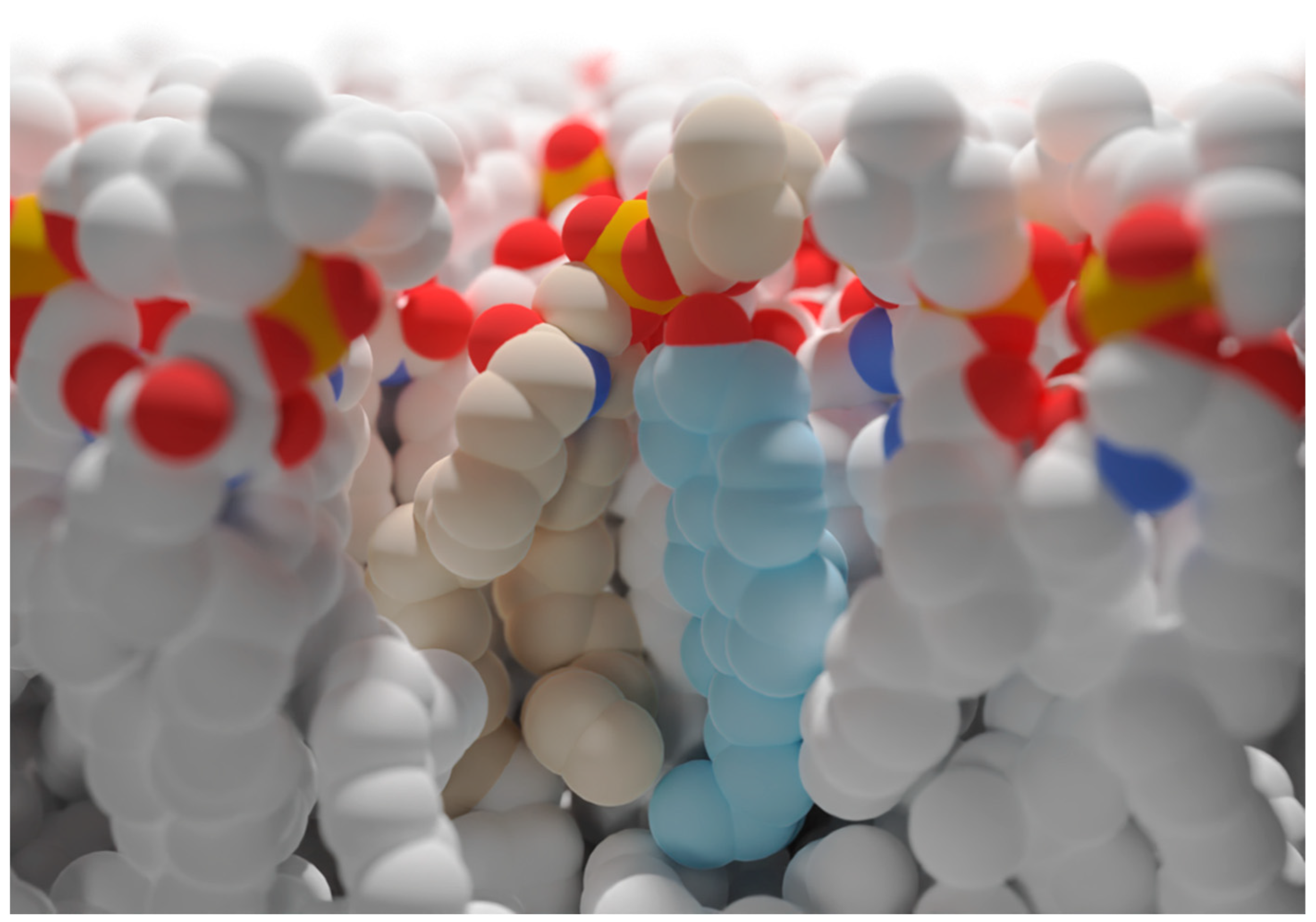
3. The Role of Cholesterol
4. Perspectives and Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Chol | cholesterol |
| Eqt | equinatoxin |
| Fra | fragaceatoxin |
| OlyA | Ostreolysin A |
| OSM | Oleoyl-sphingomyelin |
| PC | phosphatidylcholine |
| PSM | palmitoyl-sphingomyelin |
| SM | sphingomyelin |
| Stns | sticholysins |
References
- Pazos, I.F.; Alvarez, C.; Lanio, M.E.; Martinez, D.; Morera, V.; Lissi, E.A.; Campos, A.M. Modification of sticholysin II hemolytic activity by free radicals. Toxicon 1998, 36, 1383–1393. [Google Scholar] [CrossRef]
- Alvarez, C.; Lanio, M.E.; Tejuca, M.; Martínez, D.; Pazos, F.; Campos, A.M.; Encinas, M.V.; Pertinhez, T.; Schreier, S.; Lissi, E.A. The role of ionic strength on the enhancement of the hemolytic activity of sticholysin I, a cytolysin from Stichodactyla helianthus. Toxicon 1998, 36, 165–178. [Google Scholar] [CrossRef]
- Tejuca, M.; Anderluh, G.; Maček, P.; Marcet, R.; Torres, D.; Sarracent, J.; Alvarez, C.; Lanio, M.E.; Dalla Serra, M.; Menestrina, G. Antiparasite activity of sea-anemone cytolysins on Giardia duodenalis and specific targeting with anti-Giardia antibodies. Int. J. Parasitol. 1999, 29, 489–498. [Google Scholar] [CrossRef]
- Campos, A.M.; Lissi, E.A.; Vergara, C.; Lanio, M.E.; Alvarez, C.; Pazos, I.; Morera, V.; García, Y.; Martínez, D. Kinetics and mechanism of St I modification by peroxyl radicals. J. Protein Chem. 1999, 18, 297–306. [Google Scholar] [CrossRef]
- Álvarez, C.; Mancheño, J.M.; Martínez, D.; Tejuca, M.; Pazos, F.; Lanio, M.E. Sticholysins, two pore-forming toxins produced by the Caribbean sea anemone Stichodactyla helianthus: Their interaction with membranes. Toxicon 2009, 54, 1135–1147. [Google Scholar] [CrossRef]
- Alvarez, C.; Soto, C.; Cabezas, S.; Alvarado-Mesen, J.; Laborde, R.; Pazos, F.; Ros, U.; Hernandez, A.M.; Lanio, M.E. Panorama of the Intracellular Molecular Concert Orchestrated by Actinoporins, Pore-Forming Toxins from Sea Anemones. Toxins 2021, 13, 567. [Google Scholar] [CrossRef]
- Caaveiro, J.M.; Echabe, I.; Gutiérrez-Aguirre, I.; Nieva, J.L.; Arrondo, J.L.; González-Mañas, J.M. Differential interaction of equinatoxin II with model membranes in response to lipid composition. Biophys. J. 2001, 80, 1343–1353. [Google Scholar] [CrossRef] [Green Version]
- Barlič, A.; Gutiérrez-Aguirre, I.; Caaveiro, J.M.; Cruz, A.; Ruiz-Argüello, M.B.; Pérez-Gil, J.; González-Mañas, J.M. Lipid phase coexistence favors membrane insertion of equinatoxin-II, a pore-forming toxin from Actinia equina. J. Biol. Chem. 2004, 279, 34209–34216. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Aguirre, I.; Barlič, A.; Podlesek, Z.; Maček, P.; Anderluh, G.; González-Mañas, J.M. Membrane insertion of the N-terminal α-helix of equinatoxin II, a sea anemone cytolytic toxin. Biochem. J. 2004, 384, 421–428. [Google Scholar] [CrossRef]
- Kristan, K.; Podlesek, Z.; Hojnik, V.; Gutiérrez-Aguirre, I.; Guncar, G.; Turk, D.; González-Mañas, J.M.; Lakey, J.H.; Maček, P.; Anderluh, G. Pore Formation by Equinatoxin, a Eukaryotic Pore-forming Toxin, Requires a Flexible N-terminal Region and a Stable β-Sandwich. J. Biol. Chem. 2004, 279, 46509–46517. [Google Scholar] [CrossRef] [Green Version]
- Martínez, D.; Otero, A.; Álvarez, C.; Pazos, F.; Tejuca, M.; Lanio, M.E.; Gutiérrez-Aguirre, I.; Barlič, A.; Iloro, I.; Arrondo, J.L.; et al. Effect of sphingomyelin and cholesterol on the interaction of St II with lipidic interfaces. Toxicon 2007, 49, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Bellomio, A.; Morante, K.; Barlič, A.; Gutiérrez-Aguirre, I.; Viguera, A.R.; González-Mañas, J.M. Purification, cloning and characterization of fragaceatoxin C, a novel actinoporin from the sea anemone Actinia fragacea. Toxicon 2009, 54, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Mechaly, A.E.; Bellomio, A.; Morante, K.; González-Mañas, J.M.; Guerin, D.M. Crystallization and preliminary crystallographic analysis of fragaceatoxin C, a pore-forming toxin from the sea anemone Actinia fragacea. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2009, 65, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Caaveiro, J.M.; Morante, K.; González-Mañas, J.M.; Tsumoto, K. Structural basis for self-assembly of a cytolytic pore lined by protein and lipid. Nat. Commun. 2015, 6, 6337. [Google Scholar] [CrossRef] [Green Version]
