
Identification of Novel Regulators of Radiosensitivity Using High-Throughput Genetic Screening

 ,
, 
 and
and
Abstract
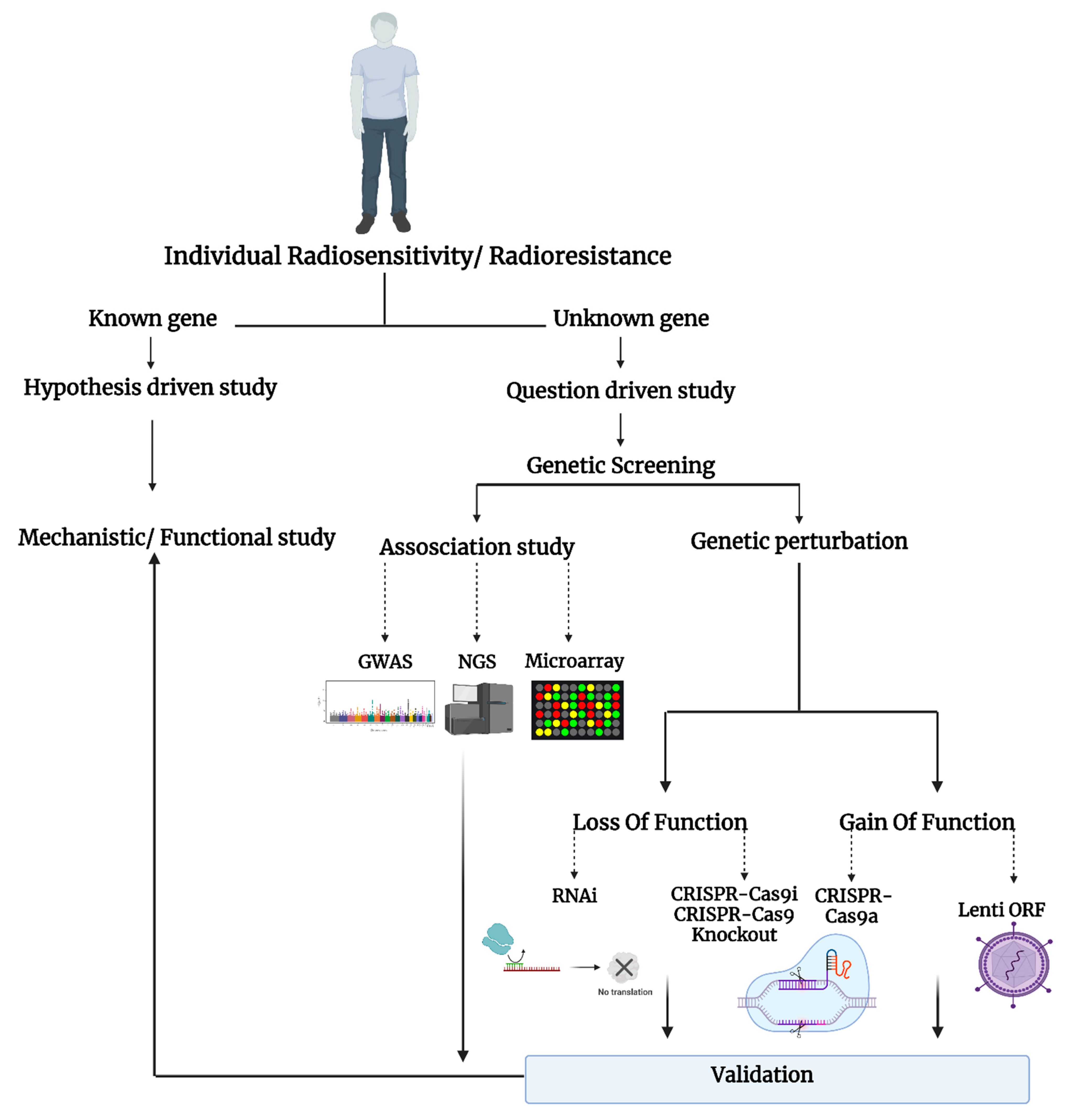
:1. Introduction
2. The Radiation Response
3. High-Throughput Screening Methods to Study Radiosensitivity and Resistance
3.1. Gene Expression Analysis (Microarray and RNA-Sequencing)
3.2. Genome-Wide Association Study (GWAS)
3.3. Genome-Wide RNAi Screening Method
3.4. Genome-Wide CRISPR-Cas9 Screening Method
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, X.; Nakano, T.; Tsuda, M.; Kanamoto, R.; Hirayama, R.; Uzawa, A.; Ide, H. Direct observation of damage clustering in irradiated DNA with atomic force microscopy. Nucleic Acids Res. 2019, 48, e18. [Google Scholar] [CrossRef] [PubMed]
- Ulyanenko, S.; Pustovalova, M.; Koryakin, S.; Beketov, E.; Lychagin, A.; Ulyanenko, L.; Kaprin, A.; Grekhova, A.; Ozerova, A.M.; Ozerov, I.V.; et al. Formation of γH2AX and pATM Foci in Human Mesenchymal Stem Cells Exposed to Low Dose-Rate Gamma-Radiation. Int. J. Mol. Sci. 2019, 20, 2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pustovalova, M.; Astrelina, T.A.; Grekhova, A.; Vorobyeva, N.; Tsvetkova, A.; Blokhina, T.; Nikitina, V.; Suchkova, Y.; Usupzhanova, D.; Brunchukov, V.; et al. Residual γH2AX foci induced by low dose X-ray radiation in bone marrow mesenchymal