
Molecular Events behind the Selectivity and Inactivation Properties of Model NaK-Derived Ion Channels


 , ,
, , 
Abstract
:1. Introduction
2. Results
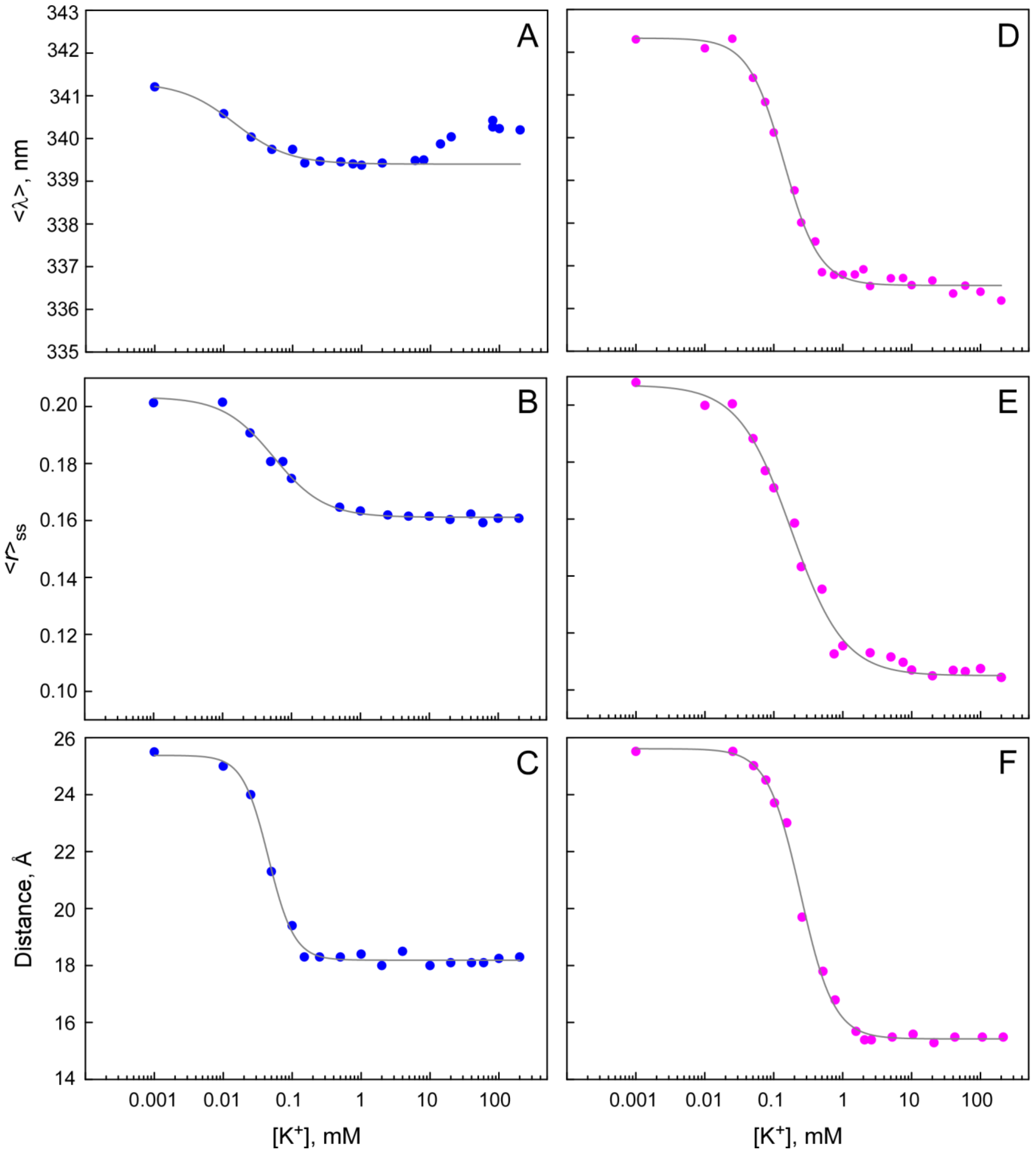
2.1. Functional Behavior of Y55W Mutants of Purified NaK and NaK2K Channels Reconstituted into Asolectin Giant Liposomes
2.2. Fluorescent Properties of Y55W Mutants of NaK and NaK2K Channels in DDM Micelles
2.3. Cation Binding to the Y55W Mutants of NaK and NaK2K Channels
3. Discussion
4. Materials and Methods
4.1. Molecular Biology, Protein Expression, and Purification
| NaK | MAKDKEFQVL10FVLTILTLIS20GTIFYSTVEG30LRPIDALWFS40VVTLTTVGDG50 |
| NaK2K | MAKDKEFQVL10FVLTILTLIS20GTIFYSTVEG30LRPIDALWFS40VVTLTTVGYG50 |
| NaK | NFSPQTDFGK60IFTILYIFIG70IGLVFGFIHK80LAVNVQLPSI90LSNLVPRGSR100 |
| NaK2K | DFSPQTDFGK60IFTILYIFIG70IGLVFGFIHK80LAVNVQLPSI90LSNLVPRGSR100 |
| NaK | SHHHHHH107 |
| NaK2K | SHHHHHH107 |
4.2. Ion Channel Reconstitution into Liposomes
4.3. Ion Channel Functional Measurements
4.4. Steady-State Fluorescence Spectroscopy
4.5. Fluorescence Monitoring of Cation Binding
4.6. Time-Resolved Fluorescence Spectroscopy
4.7. Determination of the Rotational Correlation Time of the Channel-DDM Complexes
4.8. Analysis of the Time-Resolved Anisotropy Decays to Determine the W55-W55 Intersubunit Distances
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doyle, D.A.; Morais, C.J.; Pfuetzner, R.A.; Kuo, A.; Gulbis, J.M.; Cohen, S.L.; Chait, B.T.; MacKinnon, R. The Structure of the Potassium Channel: Molecular Basis of K+ Conduction and Selectivity. Science 1998, 280, 69–77. [Google Scholar] [CrossRef]
- Uysal, S.; Vasquez, V.; Tereshko, V.; Esaki, K.; Fellouse, F.A.; Sidhu, S.S.; Koide, S.; Perozo, E.; Kossiakoff, A. Crystal Structure of Full-Length KcsA in Its Closed Conformation. Proc. Natl. Acad. Sci. USA 2009, 106, 16644–16649. [Google Scholar] [CrossRef]
- Köpfer, D.A.; Song, C.; Gruene, T.; Sheldrick, G.M.; Zachariae, U.; De Groot, B.L. Ion Permeation in K+ Channels Occurs by Direct Coulomb Knock-On. Science 2014, 346, 352–355. [Google Scholar] [CrossRef]
- Baukrowitz, T.; Yellen, G. Modulation of K+current by Frequency and External [K+]: A Tale of Two Inactivation Mechanisms. Neuron 1995, 15, 951–960. [Google Scholar] [CrossRef]
- Xu, Y.; Bhate, M.P.; McDermott, A.E. Transmembrane Allosteric Energetics Characterization for Strong Coupling between Proton and Potassium Ion Binding in the KcsA Channel. Proc. Natl. Acad. Sci. USA 2017, 114, 8788–8793. [Google Scholar] [CrossRef]