- Flores-Romero, H.; Ros, U.; Garcia-Saez, A.J. Pore formation in regulated cell death. Embo J. 2020, 39, e105753. [Google Scholar] [CrossRef]
- Subburaj, Y.; Ros, U.; Hermann, E.; Tong, R.; García-Sáez, A.J. Toxicity of an α-pore-forming toxin depends on the assembly mechanism on the target membrane as revealed by single-molecule imaging. J. Biol. Chem. 2015, 290, 4856–4865. [Google Scholar] [CrossRef] [Green Version]
- Ros, U.; García-Sáez, A.J. More than a pore: The interplay of pore-forming proteins and lipid membranes. J. Membr. Biol. 2015, 248, 545–561. [Google Scholar] [CrossRef]
- Cosentino, K.; Ros, U.; García-Sáez, A.J. Assembling the puzzle: Oligomerization of α-pore forming proteins in membranes. Biochim. Biophys. Acta 2016, 1858, 457–466. [Google Scholar] [CrossRef]
- Tejuca, M.; Serra, M.D.; Ferreras, M.; Lanio, M.E.; Menestrina, G. Mechanism of membrane permeabilization by sticholysin I, a cytolysin isolated from the venom of the sea anemone Stichodactyla helianthus. Biochemistry 1996, 35, 14947–14957. [Google Scholar] [CrossRef]
- Schön, P.; García-Sáez, A.J.; Malovrh, P.; Bacia, K.; Anderluh, G.; Schwille, P. Equinatoxin II permeabilizing activity depends on the presence of sphingomyelin and lipid phase coexistence. Biophys. J. 2008, 95, 691–698. [Google Scholar] [CrossRef] [Green Version]
- García-Sáez, A.J.; Buschhorn, S.B.; Keller, H.; Anderluh, G.; Simons, K.; Schwille, P. Oligomerization and pore formation by Equinatoxin II inhibit endocytosis and lead to plasma membrane reorganization. J. Biol. Chem. 2011, 286, 37768–37777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morante, K.; Caaveiro, J.M.; Tanaka, K.; González-Mañas, J.M.; Tsumoto, K. A Pore-Forming Toxin Requires a Specific Residue for its Activity in Membranes with Particular Physicochemical Properties. J. Biol. Chem. 2015, 290, 1850–1861. [Google Scholar] [CrossRef] [Green Version]
- Morante, K.; Caaveiro, J.M.; Viguera, A.R.; Tsumoto, K.; González-Mañas, J.M. Functional characterization of Val60, a key residue involved in the membrane-oligomerization of fragaceatoxin C, an actinoporin from Actinia fragacea. FEBS Lett. 2015, 589, 1840–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Caaveiro, J.M.; Tsumoto, K. Bidirectional Transformation of a Metamorphic Protein between the Water-Soluble and Transmembrane Native States. Biochemistry 2015, 54, 6863–6866. [Google Scholar] [CrossRef] [PubMed]
- Morante, K.; Bellomio, A.; Gil-Carton, D.; Redondo-Morata, L.; Sot, J.; Scheuring, S.; Valle, M.; Gonzalez-Manas, J.M.; Tsumoto, K.; Caaveiro, J.M.M. Identification of a Membrane-bound Prepore Species Clarifies the Lytic Mechanism of Actinoporins. J. Biol. Chem. 2016, 291, 19210–19219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kupferschmidt, K. From toxins to treatments. Science 2013, 342, 1162–1164. [Google Scholar] [CrossRef]
- Holford, M.; Daly, M.; King, G.F.; Norton, R.S. Venoms to the rescue. Science 2018, 361, 842–844. [Google Scholar] [CrossRef]
- von Reumont, B.M.; Anderluh, G.; Antunes, A.; Ayvazyan, N.; Beis, D.; Caliskan, F.; Crnkovic, A.; Damm, M.; Dutertre, S.; Ellgaard, L.; et al. Modern venomics-Current insights, novel methods, and future perspectives in biological and applied animal venom research. Gigascience 2022, 11. [Google Scholar] [CrossRef]
- King, G.F.; Hardy, M.C. Spider-venom peptides: Structure, pharmacology, and potential for control of insect pests. Annu. Rev. Entomol. 2013, 58, 475–496. [Google Scholar] [CrossRef]
- Galenkamp, N.S.; Van Meervelt, V.; Mutter, N.L.; van der Heide, N.J.; Wloka, C.; Maglia, G. Preparation of Cytolysin A (ClyA) Nanopores. Methods Mol. Biol. 2021, 2186, 11–18. [Google Scholar] [CrossRef]