stem cells do not cause accelerated senescence in the progeny of irradiated cells. Aging 2017, 9, 2397–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, L.; Yu, N.; Furusawa, Y.; Okayasu, R.; Tong, J.; Cao, J.; Fan, S. Regulation of ATM in DNA double strand break repair accounts for the radiosensitivity in human cells exposed to high linear energy transfer ionizing radiation. Mutat. Res. Mol. Mech. Mutagen. 2009, 670, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; Jay-Gerin, J.-P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Kawai, H.; Sasatani, M.; Lizuka, D.; Masuda, Y.; Inaba, T.; Suzuki, K.; Ootsuyama, A. A novel ATM/TP53/p21-mediated checkpoint only activated by chronic γ-irradiation. PLoS ONE 2014, 9, e104279. [Google Scholar] [CrossRef] [Green Version]
- Cui, F.; Hou, J.; Huang, C.; Sun, X.; Zeng, Y.; Cheng, H.; Wang, H.; Li, C. C-Myc regulates radiation-induced G2/M cell cycle arrest and cell death in human cervical cancer cells. J. Obstet. Gynaecol. Res. 2017, 43, 729–735. [Google Scholar] [CrossRef]
- Foray, N.; Bourguignon, M.; Hamada, N. Individual response to ionizing radiation. Mutat. Res. Rev. Mutat. Res. 2016, 770, 369–386. [Google Scholar] [CrossRef]
- Human Radiosensitivity: Report of the Independent Advisory Group on Ionising Radiation; Health Protection Agency: London, UK, 2013.