- Zhou, Y.; Morais-Cabral, J.H.; Kaufman, A.; Mackinnon, R. Chemistry of Ion Coordination and Hydration Revealed by a K+ Channel-Fab Complex at 2.0 Å Resolution. Nature 2001, 414, 43–48. [Google Scholar] [CrossRef]
- Lockless, S.W.; Zhou, M.; MacKinnon, R. Structural and Thermodynamic Properties of Selective Ion Binding in a K+ Channel. PLoS Biol. 2007, 5, 1079–1088. [Google Scholar] [CrossRef]
- Gao, L.; Mi, X.; Paajanen, V.; Wang, K.; Fan, Z. Activation-Coupled Inactivation in the Bacterial Potassium Channel KcsA. Proc. Natl. Acad. Sci. USA 2005, 102, 17630–17635. [Google Scholar]
- López-Barneo, J.; Hoshi, T.; Heinemann, S.H.; Aldrich, R.W. Effects of External Cations and Mutations in the Pore Region on C-Type Inactivation of Shaker Potassium Channels. Recept. Channels 1993, 1, 61–71. [Google Scholar]
- Rasmusson, R.L.; Morales, M.J.; Wang, S.; Liu, S.; Campbell, D.L.; Brahmajothi, M.V.; Strauss, H.C. Inactivation of Voltage-Gated Cardiac K+ Channels. Circ. Res. 1998, 82, 739–750. [Google Scholar] [CrossRef]
- Santos, J.S.; Syeda, R.; Montal, M. Stabilization of the Conductive Conformation of a Voltage-Gated K+ (Kv) Channel: The Lid Mechanism. J. Biol. Chem. 2013, 288, 16619–16628. [Google Scholar] [CrossRef]
- Swenson, R.P.; Armstrong, C.M. K+channels Close More Slowly in the Presence of External K+and Rb+. Nature 1981, 291, 427–429. [Google Scholar] [CrossRef]
- Derebe, M.G.; Sauer, D.B.; Zeng, W.; Alam, A.; Shia, N.; Jiang, Y. Tuning the Ion Selectivity of Tetrameric Cation Channels by Changing the Number of Ion Binding Sites. Proc. Natl. Acad. Sci. USA 2011, 108, 598–602. [Google Scholar] [CrossRef]
- Shi, N.; Ye, S.; Alam, A.; Chen, L.; Jiang, Y. Atomic Structure of a Na+- and K+-Conducting Channel. Nature 2006, 440, 570–574. [Google Scholar] [CrossRef]
- Alam, A.; Jiang, Y. High-Resolution Structure of the Open NaK Channel. Nat. Struct. Mol. Biol. 2009, 16, 30–34. [Google Scholar] [CrossRef]
- Amer Alam, Y.J. Structural Analysis of Ion Selectivity in the NaK Channel. Nat. Struct. Mol. Biol. 2009, 16, 35–41. [Google Scholar]
- Lewis, A.; Kurauskas, V.; Tonelli, M.; Henzler-Wildman, K. Ion-Dependent Structure, Dynamics, and Allosteric Coupling in a Non-Selective Cation Channel. Nat. Commun. 2021, 12, 6225. [Google Scholar] [CrossRef]