- Lucas, F.L.R.; Versloot, R.C.A.; Yakovlieva, L.; Walvoort, M.T.C.; Maglia, G. Protein identification by nanopore peptide profiling. Nat. Commun. 2021, 12, 5795. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, E. Untangling spider biology. Science 2017, 358, 288–291. [Google Scholar] [CrossRef] [PubMed]
- King, G.F. Venoms as a platform for human drugs: Translating toxins into therapeutics. Expert Opin Biol. 2011, 11, 1469–1484. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.H. Composition and mode of action of some invertebrate venoms. Annu. Rev. Pharmacol. 1964, 4, 293–304. [Google Scholar] [CrossRef]
- Lane, C. Toxins of marine origin. Annu. Rev. Pharmacol. 1968, 8, 409–426. [Google Scholar] [CrossRef]
- Baslow, M.H. Marine toxins. Annu. Rev. Pharmacol. 1971, 11, 447–454. [Google Scholar] [CrossRef]
- Steele, R.E.; David, C.N.; Technau, U. A genomic view of 500 million years of cnidarian evolution. Trends Genet. 2011, 27, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Technau, U.; Steele, R.E. Evolutionary crossroads in developmental biology: Cnidaria. Development 2011, 138, 1447–1458. [Google Scholar] [CrossRef] [Green Version]
- Maček, P. Polypeptide cytolytic toxins from sea anemones (Actiniaria). FEMS Microbiol. Immunol. 1992, 5, 121–129. [Google Scholar] [CrossRef]
- Alegre-Cebollada, J.; Oñaderra, M.; Gavilanes, J.G.; Martínez-del-Pozo, A. Sea anemone actinoporins: The transition from a folded soluble state to a functionally active membrane-bound oligomeric pore. Curr. Protein Pept. Sci. 2007, 8, 558–572. [Google Scholar] [CrossRef] [Green Version]
- García-Ortega, L.; Alegre-Cebollada, J.; García-Linares, S.; Bruix, M.; Martínez-del-Pozo, A.; Gavilanes, J.G. The behavior of sea anemone actinoporins at the water-membrane interface. Biochim. Biophys. Acta 2011, 1808, 2275–2288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Linares, S.; Rivera-de-Torre, E.; Palacios-Ortega, J.; Gavilanes, J.G.; Martínez-del-Pozo, A. The metamorphic transformation of a water-soluble monomeric protein into an oligomeric transmembrane pore. Adv. Biomembr. Lipid Self-Assem. 2017, 26, 51–97. [Google Scholar]
- Rivera-de-Torre, E.; Palacios-Ortega, J.; Gavilanes, J.G.; Martínez-del-Pozo, A.; García-Linares, S. Pore-Forming Proteins from Cnidarians and Arachnids as Potential Biotechnological Tools. Toxins 2019, 11, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-de-Torre, E.; Palacios-Ortega, J.; Slotte, J.P.; Gavilanes, J.G.; Martínez-del-Pozo, A.; García-Linares, S. Functional and Structural Variation among Sticholysins, Pore-Forming Proteins from the Sea Anemone Stichodactyla helianthus. Int. J. Mol. Sci. 2020, 21, 8915. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Ortega, J.; García-Linares, S.; Rivera-de-Torre, E.; Heras-Márquez, D.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, A. Structural foundations of sticholysin functionality. Biochim. Biophys. Acta Proteins Proteom. 2021, 1869, 140696. [Google Scholar] [CrossRef]
- Palacios-Ortega, J.; Rivera-de-Torre, E.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, A.; García-Linares, S. Biophysical approaches to study actinoporin-lipid interactions. Methods Enzymol. 2021, 649, 307–339. [Google Scholar] [CrossRef]
- Anfinsen, C.B. Studies on the principles that govern the holding of protein chains. Science 1973, 181, 223–230. [Google Scholar] [CrossRef] [Green Version]
- De los Ríos, V.; Mancheño, J.M.; Martínez-del-Pozo, A.; Alfonso, C.; Rivas, G.; Oñaderra, M.; Gavilanes, J.G. Sticholysin II, a cytolysin from the sea anemone Stichodactyla helianthus, is a monomer-tetramer associating protein. FEBS Lett. 1999, 455, 27–30. [Google Scholar] [CrossRef] [Green Version]
- García-Linares, S.; Amigot-Sánchez, R.; García-Montoya, C.; Heras-Márquez, D.; Alfonso, C.; Luque-Ortega, J.R.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Palacios-Ortega, J. Sticholysin I-II oligomerization in the absence of membranes. FEBS Lett. 2022, 596, 1029–1036. [Google Scholar] [CrossRef]
