
Discovery of Epipodophyllotoxin-Derived B2 as Promising XooFtsZ Inhibitor for Controlling Bacterial Cell Division: Structure-Based Virtual Screening, Synthesis, and SAR Study

 , and
, and
Abstract
: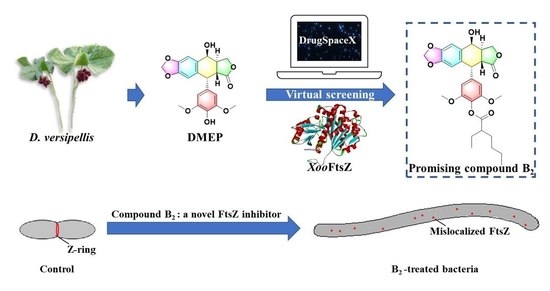
1. Introduction
2. Results and Discussion
2.1. Design and Synthesis of Target Compounds
2.2. The Anti-Xoo Bioactivity of Title Compounds
2.3. Investigation of Action Mechanism for Prepared Compound B2 Targeting XooFtsZ
2.4. Potential Mechanism of Action for 4′-Demethylepipodophyllotoxin (DMEP) Analogues
2.5. Outcome of SAR Study
2.6. In Vivo Trials against Rice Bacterial Leaf Blight Infected by Xoo
2.7. Assessment of Potential Risk of DMEP and Compound B2 through Phytotoxicity and Cytotoxicity Testing
3. Materials and Methods
3.1. Instruments and Chemicals
3.2. Experimental Section
3.3. The Strategy of Structure-Based Virtual Screening (SBVS)
3.4. Determination of the Binding Constant (KA) of Compounds-XooFtsZ Interaction
3.5. Morphological Studies Using Transmission Electron Microscopy (TEM)
3.6. Fourier Transform Infrared (FT-IR) Spectroscopy Analysis
3.7. Fluorescence Patterns for the Xoo Cells Triggered by Compounds
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DMEP | 4′-demethylepipodophyllotoxin |
| Xoo | Xanthomonas oryzae pv. Oryzae |
| FtsZ | filamentous temperature-sensitive protein Z |
| SBVS | Structure-based virtual screening |
| DMEOP | 4′-demethylpodophyllotoxin |
| D. versipellis | Dysosma versipellis |
References
- Li, P.; Hu, D.Y.; Xie, D.D.; Chen, J.X.; Jin, L.H.; Song, B.A. Design, synthesis, and evaluation of new sulfone derivatives containing a 1,3,4-oxadiazole moiety as active antibacterial agents. J. Agric. Food Chem. 2018, 66, 3093–3100. [Google Scholar] [CrossRef]
- Yuan, T.; Li, X.H.; Xiao, J.H.; Wang, S.P. Characterization of Xanthomonas oryzae-responsive cis-acting element in the promoter of rice race-specific susceptibility gene Xa13. Mol. Plant 2011, 4, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Stover, E.; Driggers, R.; Richardson, M.L.; Hall, D.G.; Lee, R.F. Incidence and severity of asiatic citrus canker on diverse citrus and citrus-related germplasm in a florida field planting. Hortscience 2014, 49, 4–9. [Google Scholar] [CrossRef]
- Borlaug, N.E. Feeding a world of 10 billion people: The miracle ahead. Biotechnol. Biotec. Eq. 1997, 11, 3–13. [Google Scholar] [CrossRef]
- Guo, S.X.; He, F.; Song, B.A.; Wu, J. Future direction of agrochemical development for plant disease in China. Food Energy Secur. 2021, 10, e293. [Google Scholar] [CrossRef]
- Sakschewski, B.; Bloh, W.V.; Huber, V.; Müller, C.; Bondeau, A. Feeding 10 billion people under climate change: How large is the production gap of current agricultural systems? Ecol. Model. 2014, 288, 103–111. [Google Scholar] [CrossRef]
- Campos, E.V.; Proença, P.L.; Oliveira, J.L.; Bakshi, M.; Abhilash, P.C.; Fraceto, L.F. Use of botanical insecticides for sustainable agriculture: Future perspectives. Ecol. Indic. 2019, 105, 483–495. [Google Scholar] [CrossRef]
- Donatelli, M.; Magarey, R.D.; Bregaglio, S.; Willocquet, L.; Whish, J.P.M.; Savary, S. Modelling the impacts of pests and diseases on agricultural systems. Agr. Syst. 2017, 155, 213–224. [Google Scholar] [CrossRef]
- Cheng, Y.L.; Zhang, Y.; Pei, R.J.; Xie, Y.F.; Yao, W.R.; Guo, Y.H.; Qian, H. Fast detection of bismerthiazol in cabbage based on fluorescence quenching of protein-capping gold nanoclusters. Anal. Sci. 2018, 34, 415–419. [Google Scholar] [CrossRef]
- Oguh, C.E.; Okpaka, C.O.; Ubani, C.S.; Okekeaji, U.; Joseph, P.S.; Amadi, E.U. Natural pesticides (biopesticides) and uses in pest management—A critical review. AJBGE 2019, 2, 1–18. Available online: https://sdiarticle4.com/prh/doc/Ms_AJBGE_53356.pdf (accessed on 1 October 2021).