- Roy, R.N.; Hendriks, K.; Kopec, W.; Abdolvand, S.; Weiss, K.L.; De Groot, B.L.; Lange, A.; Sun, H.; Coates, L. Structural Plasticity of the Selectivity Filter in a Nonselective Ion Channel. IUCrJ 2021, 8, 421–430. [Google Scholar] [CrossRef]
- Shi, C.; He, Y.; Hendriks, K.; De Groot, B.L.; Cai, X.; Tian, C.; Lange, A.; Sun, H. A Single NaK Channel Conformation Is Not Enough for Non-Selective Ion Conduction. Nat. Commun. 2018, 9, 717. [Google Scholar] [CrossRef]
- Liu, S.; Gonen, T. MicroED Structure of the NaK Ion Channel Reveals a Na+ Partition Process into the Selectivity Filter. Commun. Biol. 2018, 1, 38. [Google Scholar] [CrossRef]
- LeMasurier, M.; Heginbotham, L.; Miller, C. KcsA: It’s a Potassium Channel. J. Gen. Physiol. 2001, 118, 303–314. [Google Scholar]
- Nimigean, C.M.; Miller, C. Na+ Block and Permeation in a K+ Channel of Known Structure. J. Gen. Physiol. 2002, 120, 323. [Google Scholar] [CrossRef]
- Montoya, E.; Lourdes Renart, M.; Marcela Giudici, A.; Poveda, J.A.; Fernández, A.M.; Morales, A.; González-Ros, J.M. Differential Binding of Monovalent Cations to KcsA: Deciphering the Mechanisms of Potassium Channel Selectivity. Biochim. Biophys. Acta Biomembr. 2017, 1859, 779–788. [Google Scholar] [CrossRef]
- Chakrapani, S.; Cordero-Morales, J.F.; Perozo, E. A Quantitative Description of KcsA Gating I: Macroscopic Currents. J. Gen. Physiol. 2007, 130, 465–478. [Google Scholar]
- Renart, M.L.; Giudici, A.M.; Poveda, J.A.; Fedorov, A.; Berberan-Santos, M.N.; Prieto, M.; Díaz-García, C.; González-Ros, J.M.; Coutinho, A. Conformational Plasticity in the KcsA Potassium Channel Pore Helix Revealed by Homo-FRET Studies. Sci. Rep. 2019, 9, 6215–6228. [Google Scholar] [CrossRef]
- Giudici, A.; Díaz-García, C.; Renart, M.; Coutinho, A.; Prieto, M.; González-Ros, J.; Poveda, J. Tetraoctylammonium, a Long Chain Quaternary Ammonium Blocker, Promotes a Noncollapsed, Resting-Like Inactivated State in KcsA. Int. J. Mol. Sci. 2021, 22, 490. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy Principles of Fluorescence Spectroscopy; Springer: New York, NY, USA, 2006; ISBN 978-0-387-31278-1/978-0-387-46312-4. [Google Scholar]
- Stryer, L.; Haugland, R.P. Energy Transfer: A Spectroscopic Ruler. Proc. Natl. Acad. Sci. USA 1967, 58, 719–726. [Google Scholar] [CrossRef]
- Ghosh, S.; Saha, S.; Goswami, D.; Bilgrami, S.; Mayor, S. Dynamic Imaging of Homo-FRET in Live Cells by Fluorescence Anisotropy Microscopy. Methods Enzymol. 2012, 505, 291–327. [Google Scholar] [CrossRef]