- Tang, L.; Wei, F.; Wu, Y.; He, Y.; Shi, L.; Xiong, F.; Gong, Z.; Guo, C.; Li, X.; Deng, H.; et al. Role of metabolism in cancer cell radioresistance and radiosensitization methods. J. Exp. Clin. Cancer Res. 2018, 37, 87. [Google Scholar] [CrossRef]
- Scott, D.; Barber, J.B.; Levine, E.L.; Burrill, W.; Roberts, S.A. Radiation-induced micronucleus induction in lymphocytes identifies a high frequency of radiosensitive cases among breast cancer patients: A test for predisposition? Br. J. Cancer 1998, 77, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Palumbo, E.; Piotto, C.; Calura, E.; Fasanaro, E.; Groff, E.; Busato, F.; El Khouzai, B.; Rigo, M.; Baggio, L.; Romualdi, C.; et al. Individual Radiosensitivity in Oncological Patients: Linking Adverse Normal Tissue Reactions and Genetic Features. Front. Oncol. 2019, 9, 987. [Google Scholar] [CrossRef] [PubMed]
- Safwat, A.; Bentzen, S.M.; Turesson, I.; Hendry, J.H. Deterministic rather than stochastic factors explain most of the variation in the expression of skin telangiectasia after radiotherapy. Int. J. Radiat. Oncol. 2002, 52, 198–204. [Google Scholar] [CrossRef]
- Miyamoto, T.; Akutsu, S.N.; Tauchi, H.; Kudo, Y.; Tashiro, S.; Yamamoto, T.; Matsuura, S. Exploration of genetic basis underlying individual differences in radiosensitivity within human populations using genome editing technology. J. Radiat. Res. 2018, 59, ii75–ii82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subhashree, M.; Venkateswarlu, R.; Karthik, K.; Shangamithra, V.; Venkatachalam, P. DNA damage and the bystander response in tumor and normal cells exposed to X-rays. Mutat. Res. Toxicol. Environ. Mutagen. 2017, 821, 20–27. [Google Scholar] [CrossRef]
- Chang, H.H.Y.; Pannunzio, N.R.; Adachi, N.; Lieber, M.R. Non-homologous DNA end joining and alternative pathways to double-strand break repair. Nat. Rev. Mol. Cell Biol. 2017, 18, 495–506. [Google Scholar] [CrossRef]
- Xu, B.; Kim, S.-T.; Kastan, M.B. Involvement of Brca1 in S-Phase and G 2 -Phase Checkpoints after Ionizing Irradiation. Mol. Cell. Biol. 2001, 21, 3445–3450. [Google Scholar] [CrossRef] [Green Version]
- Sharan, S.K.; Morimatsu, M.; Albrecht, U.; Lim, D.-S.; Regel, E.; Dinh, C.; Sands, A.; Eichele, G.; Hasty, P.; Bradley, A. Embryonic lethality and radiation hypersensitivity mediated by Rad51 in mice lacking Brca2. Nature 1997, 386, 804–810. [Google Scholar] [CrossRef]
- Pearce, M.S.; A Salotti, J.; Little, M.; McHugh, K.; Lee, C.; Kim, K.P.; Howe, N.L.; Ronckers, C.M.; Rajaraman, P.; Craft, A.W.; et al. Radiation exposure from CT scans in childhood and subsequent risk of leukaemia and brain tumours: A retrospective cohort study. Lancet 2012, 380, 499–505. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.-F.; Lin, R.-X.; Huang, J.; Zhou, Z.; Yang, J.; Guo, G.-Z.; Wang, S.-Q. Identification of Differentially Expressed Genes Contributing to Radioresistance in Lung Cancer Cells using Microarray Analysis. Radiat. Res. 2005, 164, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Hung, J.-H.; Weng, Z. Analysis of Microarray and RNA-seq Expression Profiling Data. Cold Spring Harb. Protoc. 2016. [Google Scholar] [CrossRef]
- Lacombe, J.; Sima, C.; Amundson, S.A.; Zenhausern, F. Candidate gene biodosimetry markers of exposure to external ionizing radiation in human blood: A systematic review. PLoS ONE 2018, 13, e0198851. [Google Scholar] [CrossRef]