- De los Ríos, V.; Mancheño, J.M.; Lanio, M.E.; Oñaderra, M.; Gavilanes, J.G. Mechanism of the leakage induced on lipid model membranes by the hemolytic protein sticholysin II from the sea anemone Stichodactyla helianthus. Eur. J. Biochem. 1998, 252, 284–289. [Google Scholar] [CrossRef]
- Rivera-de-Torre, E.; Martínez-del-Pozo, A.; Garb, J.E. Stichodactyla helianthus’ de novo transcriptome assembly: Discovery of a new actinoporin isoform. Toxicon 2018, 150, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Rivera-de-Torre, E.; Palacios-Ortega, J.; Garb, J.E.; Slotte, J.P.; Gavilanes, J.G.; Martínez-del-Pozo, A. Structural and functional characterization of sticholysin III: A newly discovered actinoporin within the venom of the sea anemone Stichodactyla helianthus. Arch. Biochem. Biophys. 2020, 689, 108435. [Google Scholar] [CrossRef] [PubMed]
- Ferlan, I.; Lebez, D. Equinatoxin, a lethal protein from Actinia equina-I Purification and characterization. Toxicon 1974, 12, 57–58. [Google Scholar] [CrossRef]
- Maček, P.; Lebez, D. Isolation and characterization of three lethal and hemolytic toxins from the sea anemone Actinia equina L. Toxicon 1988, 26, 441–451. [Google Scholar] [CrossRef]
- Morante, K.; Bellomio, A.; Viguera, A.R.; González-Mañas, J.M.; Tsumoto, K.; Caaveiro, J.M.M. The Isolation of New Pore-Forming Toxins from the Sea Anemone Actinia fragacea Provides Insights into the Mechanisms of Actinoporin Evolution. Toxins 2019, 11, 401. [Google Scholar] [CrossRef] [Green Version]
- Devlin, J.P. Isolation and partial purification of hemolytic toxin from sea anemone, Stoichactis helianthus. J. Pharm. Sci. 1974, 63, 1478–1480. [Google Scholar] [CrossRef]
- Bernheimer, A.W.; Avigad, L.S. Properties of a toxin from the sea anemone Stoichactis helianthus, including specific binding to sphingomyelin. Proc. Natl. Acad. Sci. USA 1976, 73, 467–471. [Google Scholar] [CrossRef] [Green Version]
- Linder, R.; Bernheimer, A.W.; Kim, K.S. Interaction between sphingomyelin and a cytolysin from the sea anemone Stoichactis helianthus. Biochim. Biophys. Acta 1977, 467, 290–300. [Google Scholar] [CrossRef]
- Monastyrnaya, M.; Leychenko, E.; Isaeva, M.; Likhatskaya, G.; Zelepuga, E.; Kostina, E.; Trifonov, E.; Nurminski, E.; Kozlovskaya, E. Actinoporins from the sea anemones, tropical Radianthus macrodactylus and northern Oulactis orientalis: Comparative analysis of structure-function relationships. Toxicon 2010, 56, 1299–1314. [Google Scholar] [CrossRef]
- García-Linares, S.; Rivera-de-Torre, E.; Morante, K.; Tsumoto, K.; Caaveiro, J.M.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, Á. Differential effect of membrane composition on the pore-forming ability of four different sea anemone actinoporins. Biochemistry 2016, 55, 6630–6641. [Google Scholar] [CrossRef]
- Khoo, K.S.; Kam, W.K.; Khoo, H.E.; Gopalakrishnakone, P.; Chung, M.C. Purification and partial characterization of two cytolysins from a tropical sea anemone, Heteractis magnifica. Toxicon 1993, 31, 1567–1579. [Google Scholar] [CrossRef]
- Wang, Y.; Yap, L.L.; Chua, K.L.; Khoo, H.E. A multigene family of Heteractis magnificalysins (HMgs). Toxicon 2008, 51, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Tkacheva, E.S.; Leychenko, E.V.; Monastyrnaya, M.M.; Issaeva, M.P.; Zelepuga, E.A.; Anastuk, S.D.; Dimitrenok, P.S.; Kozlovskaya, E.P. New actinoporins from sea anemone Heteractis crispa: Cloning and functional expression. Biochemistry 2011, 76, 1131. [Google Scholar] [CrossRef] [PubMed]
- Turk, T. Cytolytic toxins from sea anemones. Toxin Rev. 1991, 10, 223–262. [Google Scholar] [CrossRef]
- De los Ríos, V.; Oñaderra, M.; Martínez-Ruiz, A.; Lacadena, J.; Mancheño, J.M.; Martínez-del-Pozo, A.; Gavilanes, J.G. Overproduction in Escherichia coli and purification of the hemolytic protein sticholysin II from the sea anemone Stichodactyla helianthus. Protein Expr. Purif. 2000, 18, 71–76. [Google Scholar] [CrossRef]
- Athanasiadis, A.; Anderluh, G.; Maček, P.; Turk, D. Crystal structure of the soluble form of equinatoxin II, a pore-forming toxin from the sea anemone Actinia equina. Structure 2001, 9, 341–346. [Google Scholar] [CrossRef] [Green Version]