- Chan, F.; Kaminski, C.; Schierle, G.K. Homo-FRET Fluorescence Anisotropy Imaging as a Tool to Study Molecular Self-Assembly in Live Cells. Chemphyschem 2011, 12, 500–509. [Google Scholar] [CrossRef]
- Su, Z.; Brown, E.C.; Wang, W.; MacKinnon, R. Novel Cell-Free High-Throughput Screening Method for Pharmacological Tools Targeting K+ Channels. Proc. Natl. Acad. Sci. USA 2016, 113, 5748–5753. [Google Scholar] [CrossRef]
- Sauer, D.B.; Zeng, W.; Canty, J.; Lam, Y.; Jiang, Y. Sodium and Potassium Competition in Potassium-Selective and Non-Selective Channels. Nat. Commun. 2013, 4, 2721. [Google Scholar] [CrossRef]
- Sauer, D.B.; Zenga, W.; Raghunathan, S.; Jiang, Y. Protein Interactions Central to Stabilizing the K+ Channel Selectivity Filter in a Four-Sited Configuration for Selective K+ Permeation. Proc. Natl. Acad. Sci. USA 2011, 108, 16634–16639. [Google Scholar] [CrossRef]
- Langan, P.S.; Vandavasi, V.G.; Kopec, W.; Sullivan, B.; Afonne, P.V.; Weiss, K.L.; De Groot, B.L.; Coates, L. The Structure of a Potassium-Selective Ion Channel Reveals a Hydrophobic Gate Regulating Ion Permeation. IUCrJ 2020, 7, 835–843. [Google Scholar] [CrossRef]
- Molina, M.L.; Barrera, F.N.; Fernandez, A.M.; Poveda, J.A.; Renart, M.L.; Encinar, J.A.; Riquelme, G.; Gonzalez-Ros, J.M. Clustering and Coupled Gating Modulate the Activity in KcsA, a Potassium Channel Model. J. Biol. Chem. 2006, 281, 18837–18848. [Google Scholar]
- Shen, R.; Guo, W.; Zhong, W. Hydration Valve Controlled Non-Selective Conduction of Na(+) and K(+) in the NaK Channel. Biochim. Biophys. Acta 2010, 1798, 1474–1479. [Google Scholar] [CrossRef]
- Díaz-García, C.; Renart, M.L.; Poveda, J.A.; Giudici, A.M.; González-Ros, J.M.; Prieto, M.; Coutinho, A. Probing the Structural Dynamics of the Activation Gate of KcsA Using Homo-FRET Measurements. Int. J. Mol. Sci. 2021, 22, 11954. [Google Scholar] [CrossRef]
- Hendriks, K.; Öster, C.; Shi, C.; Sun, H.; Lange, A. Sodium Ions Do Not Stabilize the Selectivity Filter of a Potassium Channel. J. Mol. Biol. 2021, 433, 167091. [Google Scholar] [CrossRef]
- Renart, M.L.; Barrera, F.N.; Molina, M.L.; Encinar, J.A.; Poveda, J.A.; Fernández, A.M.; Gómez, J.; González-Ros, J.M. Effects of Conducting and Blocking Ions on the Structure and Stability of the Potassium Channel KcsA. J. Biol. Chem. 2006, 281, 29905–29915. [Google Scholar] [CrossRef]
- Valeur, B.; Weber, G. Resolution of the Fluorescence Excitation Spectrum of Indole into the 1La and 1Lb Excitation Bands. Photochem. Photobiol. 1977, 25, 441–444. [Google Scholar] [CrossRef]