- Young, A.; Berry, R.; Holloway, A.F.; Blackburn, N.B.; Dickinson, J.L.; Skala, M.; Phillips, J.L.; Brettingham-Moore, K.H. RNA-seq profiling of a radiation resistant and radiation sensitive prostate cancer cell line highlights opposing regulation of DNA repair and targets for radiosensitization. BMC Cancer 2014, 14, 808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Rao, L.; Wang, H.L.; Mao, Z.W.; Lei, R.H.; Yang, Z.Y.; Qing, H.; Deng, Y.L. Transcriptome analysis of glioma cells for the dynamic response to γ-irradiation and dual regulation of apoptosis genes: A new insight into radiotherapy for glioblastomas. Cell Death Dis. 2013, 4, e895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashi-Elkeles, S.; Warnatz, H.-J.; Elkon, R.; Kupershtein, A.; Chobod, Y.; Paz, A.; Amstislavskiy, V.; Sultan, M.; Safer, H.; Nietfeld, W.; et al. Parallel Profiling of the Transcriptome, Cistrome, and Epigenome in the Cellular Response to Ionizing Radiation. Sci. Signal. 2014, 7, rs3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, M.S.; Van Vleet, T.R.; Ciurlionis, R.; Buck, W.R.; Mittelstadt, S.W.; Blomme, E.A.G.; Liguori, M.J. Comparison of RNA-Seq and Microarray Gene Expression Platforms for the Toxicogenomic Evaluation of Liver from Short-Term Rat Toxicity Studies. Front. Genet. 2019, 9, 636. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Yu, J.; Kong, D.; Xu, Y.; Zhang, Z.; Shui, J.; Li, Z.; Luo, H.; Wang, K. Population and single-cell transcriptome analyses reveal diverse transcriptional changes associated with radioresistance in esophageal squamous cell carcinoma. Int. J. Oncol. 2019, 55, 1237–1248. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, X.; Hou, Q.; Huang, M.; Zhang, H.; Jiang, Z.; Yue, J.; Wu, S. Single-cell RNA-seq of esophageal squamous cell carcinoma cell line with fractionated irradiation reveals radioresistant gene expression patterns. BMC Genom. 2019, 20, 611. [Google Scholar] [CrossRef]
- Gao, Y.; Duan, Q.; Wu, N.; Xu, B. A heterogeneous cellular response to ionizing radiation revealed by single cell transcriptome sequencing. Am. J. Cancer Res. 2021, 11, 513–529. [Google Scholar]
- Brown, A.A.; Viñuela, A.; Delaneau, O.; Spector, T.D.; Small, K.S.; Dermitzakis, E.T. Predicting causal variants affecting expression by using whole-genome sequencing and RNA-seq from multiple human tissues. Nat. Genet. 2017, 49, 1747–1751. [Google Scholar] [CrossRef] [Green Version]
- Cano-Gamez, E.; Trynka, G. From GWAS to Function: Using Functional Genomics to Identify the Mechanisms Underlying Complex Diseases. Front. Genet. 2020, 11, 424. [Google Scholar] [CrossRef]
- Vaisnav, M.; Xing, C.; Ku, H.-C.; Hwang, D.; Stojadinovic, S.; Pertsemlidis, A.; Abrams, J.M. Genome-Wide Association Analysis of Radiation Resistance in Drosophila melanogaster. PLoS ONE 2014, 9, e104858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehghan, A. Genome-Wide Association Studies; Springer: New York, NY, USA, 2018; pp. 37–49. [Google Scholar]
- Kerns, S.L.; Ostrer, H.; Stock, R.; Li, W.; Moore, J.; Pearlman, A.; Campbell, C.; Shao, Y.; Stone, N.; Kusnetz, L.; et al. Genome-Wide Association Study to Identify Single Nucleotide Polymorphisms (SNPs) Associated With the Development of Erectile Dysfunction in African-American Men After Radiotherapy for Prostate Cancer. Int. J. Radiat. Oncol. 2010, 78, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Finnon, P.; Robertson, N.; Dziwura, S.; Raffy, C.; Zhang, W.; Ainsbury, L.; Kaprio, J.; Badie, C.; Bouffler, S. Evidence for significant heritability of apoptotic and cell cycle responses to ionising radiation. Qual. Life Res. 2008, 123, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Curwen, G.B.; Cadwell, K.K.; Winther, J.F.; Tawn, E.J.; Rees, G.S.; Olsen, J.H.; Rechnitzer, C.; Schroeder, H.; Guldberg, P.; Cordell, H.J.; et al. The heritability of G2chromosomal radiosensitivity and its association with cancer in Danish cancer survivors and their offspring. Int. J. Radiat. Biol. 2010, 86, 986–995. [Google Scholar] [CrossRef] [Green Version]