- Wu, Y.X.; Ren, D.; Gao, C.; Li, J.Y.; Du, B.; Wang, Z.Y.; Qian, S. Recent advances for alkaloids as botanical pesticides for use in organic agriculture. Int. J. Pest Manage. 2021, 1–11. [Google Scholar] [CrossRef]
- Cantrell, C.L.; Dayan, F.E.; Duke, S.O. Natural products as sources for new pesticides. J. Nat. Prod. 2012, 75, 1231–1242. [Google Scholar] [CrossRef]
- Marrone, P.G. Pesticidal natural products–status and future potential. Pest Manag. Sci. 2019, 75, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.H.; Debbab, A.; Proksch, P. Fifty years of drug discovery from fungi. Fungal Divers. 2011, 50, 3–19. [Google Scholar] [CrossRef]
- Ravula, A.R.; Yenugu, S. Pyrethroid based pesticides–chemical and biological aspects. Crit. Rev. Toxicol. 2021, 51, 117–140. [Google Scholar] [CrossRef]
- Yu, X.; Che, Z.P.; Xu, H. Recent advances in the chemistry and biology of podophyllotoxins. Chem. Eur. J. 2017, 23, 4467–4526. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.K.; Guan, J.; Xiao, Z.; Mark Cosentino, L.; Lee, K.H. Anti-AIDS agents. Part 61: Anti-HIV activity of new podophyllotoxin derivatives. Bioorg. Med. Chem. 2004, 12, 4267–4273. [Google Scholar] [CrossRef]
- Zhang, Q.Y.; Zhao, W.; Tang, Y.J. Discover the leading compound of 4β-S-(5-fluorobenzoxazole)-4-deoxy-4′-demethylepipodophyllotoxin with millimolar-potency toxicity by modifying the molecule structure of podophyllotoxin. Eur. J. Med. Chem. 2018, 158, 951–964. [Google Scholar] [CrossRef]
- Zhao, W.; Cong, Y.; Li, H.M.; Li, S.Y.; Shen, Y.M.; Qi, Q.S.; Zhang, Y.M.; Li, Y.Z.; Tang, Y.J. Challenges and potential for improving the druggability of podophyllotoxin-derived drugs in cancer chemotherapy. Nat. Prod. Rep. 2021, 38, 470–488. [Google Scholar] [CrossRef]
- Zhi, X.Y.; Yang, C.; Zhang, R.; Hu, Y.; Ke, Y.Z.; Xu, H. Natural products-based insecticidal agents 13. Semisynthesis and insecticidal activity of novel phenazine derivatives of 4β-acyloxypodophyllotoxin modified in the E-ring against Mythimna separata Walker in vivo. Ind. Crop. Prod. 2013, 42, 520–526. [Google Scholar] [CrossRef]
- Zhi, X.Y.; Yu, X.; Yang, C.; Ding, G.D.; Chen, H.; Xu, H. Synthesis of 4β-acyloxypodophyllotoxin analogs modified in the C and E rings as insecticidal agents against Mythimna separata Walker. Bioorg. Med. Chem. Lett. 2014, 24, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.L.; Xu, M.; Li, S.C.; He, J.N.; Xu, H. Synthesis of some ester derivatives of 4′-demethoxyepipodophyllotoxin/2′-chloro-4′-demethoxyepipodophyllotoxin as insecticidal agents against oriental armyworm, Mythimna separata Walker. Bioorg. Med. Chem. Lett. 2017, 27, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ye, H.J.; Gao, X.H.; Feng, Y.M.; Shao, W.B.; Qi, P.Y.; Wu, Z.B.; Liu, L.W.; Wang, P.Y.; Yang, S. The discovery of natural 4′-demethylepipodophyllotoxin from renewable dysosma versipellis species as a novel bacterial cell division inhibitor for controlling intractable diseases in rice. Ind. Crop. Prod. 2021, 174, 114182. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, H.Y.; Shen, C.; Hu, X.P.; Gao, J.B.; Li, D.; Cao, D.S.; Hou, T.J. Combined strategies in structure-based virtual screening. Phys. Chem. Chem. Phys. 2020, 22, 3149–3159. [Google Scholar] [CrossRef]