- Zhou, M.; Morais-Cabral, J.H.; Mann, S.; MacKinnon, R. Potassium Channel Receptor Site for the Inactivation Gate and Quaternary Amine Inhibitors. Nature 2001, 411, 657–661. [Google Scholar] [CrossRef]
- Morais-Cabral, J.H.; Zhou, Y.; MacKinnon, R. Energetic Optimization of Ion Conduction Rate by the K+ Selectivity Filter. Nature 2001, 414, 37–40. [Google Scholar] [CrossRef]
- Triano, I.; Barrera, F.N.; Renart, M.L.; Molina, M.L.; Fernandez-Ballester, G.; Poveda, J.A.; Fernandez, A.M.; Encinar, J.A.; Ferrer-Montiel, A.V.; Otzen, D.; et al. Occupancy of Nonannular Lipid Binding Sites on KcsA Greatly Increases the Stability of the Tetrameric Protein. Biochemistry 2010, 49, 5397–5404. [Google Scholar] [CrossRef]
- Sehgal, P.; Otzen, D.E. Thermodynamics of Unfolding of an Integral Membrane Protein in Mixed Micelles. Protein Sci. 2006, 15, 890–899. [Google Scholar]
- Renart, M.L.; Triano, I.; Poveda, J.A.; Encinar, J.A.; Fernández, A.M.; Ferrer-Montiel, A.V.; Gómez, J.; González Ros, J.M. Ion Binding to KcsA: Implications in Ion Selectivity and Channel Gating. Biochemistry 2010, 49, 9480–9487. [Google Scholar] [CrossRef]
- Krishnan, M.N.; Bingham, J.P.; Siew, H.L.; Trombley, P.; Moczydlowski, E. Functional Role and Affinity of Inorganic Cations in Stabilizing the Tetrameric Structure of the KcsA K+ Channel. J. Gen. Physiol. 2005, 126, 271–283. [Google Scholar] [CrossRef]
- Krishnan, M.N.; Trombley, P.; Moczydlowski, E.G. Thermal Stability of the K+ Channel Tetramer: Cation Interactions and the Conserved Threonine Residue at the Innermost Site (S4) of the KcsA Selectivity Filter. Biochemistry 2008, 47, 5354–5367. [Google Scholar] [CrossRef]
- Giudici, A.M.; Renart, M.L.; Díaz-García, C.; Morales, A.; Poveda, J.A.; González-Ros, J.M. Accessibility of Cations to the Selectivity Filter of KcsA in the Inactivated State: An Equilibrium Binding Study. Int. J. Mol. Sci. 2019, 20, 689. [Google Scholar] [CrossRef]
- Callis, P.R. 1La and 1Lb Transitions of Tryptophan: Applications of Theory and Experimental Observations to Fluorescence of Proteins. Methods Enzymol. 1997, 278, 113–150. [Google Scholar] [CrossRef]
- Napolitano, L.M.R.; Bisha, I.; De March, M.; Marchesi, A.; Arcangeletti, M.; Demitri, N.; Mazzolini, M.; Rodriguez, A.; Magistrato, A.; Onesti, S.; et al. A Structural, Functional, and Computational Analysis Suggests Pore Flexibility as the Base for the Poor Selectivity of CNG Channels. Proc. Natl. Acad. Sci. USA 2015, 112, E3619–E3628. [Google Scholar] [CrossRef]