- Zyla, J.; Kabacik, S.; O’Brien, G.; Wakil, S.; Al-Harbi, N.; Kaprio, J.; Badie, C.; Polanska, J.; Alsbeih, G. Combining CDKN1A gene expression and genome-wide SNPs in a twin cohort to gain insight into the heritability of individual radiosensitivity. Funct. Integr. Genom. 2019, 19, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-C.; Yue, X.; Zhang, R.-X.; Liu, T.-Y.; Pan, Z.-Z.; Yang, M.-J.; Lu, Z.-H.; Wang, Z.-Y.; Peng, J.-H.; Le, L.-Y.; et al. Genome-wide RNAi Screening Identifies RFC4 as a Factor That Mediates Radioresistance in Colorectal Cancer by Facilitating Nonhomologous End Joining Repair. Clin. Cancer Res. 2019, 25, 4567–4579. [Google Scholar] [CrossRef] [Green Version]
- Herr, P.; Lundin, C.; Evers, B.; Ebner, D.; Bauerschmidt, C.; Kingham, G.; Palmai-Pallag, T.; Mortusewicz, O.; Frings, O.; Sonnhammer, E.L.L.; et al. A genome-wide IR-induced RAD51 foci RNAi screen identifies CDC73 involved in chromatin remodeling for DNA repair. Cell Discov. 2015, 1, 15034. [Google Scholar] [CrossRef] [Green Version]
- van Haaften, G.; Vastenhouw, N.L.; Nollen, E.A.A.; Plasterk, R.H.A.; Tijsterman, M. Gene interactions in the DNA damage-response pathway identified by genome-wide RNA-interference analysis of synthetic lethality. Proc. Natl. Acad. Sci. USA 2004, 101, 12992–12996. [Google Scholar] [CrossRef] [Green Version]
- van Haaften, G.; Romeijn, R.; Pothof, J.; Koole, W.; Mullenders, L.H.F.; Pastink, A.; Plasterk, R.H.A. Identification of Conserved Pathways of DNA-Damage Response and Radiation Protection by Genome-Wide RNAi. Curr. Biol. 2006, 16, 1344–1350. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.-D.; Yu, J.; Sun, Z.-Y.; Chen, Y.; Zheng, H.-M.; Lin, M.-L.; Ou-Yang, S.; Liu, G.-L.; Zhang, J.-W.; Shao, F.-M. Genome-wide CRISPR/Cas9 screening identifies CARHSP1 responsible for radiation resistance in glioblastoma. Cell Death Dis. 2021, 12, 724. [Google Scholar] [CrossRef]
- Ziyan, Z.; Shen, M.; Jixi, L.; Kang, L.; Shi, S.; Chen, W.; Chen, S.; Yin, Y.; Qin, Y.; Kang, M. Genome-Scale CRISPR-Cas9 Knockout Screening in Nasopharyngeal Carcinoma for Radiosensitive and Radioresistant Genes. Int. J. Radiat. Oncol. 2021, 111, e389. [Google Scholar] [CrossRef]
- Hayman, T.J.; Baro, M.; MacNeil, T.; Phoomak, C.; Aung, T.N.; Cui, W.; Leach, K.; Iyer, R.; Challa, S.; Sandoval-Schaefer, T.; et al. STING enhances cell death through regulation of reactive oxygen species and DNA damage. Nat. Commun. 2021, 12, 2327. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Li, L.; Fan, K.; Li, Y.; Gao, Y. A Genome-Scale CRISPR Knock-Out Screen Identifies MicroRNA-5197-5p as a Promising Radiosensitive Biomarker in Colorectal Cancer. Front. Oncol. 2021, 11, 696713. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Pierce, S.E.; Li, A.; Spees, K.; Anderson, G.R.; Seoane, J.A.; Lo, Y.-H.; Dubreuil, M.; Olivas, M.; Kamber, R.A.; et al. CRISPR screens in cancer spheroids identify 3D growth-specific vulnerabilities. Nature 2020, 580, 136–141. [Google Scholar] [CrossRef]