- Hinds, M.G.; Zhang, W.; Anderluh, G.; Hansen, P.E.; Norton, R.S. Solution structure of the eukaryotic pore-forming cytolysin equinatoxin II: Implications for pore formation. J. Mol. Biol. 2002, 315, 1219–1229. [Google Scholar] [CrossRef] [Green Version]
- Mancheño, J.M.; Martín-Benito, J.; Martínez-Ripoll, M.; Gavilanes, J.G.; Hermoso, J.A. Crystal and electron microscopy structures of sticholysin II actinoporin reveal insights into the mechanism of membrane pore formation. Structure 2003, 11, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- García-Linares, S.; Castrillo, I.; Bruix, M.; Menéndez, M.; Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Gavilanes, J.G. Three-dimensional structure of the actinoporin sticholysin I. Influence of long-distance effects on protein function. Arch. Biochem. Biophys. 2013, 532, 39–45. [Google Scholar] [CrossRef]
- Mechaly, A.E.; Bellomio, A.; Gil-Carton, D.; Morante, K.; Valle, M.; González-Mañas, J.M.; Guerin, D.M. Structural insights into the oligomerization and architecture of eukaryotic membrane pore-forming toxins. Structure 2011, 19, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Hong, Q.; Gutiérrez-Aguirre, I.; Barlič, A.; Malovrh, P.; Kristan, K.; Podlesek, Z.; Maček, P.; Turk, D.; González-Mañas, J.M.; Lakey, J.H.; et al. Two-step Membrane Binding by Equinatoxin II, a Pore-forming Toxin from the Sea Anemone, Involves an Exposed Aromatic Cluster and a Flexible Helix. J. Biol. Chem. 2002, 277, 41916–41924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norton, R.S. Structures of sea anemone toxins. Toxicon 2009, 54, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, D. The discovery of the α-helix and β-sheet, the principal structural features of proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 11207–11210. [Google Scholar] [CrossRef] [Green Version]
- Malovrh, P.; Viero, G.; Serra, M.D.; Podlesek, Z.; Lakey, J.H.; Maček, P.; Menestrina, G.; Anderluh, G. A novel mechanism of pore formation: Membrane penetration by the N-terminal amphipathic region of equinatoxin. J. Biol. Chem. 2003, 278, 22678–22685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alegre-Cebollada, J.; Martínez-del-Pozo, A.; Gavilanes, J.G.; Goormaghtigh, E. Infrared spectroscopy study on the conformational changes leading to pore formation of the toxin sticholysin II. Biophys. J. 2007, 93, 3191–3201. [Google Scholar] [CrossRef]
- Alegre-Cebollada, J.; Cunietti, M.; Herrero-Galán, E.; Gavilanes, J.G.; Martínez-del-Pozo, A. Calorimetric scrutiny of lipid binding by sticholysin II toxin mutants. J. Mol. Biol. 2008, 382, 920–930. [Google Scholar] [CrossRef] [Green Version]
- García-Linares, S.; Palacios-Ortega, J.; Yasuda, T.; Astrand, M.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Toxin-induced pore formation is hindered by intermolecular hydrogen bonding in sphingomyelin bilayers. Biochim. Biophys. Acta 2016, 1858, 1189–1195. [Google Scholar] [CrossRef] [Green Version]
- Palacios-Ortega, J.; García-Linares, S.; Astrand, M.; Al Sazzad, M.A.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Regulation of Sticholysin II-Induced Pore Formation by Lipid Bilayer Composition, Phase State, and Interfacial Properties. Langmuir 2016, 32, 3476–3484. [Google Scholar] [CrossRef]
- Rivera-de-Torre, E.; Palacios-Ortega, J.; García-Linares, S.; Gavilanes, J.G.; Martínez-del-Pozo, A. One single salt bridge explains the different cytolytic activities shown by actinoporins sticholysin I and II from the venom of Stichodactyla helianthus. Arch. Biochem. Biophys. 2017, 636, 79–89. [Google Scholar] [CrossRef]
- Palacios-Ortega, J.; García-Linares, S.; Rivera-de-Torre, E.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Differential Effect of Bilayer Thickness on Sticholysin Activity. Langmuir 2017, 33, 11018–11027. [Google Scholar] [CrossRef]