- Kopec, W.; Köpfer, D.A.; Vickery, O.N.; Bondarenko, A.S.; Jansen, T.L.C.; de Groot, B.L.; Zachariae, U. Direct Knock-on of Desolvated Ions Governs Strict Ion Selectivity in K+ Channels. Nat. Chem. 2018, 10, 813–820. [Google Scholar] [CrossRef]
- Wang, S.; Lee, S.J.; Maksaev, G.; Fang, X.; Zuo, C.; Nichols, C.G. Potassium Channel Selectivity Filter Dynamics Revealed by Single-Molecule FRET. Nat. Chem. Biol. 2019, 15, 377–383. [Google Scholar] [CrossRef]
- Thompson, A.N.; Kim, I.; Panosian, T.D.; Iverson, T.M.; Allen, T.W.; Nimigean, C.M. Mechanism of Potassium-Channel Selectivity Revealed by Na(+) and Li(+) Binding Sites within the KcsA Pore. Nat. Struct. Mol. Biol. 2009, 16, 1317–1324. [Google Scholar] [CrossRef]
- Ulmschneider, M.B.; Bagnéris, C.; McCusker, E.C.; DeCaen, P.G.; Delling, M.; Clapham, D.E.; Ulmschneider, J.P.; Wallace, B.A. Molecular Dynamics of Ion Transport through the Open Conformation of a Bacterial Voltage-Gated Sodium Channel. Proc. Natl. Acad. Sci. USA 2013, 110, 6364–6369. [Google Scholar] [CrossRef]
- Naylor, C.E.; Bagnéris, C.; DeCaen, P.G.; Sula, A.; Scaglione, A.; Clapham, D.E.; Wallace, B. Molecular Basis of Ion Permeability in a Voltage-Gated Sodium Channel. EMBO J. 2016, 35, 820–830. [Google Scholar] [CrossRef]
- Renart, M.L.; Montoya, E.; Giudici, A.M.; Poveda, J.A.; Fernández, A.M.; Morales, A.; González-Ros, J.M. Selective Exclusion and Selective Binding Both Contribute to Ion Selectivity in KcsA, a Model Potassium Channel. J. Biol. Chem. 2017, 292, 15552–15560. [Google Scholar] [CrossRef]
- Cheng, W.W.L.; McCoy, J.G.; Thompson, A.N.; Nichols, C.G.; Nimigean, C.M. Mechanism for Selectivity-Inactivation Coupling in KcsA Potassium Channels. Proc. Natl. Acad. Sci. USA 2011, 108, 5272–5277. [Google Scholar] [CrossRef]
- Cuello, L.G.; Jogini, V.; Cortes, D.M.; Perozo, E. Structural Mechanism of C-Type Inactivation in K(+) Channels. Nature 2010, 466, 203–208. [Google Scholar]
- Devaraneni, P.K.; Komarov, A.G.; Costantino, C.A.; Devereaux, J.J.; Matulef, K.; Valiyaveetil, F.I. Semisynthetic K+ Channels Show That the Constricted Conformation of the Selectivity Filter Is Not the C-Type Inactivated State. Proc. Natl. Acad. Sci. USA 2013, 110, 15698–15703. [Google Scholar] [CrossRef]
- Liu, S.; Lockless, S.W. Equilibrium Selectivity Alone Does Not Create K+ -Selective Ion Conduction in K+ Channels. Nat. Commun. 2013, 4, 2746–2753. [Google Scholar] [CrossRef]