- Oh, J.H.; Kerns, S.; Ostrer, H.; Powell, S.N.; Rosenstein, B.; Deasy, J. Computational methods using genome-wide association studies to predict radiotherapy complications and to identify correlative molecular processes. Sci. Rep. 2017, 7, 43381. [Google Scholar] [CrossRef]
- Ngo, V.; Davis, R.E.; Lamy, L.; Yu, X.; Zhao, H.; Lenz, G.; Lam, L.T.; Dave, S.; Yang, L.; Powell, J.; et al. A loss-of-function RNA interference screen for molecular targets in cancer. Nature 2006, 441, 106–110. [Google Scholar] [CrossRef]
- Peña-Rico, M.A.; Calvo-Vidal, M.N.; Villalonga-Planells, R.; Martínez-Soler, F.; Giménez-Bonafé, P.; Navarro-Sabaté, À.; Tortosa, A.; Bartrons, R.; Manzano, A. TP53 induced glycolysis and apoptosis regulator (TIGAR) knockdown results in radiosensitization of glioma cells. Radiother. Oncol. 2011, 101, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Shahar, O.D.; Ram, E.V.S.R.; Shimshoni, E.; Hareli, S.; Meshorer, E.; Goldberg, M. Live imaging of induced and controlled DNA double-strand break formation reveals extremely low repair by homologous recombination in human cells. Oncogene 2011, 31, 3495–3504. [Google Scholar] [CrossRef]
- Koike-Yusa, H.; Li, Y.; Tan, E.-P.; Velasco-Herrera, M.D.C.; Yusa, K. Genome-wide recessive genetic screening in mammalian cells with a lentiviral CRISPR-guide RNA library. Nat. Biotechnol. 2013, 32, 267–273. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and Applications of CRISPR-Cas9 for Genome Engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Vachey, G.; Déglon, N. CRISPR/Cas9-Mediated Genome Editing for Huntington’s Disease. In Huntington’s Disease; Precious, S.V., Rosser, A.E., Dunnett, S.B., Eds.; Springer: New York, NY, USA, 2018; pp. 463–481. [Google Scholar]
- Moreno Roig, E.; Groot, A.J.; Yaromina, A.; Hendricks, T.C.; Barbeau, L.M.O.; Giuranno, L.; Dams, G. HIF-1α and HIF-2α Differently Regulate the Radiation Sensitivity of NSCLC Cells. Cells 2019, 8, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behbahani, R.; Danyaei, A.; Teimoori, A.; Neisi, N.; Tahmasbi, M. Breast cancer radioresistance may be overcome by osteopontin gene knocking out with CRISPR/Cas9 technique. Cancer/Radiothérapie 2021, 25, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Murakami, N.; Kühnel, A.; Schmid, T.E.; Ilicic, K.; Stangl, S.; Braun, I.S.; Gehrmann, M.; Molls, M.; Itami, J.; Multhoff, G. Role of membrane Hsp70 in radiation sensitivity of tumor cells. Radiat. Oncol. 2015, 10, 149. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Li, C.; Chen, F.; Shen, L.; Li, Z.; Li, N. CRISPR/Cas9 genome-wide screening identifies LUC7L2 that promotes radioresistance via autophagy in nasopharyngeal carcinoma cells. Cell Death Discov. 2021, 7, 392. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Wang, F.; Zhang, W.; Wang, D.; Li, X.; Bartlam, M.; Yao, X.; Rao, Z. Structure-Functional Analyses of CRHSP-24 Plasticity and Dynamics in Oxidative Stress Response. J. Biol. Chem. 2011, 286, 9623–9635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeiffer, J.R.; McAvoy, B.L.; Fecteau, R.E.; Deleault, K.M.; Brooks, S.A. CARHSP1 Is Required for Effective Tumor Necrosis Factor Alpha mRNA Stabilization and Localizes to Processing Bodies and Exosomes. Mol. Cell. Biol. 2011, 31, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Pei, X.; Li, S.-W.; Yang, J.; Li, C.; Tang, J.; Hu, K.; Huang, G.; Min, W.-P.; Sang, Y. CRISPR/Cas9 library screening uncovered methylated PKP2 as a critical driver of lung cancer radioresistance by stabilizing β-catenin. Oncogene 2021, 40, 2842–2857. [Google Scholar] [CrossRef]