- Palacios-Ortega, J.; García-Linares, S.; Rivera-de-Torre, E.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Sticholysin, Sphingomyelin, and Cholesterol: A Closer Look at a Tripartite Interaction. Biophys. J. 2019, 116, 2253–2265. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Ortega, J.; Rivera-de-Torre, E.; Gavilanes, J.G.; Slotte, J.P.; Martinez-del-Pozo, A. Evaluation of different approaches used to study membrane permeabilization by actinoporins on model lipid vesicles. BBA-Biomembranes 2020, 1862, 183311. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Ortega, J.; Rivera-de-Torre, E.; García-Linares, S.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Oligomerization of Sticholysins from Forster Resonance Energy Transfer. Biochemistry 2021, 60, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.L.; Michaels, D.W.; Mayer, M.M. Membrane damage by a toxin from the sea anemone Stoichactis helianthus. II. Effect of membrane lipid composition in a liposome system. Biochim. Biophys. Acta 1979, 555, 79–88. [Google Scholar] [CrossRef]
- Belmonte, G.; Pederzolli, C.; Maček, P.; Menestrina, G. Pore formation by the sea anemone cytolysin equinatoxin-II in red blood cells and model lipid membranes. J. Membr. Biol. 1993, 131, 11–22. [Google Scholar] [CrossRef]
- Valcarcel, C.A.; Dalla Serra, M.; Potrich, C.; Bernhart, I.; Tejuca, M.; Martínez, D.; Pazos, F.; Lanio, M.E.; Menestrina, G. Effects of lipid composition on membrane permeabilization by sticholysin I and II, two cytolysins of the sea anemone Stichodactyla helianthus. Biophys. J. 2001, 80, 2761–2774. [Google Scholar] [CrossRef] [Green Version]
- Maula, T.; Isaksson, Y.J.; García-Linares, S.; Niinivehmas, S.; Pentikainen, O.T.; Kurita, M.; Yamaguchi, S.; Yamamoto, T.; Katsumura, S.; Gavilanes, J.G.; et al. 2NH and 3OH are crucial structural requirements in sphingomyelin for sticholysin II binding and pore formation in bilayer membranes. Biochim. Biophys. Acta 2013, 1828, 1390–1395. [Google Scholar] [CrossRef] [Green Version]
- Alm, I.; García-Linares, S.; Gavilanes, J.G.; Martínez-del-Pozo, A.; Slotte, J.P. Cholesterol stimulates and ceramide inhibits sticholysin II-induced pore formation in complex bilayer membranes. Biochim. Biophys. Acta -Biomembranes 2015, 1848, 925–931. [Google Scholar] [CrossRef] [Green Version]
- García-Linares, S.; Alm, I.; Maula, T.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, A. The effect of cholesterol on the long-range network of interactions established among sea anemone Sticholysin II residues at the water-membrane interface. Mar. Drugs 2015, 13, 1647–1665. [Google Scholar] [CrossRef]
- Meinardi, E.; Florin-Christensen, M.; Paratcha, G.; Azcurra, J.M.; Florin-Christensen, J. The molecular basis of the self/nonself selectivity of a coelenterate toxin. Biochem. Biophys. Res. Commun. 1995, 216, 348–354. [Google Scholar] [CrossRef]
- Varanda, W.; Finkelstein, A. Ion and nonelectrolyte permeability properties of channels formed in planar lipid bilayer membranes by the cytolytic toxin from the sea anemone, Stoichactis helianthus. J. Membr. Biol. 1980, 55, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Alegre-Cebollada, J.; Rodríguez-Crespo, I.; Gavilanes, J.G.; Martínez-del-Pozo, A. Detergent-resistant membranes are platforms for actinoporin pore-forming activity on intact cells. FEBS J. 2006, 273, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Bakrač, B.; Anderluh, G. Molecular mechanism of sphingomyelin-specific membrane binding and pore formation by actinoporins. Adv. Exp. Med. Biol. 2009, 677, 106–115. [Google Scholar]
- Pedrera, L.; Fanani, M.L.; Ros, U.; Lanio, M.E.; Maggio, B.; Álvarez, C. Sticholysin I-membrane interaction: An interplay between the presence of sphingomyelin and membrane fluidity. Biochim. Biophys. Acta 2014, 1838, 1752–1759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera-de-Torre, E.; García-Linares, S.; Alegre-Cebollada, J.; Lacadena, J.; Gavilanes, J.G.; Martínez-del-Pozo, A. Synergistic action of actinoporin isoforms from the same sea anemone species assembled into functionally active heteropores. J. Biol. Chem. 2016, 291, 14109–14119. [Google Scholar] [CrossRef] [Green Version]
- Pedrera, L.; Gomide, A.B.; Sánchez, R.E.; Ros, U.; Wilke, N.; Pazos, F.; Lanio, M.E.; Itri, R.; Fanani, M.L.; Álvarez, C. The presence of sterols favors sticholysin I-membrane association and pore formation regardless of their ability to form laterally segregated domains. Langmuir 2015, 31, 9911–9923. [Google Scholar] [CrossRef]