- Hoshi, T.; Zagotta, W.N.; Aldrich, R.W. Biophysical and Molecular Mechanisms of Shaker Potassium Channel Inactivation. Science 1990, 250, 533–538. [Google Scholar] [CrossRef]
- Fedida, D.; Maruoka, N.D.; Lin, S. Modulation of Slow Inactivation in Human Cardiac Kv1.5 Channels by Extra- and Intracellular Permeant Cations. J. Physiol. 1999, 515 Pt 2, 315–329. [Google Scholar] [CrossRef]
- Rasmusson, R.L.; Morales, M.J.; Castellino, R.C.; Zhang, Y.; Campbell, D.L.; Strauss, H.C. C-Type Inactivation Controls Recovery in a Fast Inactivating Cardiac K+ Channel (Kv1.4) Expressed in Xenopus Oocytes. J. Physiol. 1995, 489 Pt 3, 709–721. [Google Scholar] [CrossRef]
- Boiteux, C.; Posson, D.J.; Allen, T.W.; Nimigean, C.M. Selectivity Filter Ion Binding Affinity Determines Inactivation in a Potassium Channel. Proc. Natl. Acad. Sci. USA 2020, 117, 29968–29978. [Google Scholar] [CrossRef]
- Cordero-Morales, J.F.; Cuello, L.G.; Zhao, Y.; Jogini, V.; Cortes, D.M.; Roux, B.; Perozo, E. Molecular Determinants of Gating at the Potassium-Channel Selectivity Filter. Nat. Struct. Mol. Biol. 2006, 13, 311–318. [Google Scholar]
- Molina, M.L.; Encinar, J.A.; Barrera, F.N.; Fernández-Ballester, G.; Riquelme, G.; González-Ros, J.M. Influence of C-Terminal Protein Domains and Protein-Lipid Interactions on Tetramerization and Stability of the Potassium Channel KcsA. Biochemistry 2004, 43, 14924–14931. [Google Scholar] [CrossRef]
- Pace, C.N.; Scholtz, J.M. Measuring the Conformational Stability of a Protein. In Protein Structure: A Practical Approach; Oxford University Press: Oxford, UK, 1997; pp. 297–321. [Google Scholar]
- Poveda, J.A.; Giudici, A.M.; Renart, M.L.; Millet, O.; Morales, A.; González-Ros, J.M.; Oakes, V.; Furini, S.; Domene, C. Modulation of the Potassium Channel KcsA by Anionic Phospholipids: Role of Arginines at the Non-Annular Lipid Binding Sites. Biochim. Biophys. Acta Biomembr. 2019, 1861, 183029–183043. [Google Scholar] [CrossRef]
- Hille, B. Ion Channels of Excitable Membranes, 3rd ed.; University Press: Oxford, UK, 2001. [Google Scholar]
- Berberan-Santos, M.N.; Prieto, M.J.E. Energy Transfer in Spherical Geometry. Application to Micelles. J. Chem. Soc. Faraday Trans. 2 Mol. Chem. Phys. 1987, 83, 1391–1410. [Google Scholar] [CrossRef]
- Stryer, L. Fluorescence Energy Transfer as a Spectroscopic Ruler. Annu. Rev. Biochem. 1978, 47, 819–846. [Google Scholar] [CrossRef]