- Meyers, R.M.; Bryan, J.G.; McFarland, J.M.; Weir, B.A.; Sizemore, A.E.; Xu, H.; Dharia, N.V.; Montgomery, P.G.; Cowley, G.S.; Pantel, S.; et al. Computational correction of copy number effect improves specificity of CRISPR–Cas9 essentiality screens in cancer cells. Nat. Genet. 2017, 49, 1779–1784. [Google Scholar] [CrossRef] [Green Version]
- Lan, B.; Zeng, S.; Zhang, S.; Ren, X.; Xing, Y.; Kutschick, I.; Pfeffer, S.; Frey, B.; Britzen-Laurent, N.; Grützmann, R.; et al. CRISPR-Cas9 Screen Identifies DYRK1A as a Target for Radiotherapy Sensitization in Pancreatic Cancer. Cancers 2022, 14, 326. [Google Scholar] [CrossRef]
- Amundson, S.A.; Do, K.T.; Vinikoor, L.C.; Lee, R.A.; Koch-Paiz, C.A.; Ahn, J.; Reimers, M.; Chen, Y.; Scudiero, D.A.; Weinstein, J.N.; et al. Integrating Global Gene Expression and Radiation Survival Parameters across the 60 Cell Lines of the National Cancer Institute Anticancer Drug Screen. Cancer Res. 2008, 68, 415–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lü, X.; de la Peña, L.; Barker, C.; Camphausen, K.; Tofilon, P.J. Radiation-Induced Changes in Gene Expression Involve Recruitment of Existing Messenger RNAs to and away from Polysomes. Cancer Res. 2006, 66, 1052–1061. [Google Scholar] [CrossRef] [Green Version]
- Kumaraswamy, S.; Chinnaiyan, P.; Shankavaram, U.T.; Lü, X.; Camphausen, K.; Tofilon, P.J. Radiation-Induced Gene Translation Profiles Reveal Tumor Type and Cancer-Specific Components. Cancer Res. 2008, 68, 3819–3826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stackhouse, C.T.; Anderson, J.C.; Yue, Z.; Nguyen, T.; Eustace, N.J.; Langford, C.P.; Wang, J.; Iv, J.R.R.; Xing, C.; Mikhail, F.M.; et al. An in vivo model of glioblastoma radiation resistance identifies long non-coding RNAs and targetable kinases. JCI Insight 2022, e148717. [Google Scholar] [CrossRef] [PubMed]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef]
- Newby, G.A.; Yen, J.S.; Woodard, K.J.; Mayuranathan, T.; Lazzarotto, C.R.; Li, Y.; Sheppard-Tillman, H.; Porter, S.N.; Yao, Y.; Mayberry, K.; et al. Base editing of haematopoietic stem cells rescues sickle cell disease in mice. Nature 2021, 595, 295–302. [Google Scholar] [CrossRef]
- Nelson, J.W.; Randolph, P.B.; Shen, S.P.; Everette, K.A.; Chen, P.J.; Anzalone, A.V.; An, M.; Newby, G.A.; Chen, J.C.; Hsu, A.; et al. Engineered pegRNAs improve prime editing efficiency. Nat. Biotechnol. 2021, 40, 402–410. [Google Scholar] [CrossRef]
- Greve, B.; Bölling, T.; Amler, S.; Rössler, U.; Gomolka, M.; Mayer, C.; Popanda, O.; Dreffke, K.; Rickinger, A.; Fritz, E.; et al. Evaluation of Different Biomarkers to Predict Individual Radiosensitivity in an Inter-Laboratory Comparison–Lessons for Future Studies. PLoS ONE 2012, 7, e47185. [Google Scholar] [CrossRef]


{kind=link}
| Authors | Method/Dose Type | Model/Cell Type | Findings |
|---|---|---|---|
| Wang et al. [38] | Genome-wide RNAi screen/ Single dose 6 Gy, X-ray | Colorectal cancer cells exposed to X-rays both in vitro and in a mouse model | RFC4 protects colorectal cancer cells from radiation-induced DSBs and apoptosis both in vitro and in vivo; RFC4 enhances radioresistance. |
| Herr et al. [39] | Genome-wide RNAi screen/ Single dose 4 Gy (1.96 Gy min−1, Cs137) | Human bone osteosarcoma epithelial cells (U2OS line) | CDC73 is an important regulator of HRR-mediated DNA repair and genome stability. CDC73 enhances radioresistance. |