- Yang, T.B.; Li, Z.J.; Chen, Y.J.; Feng, D.; Wang, G.C.; Fu, Z.Y.; Ding, X.Y.; Tan, X.Q.; Zhao, J.H.; Luo, X.M.; et al. DrugSpaceX: A large screenable and synthetically tractable database extending drug space. Nucleic Acids Res. 2021, 49, D1170–D1178. [Google Scholar] [CrossRef]
- Meng, X.Y.; Zhang, H.X.; Mezei, M.; Cui, M. Molecular Docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef]
- Novikov, F.N.; Chilov, G.G. Molecular docking: Theoretical background, practical applications and perspectives. Mendeleev Commun. 2009, 19, 237–242. [Google Scholar] [CrossRef]
- Ferreira, L.G.; dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular Docking and Structure-Based Drug Design Strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef]
- Anil Kumar, K.; Kumar Singh, S.; Siva Kumar, B.; Doble, M. Synthesis, anti-fungal activity evaluation and QSAR studies on podophyllotoxin derivatives. Cent. Eur. J. Chem. 2007, 5, 880–897. [Google Scholar] [CrossRef]
- Xiao, L.; Zhao, W.; Li, H.M.; Wan, D.J.; Li, D.S.; Chen, T.; Tang, Y.J. Design and synthesis of the novel DNA topoisomerase II inhibitors: Esterification and amination substituted 4′-demethylepipodophyllotoxin derivates exhibiting anti-tumor activity by activating ATM/ATR signaling pathways. Eur. J. Med. Chem. 2014, 80, 267–277. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Zhang, J.Q.; Hui, L.; Chen, S.W.; Tian, X. First synthesis and biological evaluation of novel spin-labeled derivatives of deoxypodophyllotoxin. Eur. J. Med. Chem. 2010, 45, 1673–1677. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.A.; Corral, J.M.M.D.; Gordaliza, M.; Go´mez-Zurita, M.A.; Puente, M.L.D.L.; Betancur-Galvis, L.A.; Sierra, J.; Feliciano, A.S. Antiviral Synthesis, cytotoxicity and antiviral activity of podophyllotoxin analogues modified in the E-ring. Eur. J. Med. Chem. 2003, 38, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.E.; Long, X.X.; Ni, W.Z.; Ye, Z.Q.; He, Z.L.; Stoffella, P.J.; Calvert, D.V. Assessing copper thresholds for phytotoxicity and potential dietary toxicity in selected vegetable crops. J. Environ. Sci. Health B 2002, 37, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.M.L. Tetrazolium (MTT) Assay for Cellular Viability and Activity. Methods Mol. Biol. 1998, 79, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T.J. Rapid colorimetic assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. Available online: https://www.nature.com/articles/srep42717 (accessed on 1 October 2021). [CrossRef]