- Poveda, J.A.; Prieto, M.; Encinar, J.A.; González-Ros, J.M.; Mateo, C.R. Intrinsic Tyrosine Fluorescence as a Tool to Study the Interaction of the Shaker B “Ball” Peptide with Anionic Membranes. Biochemistry 2003, 42, 7124–7132. [Google Scholar]
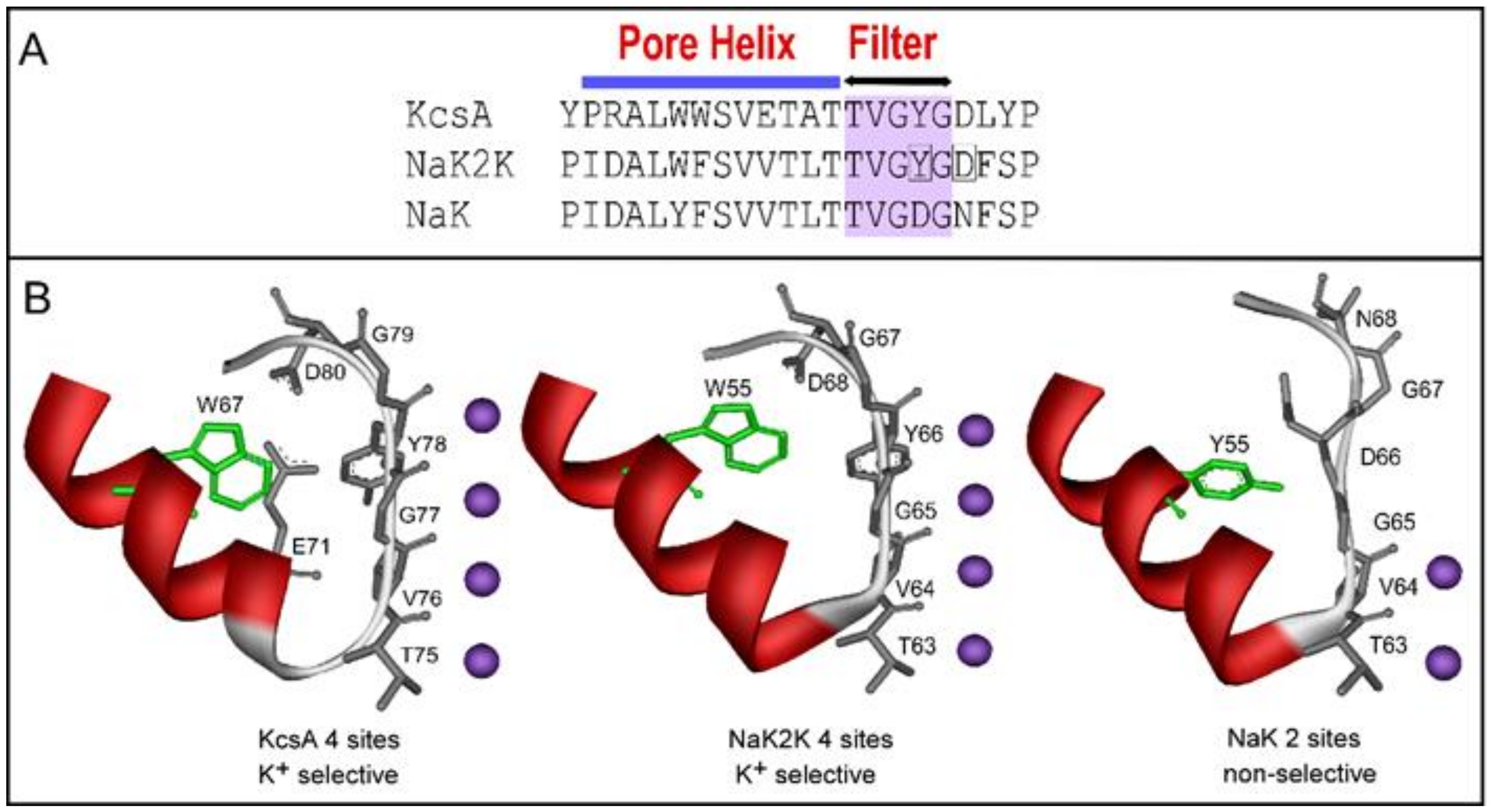

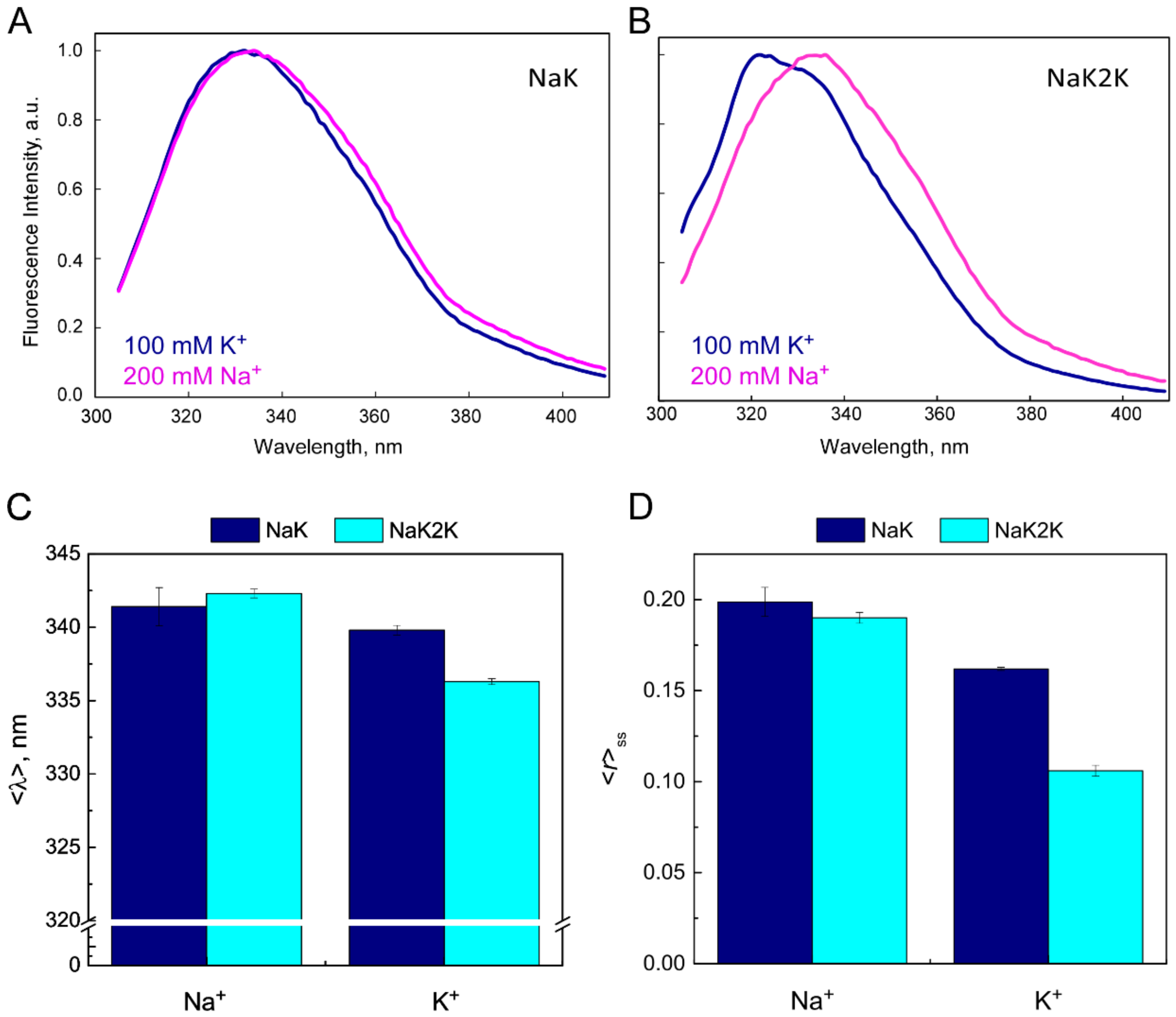

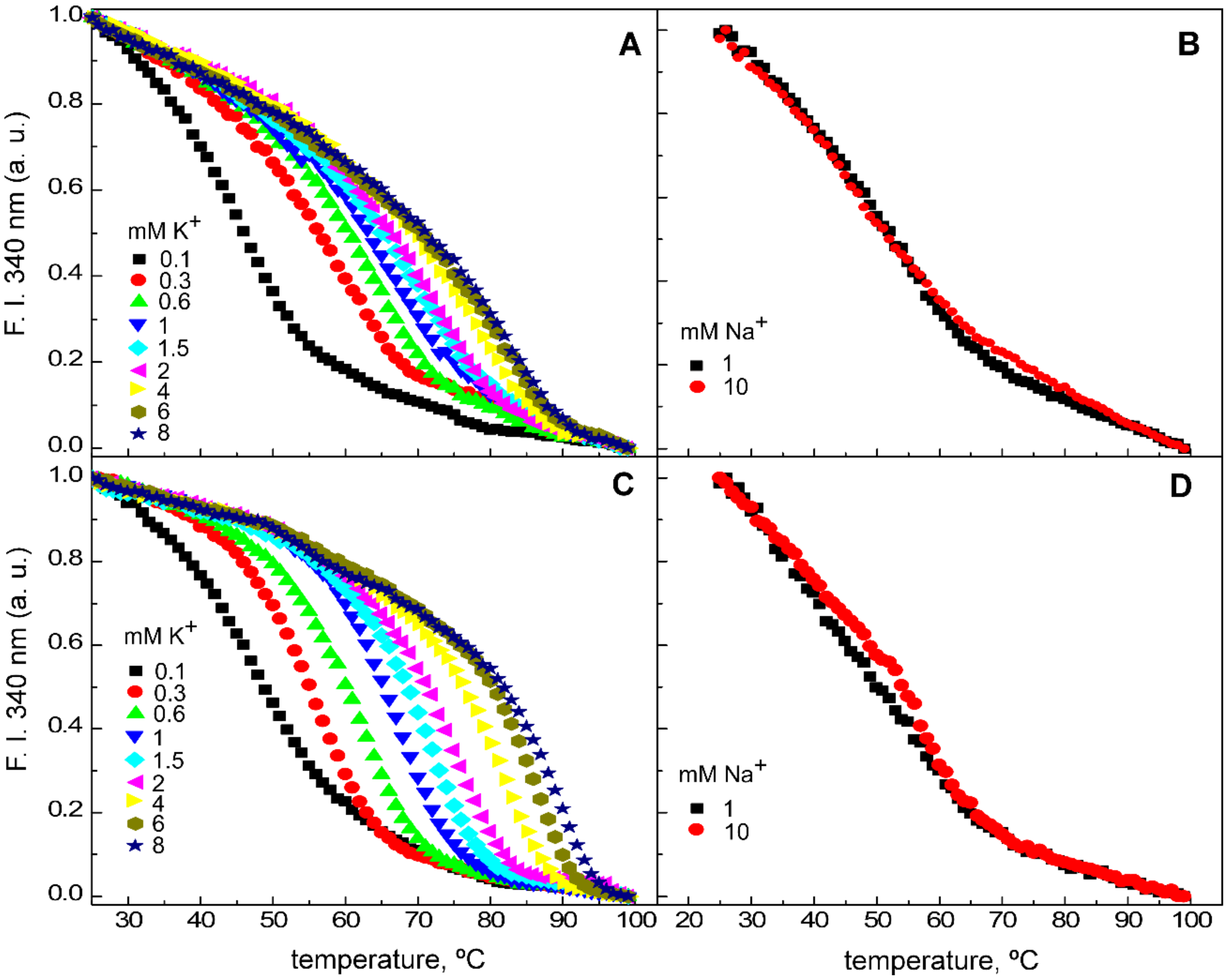
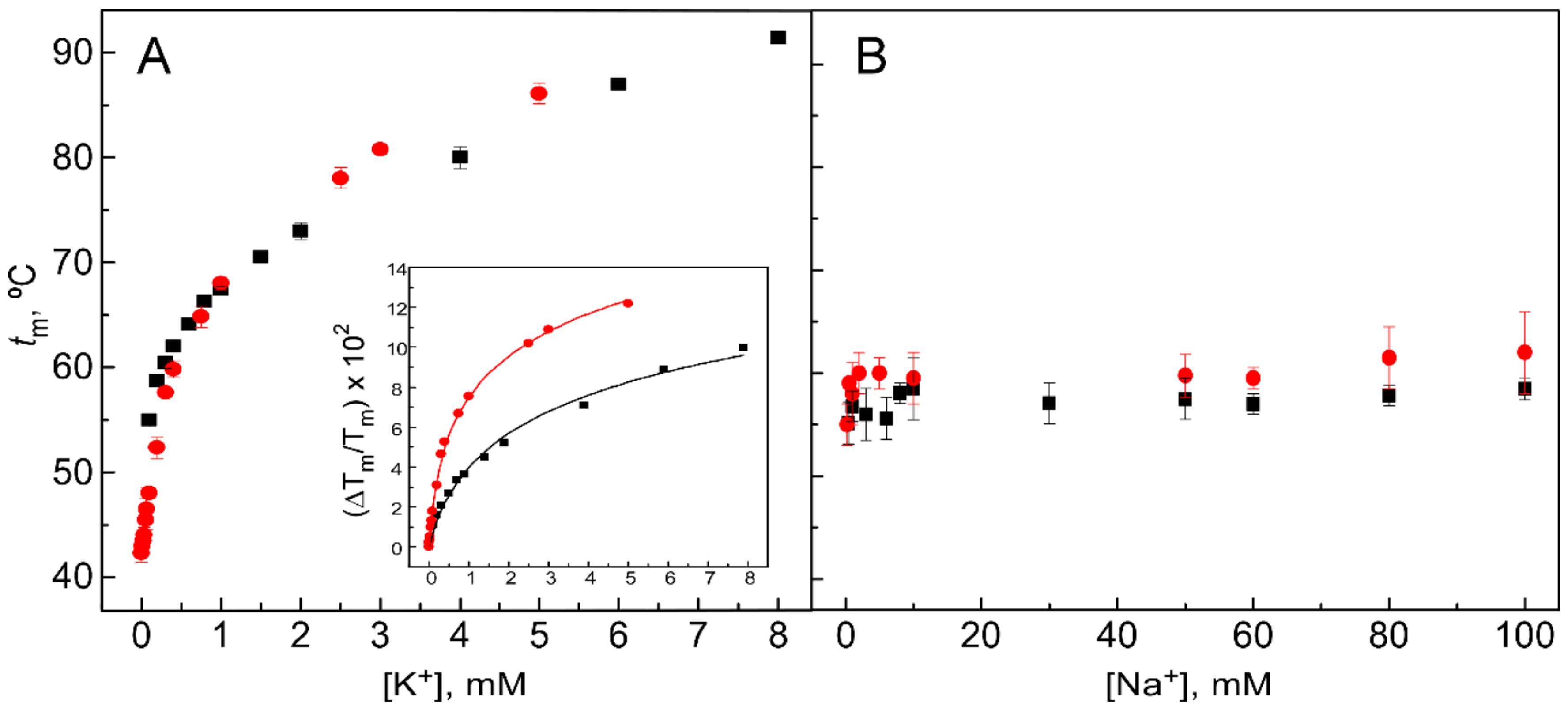

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Channel | K+ | Na+ | ||||
|---|---|---|---|---|---|---|
| R0 * (Å) | Homo-FRET Distance ** (Å) | X-ray Distance *** (Å) | R0 * (Å) | Homo-FRET Distance ** (Å) | X-ray Distance *** (Å) | |
| NaK | 13.0 | 18.3 ± 0.2 | 19.4 (PDB: 3E8H) | 12.9 | 25.8 ± 0.8 | 19.1 (PDB: 2AHY) |
| NaK2K | 13.7 | 15.5 ± 0.1 | 16.9 (PDB: 3T4Z) | 12.2 | 25 ± 1 | n.a.**** |
| KcsA | 12.0 | 15.3 ± 0.1 | 15.2 (PDB: 3PJS) | 12.0 | 18.2 ± 0.1 | 18.1 (PDB: 2ITC) |
| Observable Parameter | NaK KD (mM) | NaK2K KD (mM) |
|---|---|---|
| tm | 0.5 (0.3–0.7) | 0.10 a (0.05–0.2) |
| <λ> | 0.05 (0.01–0.09) | 0.12 (0.07–0.17) |
| <r> | 0.033 (0.014–0.052) | 0.17 a (0.13–0.21) |
| R | 0.044 (0.030–0.057) | 0.23 a (0.21–0.24) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giudici, A.M.; Renart, M.L.; Coutinho, A.; Morales, A.; González-Ros, J.M.; Poveda, J.A. Molecular Events behind the Selectivity and Inactivation Properties of Model NaK-Derived Ion Channels. Int. J. Mol. Sci. 2022, 23, 9246. https://doi.org/10.3390/ijms23169246
Giudici AM, Renart ML, Coutinho A, Morales A, González-Ros JM, Poveda JA. Molecular Events behind the Selectivity and Inactivation Properties of Model NaK-Derived Ion Channels. International Journal of Molecular Sciences. 2022; 23(16):9246. https://doi.org/10.3390/ijms23169246
Chicago/Turabian StyleGiudici, Ana Marcela, María Lourdes Renart, Ana Coutinho, Andrés Morales, José Manuel González-Ros, and José Antonio Poveda. 2022. "Molecular Events behind the Selectivity and Inactivation Properties of Model NaK-Derived Ion Channels" International Journal of Molecular Sciences 23, no. 16: 9246. https://doi.org/10.3390/ijms23169246
APA StyleGiudici, A. M., Renart, M. L., Coutinho, A., Morales, A., González-Ros, J. M., & Poveda, J. A. (2022). Molecular Events behind the Selectivity and Inactivation Properties of Model NaK-Derived Ion Channels. International Journal of Molecular Sciences, 23(16), 9246. https://doi.org/10.3390/ijms23169246