| van Haaften et al. [40] | Genome-wide RNAi screen/ Single dose 60 Gy, Gammacell 1000 (Cs-137) | C. elegans strains: wild-type Bristol N2, NL1832 (pk732), and TY1774 yIs2 [xol-1::lacZ rol-6 (pRF4)] IV. | Genes involved in the cellular response to DNA DSBs were identified. |
| van Haaften et al. [41] | Genome-wide RNAi screen/Single dose 140 Gy a Gammacell 1000 (Cs-137) | C. elegans strains were used: wild-type Bristol N2, atm-1 (gk186), lig-4 (ok716), and cku-80 (rb964) | A total of 45 C. elegans genes were identified that increased sensitivity to ionizing radiation in germ cells. |
| Kerns et al. [34] | GWAS/ 39 to 42 fractions of 1.8 Gy Xray | DNA isolated from lymphocytes | The location of SNP that is associated with erectile dysfunction as a side effect of RT was identified. These SNPs are specific for only patients with African ancestry. |
| Zyla et al. [37] | GWAS/ Single dose of 2 Gy of X-ray (0.5 Gy/min) | Blood T lymphocytes | SNPs influencing radiation sensitivity were identified. |
| Vaisnav et al. [32] | GWAS/Continuous exposure (4 h and 45 min) of gamma rays, 4.85 Gy/min, resulting in a total dose of 1382 Gy | Drosophila Genetic Reference Panel (DGRP) | Novel genes associated with variation in radiation resistance were identified. |
| Zhu et al. [42] | Whole CRISPR-Cas9 screen (positive screen) Treated with dose rate of 12, 15 Gy/min with X-ray Irradiator for three rounds | Glioblastoma cells | CARHSP1 enhances radioresistance in glioblastoma cancer cells. |
| Ziyan et al. [43] | Whole CRISPR-Cas9 screen (negative screen)/ Single dose 2 Gy | Nasopharyngeal carcinoma | Nine genes involved in the radiosensitivity or radioresistance of NPC cells were identified. |
| Hayman et al. [44] | Whole CRISPR-Cas9 screen (positive screen) | Neck squamous carcinoma cells (HNSCC) | Knockout of STING significantly increases radiation survival in both in vitro and in vivo models. |
| Yu et al. [45] | Whole CRISPR-Cas9 screen (negative screen) 6 & 12 single doses of X rays; dose rate: 5 Gy/min | Colorectal cancer cells | By inhibiting expression of cell cycle regulatory protein CDK6 and promoting cell cycle arrest in G1/S phase, microRNA-5197-5p (miR-5197) was reported as a radiosensitization factor. |
| Han et al. [46] | Whole CRISPR-Cas9 screen (positive screen) | Non-small-cell lung carcinoma cell lines | Key differences between 2D monolayer and 3D spheroid cancer models in CRISPR screen was demonstrated. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamaddondoust, R.N.; Wong, A.; Chandrashekhar, M.; Azzam, E.I.; Alain, T.; Wang, Y. Identification of Novel Regulators of Radiosensitivity Using High-Throughput Genetic Screening. Int. J. Mol. Sci. 2022, 23, 8774. https://doi.org/10.3390/ijms23158774
Tamaddondoust RN, Wong A, Chandrashekhar M, Azzam EI, Alain T, Wang Y. Identification of Novel Regulators of Radiosensitivity Using High-Throughput Genetic Screening. International Journal of Molecular Sciences. 2022; 23(15):8774. https://doi.org/10.3390/ijms23158774
Chicago/Turabian StyleTamaddondoust, Rosette N., Alicia Wong, Megha Chandrashekhar, Edouard I. Azzam, Tommy Alain, and Yi Wang. 2022. "Identification of Novel Regulators of Radiosensitivity Using High-Throughput Genetic Screening" International Journal of Molecular Sciences 23, no. 15: 8774. https://doi.org/10.3390/ijms23158774
APA StyleTamaddondoust, R. N., Wong, A., Chandrashekhar, M., Azzam, E. I., Alain, T., & Wang, Y. (2022). Identification of Novel Regulators of Radiosensitivity Using High-Throughput Genetic Screening. International Journal of Molecular Sciences, 23(15), 8774. https://doi.org/10.3390/ijms23158774