- Zhou, X.; Feng, Y.M.; Qi, P.Y.; Shao, W.B.; Wu, Z.B.; Liu, L.W.; Wang, Y.; Ma, H.D.; Wang, P.Y.; Li, Z.; et al. Synthesis and Docking Study of N-(Cinnamoyl)-N′-(substituted) acryloyl Hydrazide Derivatives Containing Pyridinium Moieties as a Novel Class of Filamentous Temperature-Sensitive Protein Z Inhibitors against the Intractable Xanthomonas oryzae pv. oryzae Infections in Rice. J. Agric. Food Chem. 2020, 68, 8132–8142. [Google Scholar] [CrossRef]
- Wang, Z.C.; Li, X.Y.; Wang, W.L.; Zhang, W.Y.; Yu, L.; Hu, D.Y.; Song, B.A. Interaction research on the antiviral molecule Dufulin targeting on southern rice black streaked dwarf virus P9-1 nonstructural protein. Viruses 2015, 7, 1454–1473. [Google Scholar] [CrossRef]
- Bi, S.Y.; Ding, L.; Tian, Y.; Song, D.Q.; Zhou, X.; Liu, X.; Zhang, H.Q. Investigation of the interaction between flavonoids and human serum albumin. J. Mol. Struct. 2004, 703, 37–45. [Google Scholar] [CrossRef]
- Zhou, X.; Ye, Y.Q.; Liu, S.S.; Shao, W.B.; Liu, L.W.; Yang, S.; Wu, Z.B. Design, synthesis and anti-TMV activity of novel α-aminophosphonate derivatives containing a chalcone moiety that induce resistance against plant disease and target the TMV coat protein. Pestic. Biochem. Phys. 2021, 172, 104749. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, X.; Wu, H.G.; Yu, Z.Z.; Li, H.; Yang, S. Nanospheric heterogeneous acid-enabled direct upgrading of biomass feedstocks to novel benzimidazoles with potent antibacterial activities. Ind. Crop. Prod. 2020, 150, 112406. [Google Scholar] [CrossRef]
- Liu, C.; Cheng, F.F.; Yang, X.Q. Inactivation of soybean trypsin inhibitor by epigallocatechin gallate: Stopped-flow/fluorescence, thermodynamics, and docking studies. J. Agric. Food Chem. 2017, 65, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Pu, P.; Zheng, X.; Jiao, L.N.; Chen, L.; Yang, H.; Zhang, Y.H.; Liang, G.Z. Six flavonoids inhibit the antigenicity of β-lactoglobulin by noncovalent interactions: A spectroscopic and molecular docking study. Food Chem. 2021, 339, 128106. [Google Scholar] [CrossRef]
- Beuria, T.K.; Santra, M.K.; Panda, D. Sanguinarine blocks cytokinesis in bacteria by inhibiting FtsZ assembly and bundling. Biochemistry 2005, 44, 16584–16593. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Bhattacharya, A.; Rai, A.; Dhaked, H.P.S.; Awasthi, D.; Ojima, I.; Panda, D. SB-RA-2001 inhibits bacterial proliferation by targeting FtsZ assembly. Biochemistry 2014, 53, 2979–2992. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.Y.; Xiang, M.; Luo, M.; Liu, H.W.; Zhou, X.; Wu, Z.B.; Liu, L.W.; Li, Z.; Yang, S. Novel piperazine-tailored ursolic acid hybrids as significant antibacterial agents targeting phytopathogens Xanthomonas oryzae pv. oryzae and X. axonopodis pv. citri probably directed by activation of apoptosis. Pest Manag. Sci. 2020, 76, 2746–2754. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.L.; Huang, X.; Liu, L.W.; Wang, P.Y.; Long, Q.S.; Tao, Q.Q.; Li, Z.; Yang, S. Identification of racemic and chiral carbazole derivatives containing an isopropanolamine linker as prospective surrogates against plant pathogenic bacteria: In vitro and in vivo assays and quantitative proteomics. J. Agric. Food Chem. 2019, 67, 7512–7525. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.W.; Ji, Q.T.; Ren, G.G.; Wang, F.; Su, F.; Wang, P.Y.; Zhou, X.; Wu, Z.B.; Li, Z.; Yang, S. Antibacterial functions and proposed modes of action of novel 1,2,3,4-tetrahydro-β-carboline derivatives that possess an attractive 1,3-diaminopropan-2-ol pattern against rice bacterial blight, kiwifruit bacterial canker, and citrus bacterial canker. J. Agric. Food Chem. 2020, 68, 12558–12568. [Google Scholar] [CrossRef]
- Tao, Q.Q.; Liu, L.W.; Wang, P.Y.; Long, Q.S.; Zhao, Y.L.; Jin, L.H.; Xu, W.M.; Chen, Y.; Li, Z.; Yang, S. Synthesis and in vitro and in vivo biological activity evaluation and quantitative proteome profiling of oxadiazoles bearing flexible heterocyclic patterns. J. Agric. Food Chem. 2019, 67, 7626–7639. [Google Scholar] [CrossRef]
- Zeng, D.; Wang, M.W.; Xiang, M.; Liu, L.W.; Wang, P.Y.; Li, Z.; Yang, S. Design, synthesis, and antimicrobial behavior of novel oxadiazoles containing various N-containing heterocyclic pendants. Pest Manag. Sci. 2020, 76, 2681–2692. [Google Scholar] [CrossRef]
- Xiang, M.; Song, Y.L.; Ji, J.; Zhou, X.; Liu, L.W.; Wang, P.Y.; Wu, Z.B.; Li, Z.; Yang, S. Synthesis of novel 18β-glycyrrhetinic piperazine amides displaying significant in vitro and in vivo antibacterial activities against intractable plant bacterial diseases. Pest Manag. Sci. 2020, 76, 2959–2971. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.Y.; Yuan, W.C.; Li, Y.; Huang, X.H.; Guo, Q.; Tang, Z.W.; Fang, Z.Y.; Lin, H.; Wong, W.L.; Wong, K.Y.; et al. Antibacterial activity of indolyl-quinolinium derivatives and study their mode of action. Bioorg. Med. Chem. 2019, 27, 1274–1282. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | Inhibition (%) | EC50 (mg L−1) | MIC (mg L−1) | ||||
|---|---|---|---|---|---|---|---|
| Entry | R1 | R2 | R3 | 100 (mg L−1) | 20 (mg L−1) | ||
| A1 |  |  |  | 18.3 ± 2.15 | 10.6 ± 0.90 | >200 | >400 |
| B1 | 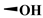 |  |  | 17.4 ± 6.00 | 5.33 ± 1.13 | >200 | >400 |
| B2 |  |  |  | 44.2 ± 3.20 | 7.30 ± 2.20 | 153 ± 9.20 | 400 |
| B3 |  |  |  | 10.2 ± 1.20 | 4.33 ± 0.80 | >200 | >400 |
| DMEP |  | H |  | 81.3 ± 10.2 | 26.2 ± 4.10 | 39.7 ± 0. 18 | 50 |
| BT | 92.7 ± 1.70 | 30.0 ± 2.20 | 36.3 ± 2.50 | 50 | |||
| Compounds | Inhibition Rate (%) | IC50 (μM) | |
|---|---|---|---|
| 200 μM | 100 μM | ||
| B2 | 54.8 ± 5.20 | 48.6 ± 4.10 | 159.4 ± 16.7 |
| Berberine hydrochloride | 48.7 ± 4.10 | 34.1 ± 2.60 | 225.0 ± 18.5 |
| Compounds | Stern–Volmer Quenching Constants | Binding Parameters | ||||
|---|---|---|---|---|---|---|
| Ksv (M−1) | Kq (M−1 S−1) | R | KA (M−1) | n | R2 | |
| B2 | 3.845 × 103 | 3.845 × 1011 | 0.96 | 103.22 | 0.92 | 0.95 |
| DMEP | 7.593 × 103 | 7.593 × 1011 | 0.98 | 103.48 | 0.92 | 0.98 |
| Berberine | 1.789 × 104 | 1.789 × 1012 | 0.91 | 103.44 | 0.81 | 0.98 |