- Kristan, K.C.; Viero, G.; Dalla Serra, M.; Maček, P.; Anderluh, G. Molecular mechanism of pore formation by actinoporins. Toxicon 2009, 54, 1125–1134. [Google Scholar] [CrossRef]
- Ros, U.; Edwards, M.A.; Epand, R.F.; Lanio, M.E.; Schreier, S.; Yip, C.M.; Alvarez, C.; Epand, R.M. The sticholysin family of pore-forming toxins induces the mixing of lipids in membrane domains. Biochim. Biophys. Acta 2013, 1828, 2757–2762. [Google Scholar] [CrossRef] [Green Version]
- Cosentino, K.; Hermann, E.; von Kugelgen, N.; Unsay, J.D.; Ros, U.; Garcia-Saez, A.J. Force Mapping Study of Actinoporin Effect in Membranes Presenting Phase Domains. Toxins 2021, 13, 669. [Google Scholar] [CrossRef]
- Brameshuber, M.; Sevcsik, E.; Rossboth, B.K.; Manner, C.; Deigner, H.P.; Peksel, B.; Peter, M.; Torok, Z.; Hermetter, A.; Schutz, G.J. Oxidized Phospholipids Inhibit the Formation of Cholesterol-Dependent Plasma Membrane Nanoplatforms. Biophys. J. 2016, 110, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Bunge, A.; Muller, P.; Stockl, M.; Herrmann, A.; Huster, D. Characterization of the ternary mixture of sphingomyelin, POPC, and cholesterol: Support for an inhomogeneous lipid distribution at high temperatures. Biophys. J. 2008, 94, 2680–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engberg, O.; Yasuda, T.; Hautala, V.; Matsumori, N.; Nyholm, T.K.; Murata, M.; Slotte, J.P. Lipid Interactions and Organization in Complex Bilayer Membranes. Biophys. J. 2016, 110, 1563–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanashima, S.; Murakami, K.; Yura, M.; Yano, Y.; Umegawa, Y.; Tsuchikawa, H.; Matsumori, N.; Seo, S.; Shinoda, W.; Murata, M. Cholesterol-Induced Conformational Change in the Sphingomyelin Headgroup. Biophys. J. 2019, 117, 307–318. [Google Scholar] [CrossRef] [PubMed]
- García-Linares, S.; Maula, T.; Rivera-de-Torre, E.; Gavilanes, J.G.; Slotte, J.P.; Martínez-del-Pozo, A. Role of the tryptophan residues in the specific interaction of the sea anemone Stichodactyla helianthus’s actinoporin Sticholysin II with biological membranes. Biochemistry 2016, 55, 6406–6420. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Buboltz, J.T.; Feigenson, G.W. Maximum solubility of cholesterol in phosphatidylcholine and phosphatidylethanolamine bilayers. Biochim. Biophys. Acta 1999, 1417, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Bjorkbom, A.; Rog, T.; Kaszuba, K.; Kurita, M.; Yamaguchi, S.; Lonnfors, M.; Nyholm, T.K.; Vattulainen, I.; Katsumura, S.; Slotte, J.P. Effect of Sphingomyelin Headgroup Size on Molecular Properties and Interactions with Cholesterol. Biophys. J. 2010, 99, 3300–3308. [Google Scholar] [CrossRef] [Green Version]
- Jaikishan, S.; Björkbom, A.; Slotte, J.P. Sphingomyelin analogs with branched N-acyl chains: The position of branching dramatically affects acyl chain order and sterol interactions in bilayer membranes. Biochim. Biophys. Acta 2010, 1798, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Feigenson, G.W. A microscopic interaction model of maximum solubility of cholesterol in lipid bilayers. Biophys. J. 1999, 76, 2142–2157. [Google Scholar] [CrossRef] [Green Version]
- Endapally, S.; Frias, D.; Grzemska, M.; Gay, A.; Tomchick, D.R.; Radhakrishnan, A. Molecular Discrimination between Two Conformations of Sphingomyelin in Plasma Membranes. Cell 2019, 176, 1040–1053. [Google Scholar] [CrossRef] [Green Version]
- Sepcic, K.; Berne, S.; Rebolj, K.; Batista, U.; Plemenitas, A.; Sentjurc, M.; Maček, P. Ostreolysin, a pore-forming protein from the oyster mushroom, interacts specifically with membrane cholesterol-rich lipid domains. FEBS Lett. 2004, 575, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Feinstein, M.; Fernandez, S.; Sha’Afi, R. Fluidity of natural membranes and phosphatidylserine and ganglioside dispersions: Effects of local anesthetics, cholesterol and protein. Biochim. Biophys. Acta Biomembr. 1975, 413, 354–370. [Google Scholar] [CrossRef]
- Bruzik, K.S.; Sobon, B.; Salamonczyk, G.M. Nuclear magnetic resonance study of sphingomyelin bilayers. Biochemistry 1990, 29, 4017–4021. [Google Scholar] [CrossRef] [PubMed]
- Aittoniemi, J.; Rog, T.; Niemelä, P.; Pasenkiewicz-Gierula, M.; Karttunen, M.; Vattulainen, I. Tilt: Major factor in sterols’ ordering capability in membranes. J. Phys. Chem. B 2006, 110, 25562–25564. [Google Scholar] [CrossRef] [PubMed]
- Róg, T.; Pasenkiewicz-Gierula, M. Cholesterol-sphingomyelin interactions: A molecular dynamics simulation study. Biophys. J. 2006, 91, 3756–3767. [Google Scholar] [CrossRef] [Green Version]
- García-Linares, S.; Richmond, R.; García-Mayoral, M.F.; Bustamante, N.; Bruix, M.; Gavilanes, J.G.; Martínez-del-Pozo, A. The sea anemone actinoporin (Arg-Gly-Asp) conserved motif is involved in maintaining the competent oligomerization state of these pore-forming toxins. FEBS J. 2014, 281, 1465–1478. [Google Scholar] [CrossRef]
- Malovrh, P.; Barlič, A.; Podlesek, Z.; Maček, P.; Menestrina, G.; Anderluh, G. Structure-function studies of tryptophan mutants of equinatoxin II, a sea anemone pore-forming protein. Biochem. J. 2000, 346, 223–232. [Google Scholar] [CrossRef]
- Wacklin, H.P.; Bremec, B.B.; Moulin, M.; Rojko, N.; Haertlein, M.; Forsyth, T.; Anderluh, G.; Norton, R.S. Neutron reflection study of the interaction of the eukaryotic pore-forming actinoporin equinatoxin II with lipid membranes reveals intermediate states in pore formation. Biochim. Biophys. Acta 2016, 1858, 640–652. [Google Scholar] [CrossRef]
- Sepehri, A.; Nepal, B.; Lazaridis, T. Lipid interactions of an actinoporin pore-forming oligomer. Biophys. J. 2021, 120, 1357–1366. [Google Scholar] [CrossRef]
- Mutter, N.L.; Huang, G.; van der Heide, N.J.; Lucas, F.L.R.; Galenkamp, N.S.; Maglia, G.; Wloka, C. Preparation of Fragaceatoxin C (FraC) Nanopores. Methods Mol. Biol. 2021, 2186, 3–10. [Google Scholar] [CrossRef]
- Wloka, C.; Mutter, N.L.; Soskine, M.; Maglia, G. α-Helical Fragaceatoxin C Nanopore Engineered for Double-Stranded and Single-Stranded Nucleic Acid Analysis. Angew. Chem. Int. Ed. Engl. 2016, 55, 12494–12498. [Google Scholar] [CrossRef]
- de Lannoy, C.; Lucas, F.L.R.; Maglia, G.; de Ridder, D. In silico assessment of a novel single-molecule protein fingerprinting method employing fragmentation and nanopore detection. iScience 2021, 24, 103202. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Xiang, Y.; Liu, Q.; Wang, L.; Ma, Q.; Ma, W.; Zeng, D.; Yin, Y.; Wang, D. Nanopore Technology for the Application of Protein Detection. Nanomaterials 2021, 11, 1942. [Google Scholar] [CrossRef]
- Versloot, R.C.A.; Lucas, F.L.R.; Yakovlieva, L.; Tadema, M.J.; Zhang, Y.; Wood, T.M.; Martin, N.I.; Marrink, S.J.; Walvoort, M.T.C.; Maglia, G. Quantification of Protein Glycosylation Using Nanopores. Nano Lett. 2022, 22, 5357–5364. [Google Scholar] [CrossRef] [PubMed]
- Haugland, M.M.; Borsley, S.; Cairns-Gibson, D.F.; Elmi, A.; Cockroft, S.L. Synthetically Diversified Protein Nanopores: Resolving Click Reaction Mechanisms. ACS Nano 2019, 13, 4101–4110. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palacios-Ortega, J.; Heras-Márquez, D.; Amigot-Sánchez, R.; García-Montoya, C.; Torrijos, C.; Laxalde, D.; Gavilanes, J.G.; García-Linares, S.; Martínez-del-Pozo, Á. Sea Anemones, Actinoporins, and Cholesterol. Int. J. Mol. Sci. 2022, 23, 8771. https://doi.org/10.3390/ijms23158771
Palacios-Ortega J, Heras-Márquez D, Amigot-Sánchez R, García-Montoya C, Torrijos C, Laxalde D, Gavilanes JG, García-Linares S, Martínez-del-Pozo Á. Sea Anemones, Actinoporins, and Cholesterol. International Journal of Molecular Sciences. 2022; 23(15):8771. https://doi.org/10.3390/ijms23158771
Chicago/Turabian StylePalacios-Ortega, Juan, Diego Heras-Márquez, Rafael Amigot-Sánchez, Carmen García-Montoya, Carlos Torrijos, Diego Laxalde, José G. Gavilanes, Sara García-Linares, and Álvaro Martínez-del-Pozo. 2022. "Sea Anemones, Actinoporins, and Cholesterol" International Journal of Molecular Sciences 23, no. 15: 8771. https://doi.org/10.3390/ijms23158771