 | Inhibition (%) | EC50 (mg L−1) | MIC (mg L−1) | ||||
|---|---|---|---|---|---|---|---|
| Entry | R1 | R2 | R3 | 100 (mg L−1) | 20 (mg L−1) | ||
| Podophyllotoxin |  | CH3 |  | - | - | >200 | >400 |
| Picropodophyllotoxin |  | CH3 |  | - | - | >200 | >400 |
| DMEOP |  | H |  | - | - | >200 | >400 |
| Deoxypodophyllotoxin | H | CH3 |  | 10.3 ± 3.50 | 12.2 ± 4.80 | >200 | >400 |
| Teniposide | 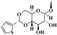 | H |  | 20.3 ± 9.21 | - | >200 | >400 |
| Etoposide |  | H |  | 18.8 ± 9.60 | - | >200 | >400 |
| DMEP |  | H |  | 81.3 ± 10.2 | 26.2 ± 4.10 | 39.7 ± 0. 18 | 50 |
| BT | 92.7 ± 1.70 | 30.0 ± 2.20 | 36.3 ± 2.50 | 50 | |||
| Compounds | Stern–Volmer Quenching Constants | Binding Parameters | ||||
|---|---|---|---|---|---|---|
| Ksv (M−1) | Kq (M−1 S−1) | R | KA (M−1) | n | R2 | |
| Podophyllotoxin | 2.203 × 103 | 2.203 × 1011 | 0.94 | 101.34 | 0.54 | 0.94 |
| Picropodophyllotoxin | 3.603 × 103 | 3.603 × 1011 | 0.95 | 101.81 | 0.63 | 0.96 |
| DMEOP | 1.757 × 103 | 1.757 × 1011 | 0.98 | 102.26 | 0.79 | 0.96 |
| Deoxypodophyllotoxin | 2.276 × 103 | 2.276 × 1011 | 0.98 | 101.04 | 0.48 | 0.97 |
| Teniposide | 2.920 × 103 | 2.920 × 1011 | 0.98 | 101.73 | 0.61 | 0.97 |
| Etoposide | 2.396 × 103 | 2.396 × 1011 | 0.98 | 102.50 | 0.82 | 0.97 |
| DMEP | 7.593 × 103 | 7.593 × 1011 | 0.98 | 103.48 | 0.92 | 0.98 |
| Berberine | 1.789 × 104 | 1.789 × 1012 | 0.91 | 103.44 | 0.81 | 0.98 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.-L.; Liu, S.-S.; Yang, J.; Xie, J.; Zhou, X.; Wu, Z.-B.; Liu, L.-W.; Wang, P.-Y.; Yang, S. Discovery of Epipodophyllotoxin-Derived B2 as Promising XooFtsZ Inhibitor for Controlling Bacterial Cell Division: Structure-Based Virtual Screening, Synthesis, and SAR Study. Int. J. Mol. Sci. 2022, 23, 9119. https://doi.org/10.3390/ijms23169119
Song Y-L, Liu S-S, Yang J, Xie J, Zhou X, Wu Z-B, Liu L-W, Wang P-Y, Yang S. Discovery of Epipodophyllotoxin-Derived B2 as Promising XooFtsZ Inhibitor for Controlling Bacterial Cell Division: Structure-Based Virtual Screening, Synthesis, and SAR Study. International Journal of Molecular Sciences. 2022; 23(16):9119. https://doi.org/10.3390/ijms23169119
Chicago/Turabian StyleSong, Ying-Lian, Shuai-Shuai Liu, Jie Yang, Jiao Xie, Xiang Zhou, Zhi-Bing Wu, Li-Wei Liu, Pei-Yi Wang, and Song Yang. 2022. "Discovery of Epipodophyllotoxin-Derived B2 as Promising XooFtsZ Inhibitor for Controlling Bacterial Cell Division: Structure-Based Virtual Screening, Synthesis, and SAR Study" International Journal of Molecular Sciences 23, no. 16: 9119. https://doi.org/10.3390/ijms23169119
APA StyleSong, Y.-L., Liu, S.-S., Yang, J., Xie, J., Zhou, X., Wu, Z.-B., Liu, L.-W., Wang, P.-Y., & Yang, S. (2022). Discovery of Epipodophyllotoxin-Derived B2 as Promising XooFtsZ Inhibitor for Controlling Bacterial Cell Division: Structure-Based Virtual Screening, Synthesis, and SAR Study. International Journal of Molecular Sciences, 23(16), 9119. https://doi.org/10.3390/ijms23169119